
Genetic Analysis of Methane Emission Traits in Holstein Dairy Cattle
 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics and Animal Care
2.2. Data Collection
2.2.1. Ontario Dairy Research Centre
2.2.2. Dairy Research and Technology Center
2.3. Variation in Methane Testing
2.4. Data Set and Methane Traits
2.5. Variance Components
2.6. Rank Correlations and Accuracy of Estimated Breeding Values
3. Results and Discussion
3.1. Descriptive Statistics
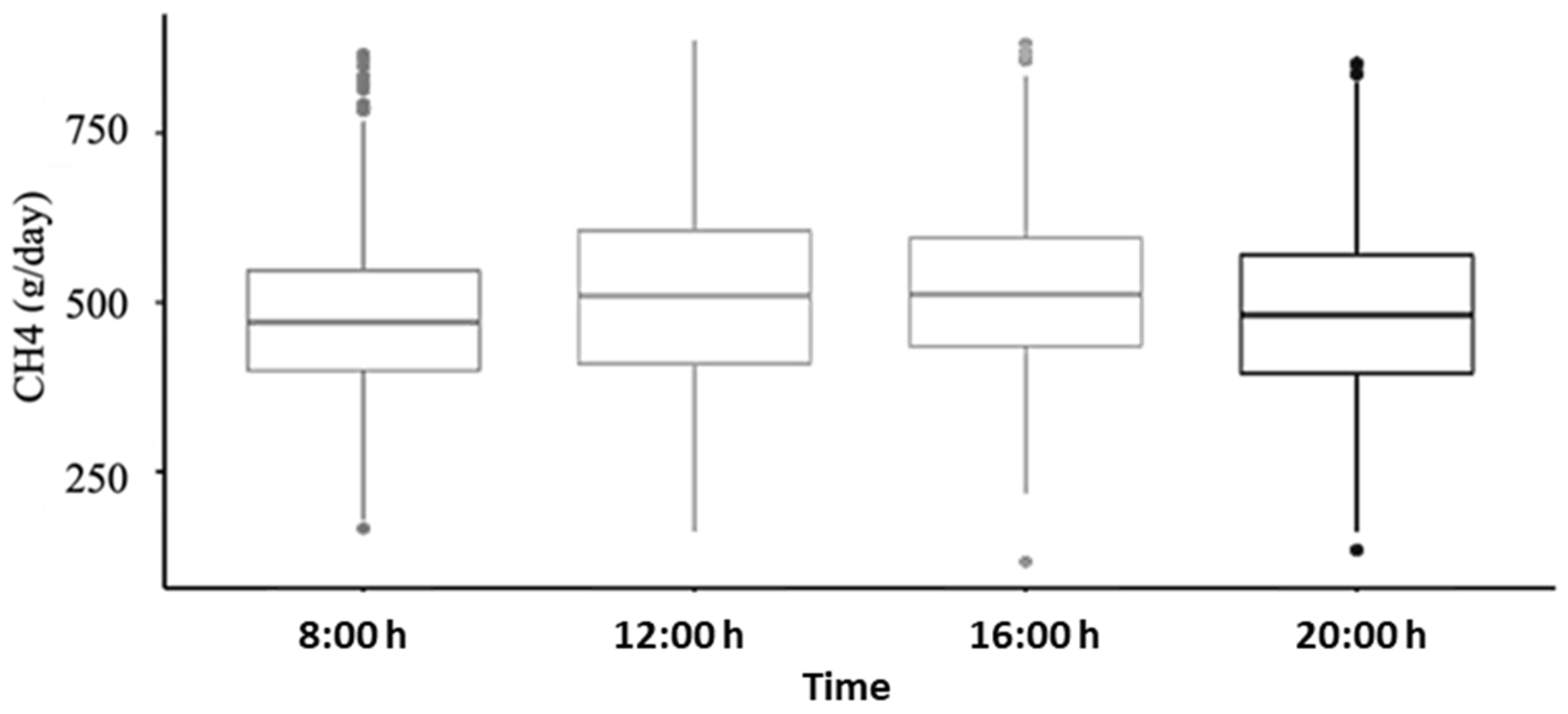
3.2. Variation over Time
3.3. Variance Component Estimates
3.4. Correlations
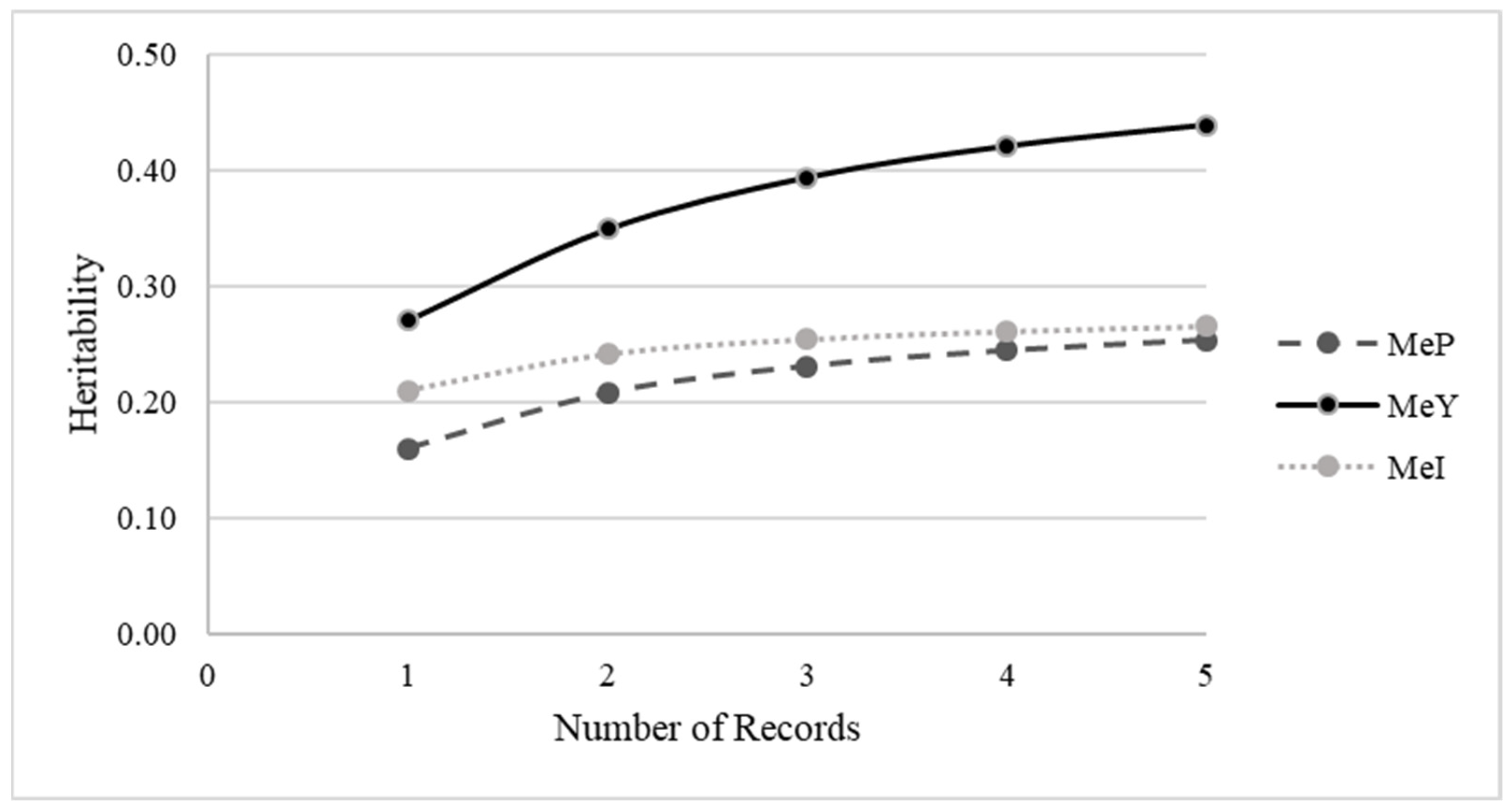
3.5. Accuracy of EBV and EBV Rank Correlations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berndt, A.; Boland, T.M.; Deighton, M.H.; Gere, J.I.; Grainger, C.; Hegarty, R.S.; Iwaasa, A.D.; Koolaard, G.C.; Lassey, K.R.; Luo, D.; et al. Guidelines for Use of Sulphur Hexafluoride (SF 6) Tracer Technique to Measure Enteric Methane Emissions from Ruminants; Ministry of Primary Industries: Wellington, New Zealand, 2014; pp. 1–66.
- Hammond, K.J.; Crompton, L.A.; Bannink, A.; Dijkstra, J.; Yáñez-Ruiz, D.R.; O’Kiely, P.; Kebreab, E.; Eugène, M.A.; Yu, Z.; Shingfield, K.J.; et al. Review of current in vivo measurement techniques for quantifying enteric methane emission from ruminants. Anim. Feed. Sci. Technol. 2016, 219, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Huhtanen, P.; Ramin, M.; Hristov, A.N. Enteric methane emission can be reliably measured by the GreenFeed monitoring unit. Livest. Sci. 2019, 222, 31–40. [Google Scholar] [CrossRef]
- González-Recio, O.; López-Paredes, J.; Ouatahar, L.; Charfeddine, N.; Ugarte, E.; Alenda, R.; Jiménez-Montero, J.A. Mitigation of greenhouse gases in dairy cattle via genetic selection: 2. Incorporating methane emissions into the breeding goal. J. Dairy Sci. 2020, 103, 7210–7221. [Google Scholar] [CrossRef]
- López-Paredes, J.; Goiri, I.; Atxaerandio, R.; García-Rodríguez, A.; Ugarte, E.; Jiménez-Montero, J.A.; Alenda, R.; González-Recio, O. Mitigation of greenhouse gases in dairy cattle via genetic selection: 1. Genetic parameters of direct methane using noninvasive methods and proxies of methane. J. Dairy Sci. 2020, 103, 7199–7209. [Google Scholar] [CrossRef] [PubMed]
- ICAR Section 20—Recording Dairy Cattle Methane Emission for Genetic Evaluation. Available online: https://www.icar.org/Guidelines/20-Recording-Dairy-Cattle-Methane-Emission-for-Genetic-Evaluation.pdf (accessed on 20 July 2020).
- National Inventory Report 2019 National Inventory Report 1990–2017: Greenhouse Gas Sources and Sinks in Canada. Available online: https://publications.gc.ca/collections/collection_2019/eccc/En81-4-2017-1-eng.pdf (accessed on 20 July 2020).
- VandeHaar, M.J.; Armentano, L.E.; Weigel, K.; Spurlock, D.M.; Tempelman, R.J.; Veerkamp, R. Harnessing the genetics of the modern dairy cow to continue improvements in feed efficiency. J. Dairy Sci. 2016, 99, 4941–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, D.; Bannink, A.; Hatew, B.; van Laar, H.; Dijkstra, J. Effects of grass silage quality and level of feed intake on enteric methane production in lactating dairy cows. J. Anim. Sci. 2017, 95, 3687–3699. [Google Scholar] [CrossRef]
- Breider, I.S.; Wall, E.; Garnsworthy, P.C. Short communication: Heritability of methane production and genetic correlations with milk yield and body weight in Holstein-Friesian dairy cows. J. Dairy Sci. 2019, 102, 7277–7281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassen, J.; Løvendahl, P. Heritability estimates for enteric methane emissions from Holstein cattle measured using noninvasive methods. J. Dairy Sci. 2016, 99, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Haas, Y.; Windig, J.J.; Calus, M.P.L.; Dijkstra, J.; de Haan, M.; Bannink, A.; Veerkamp, R.F.; Heath, A.C.; Martin, N.G.; Montgomery, G.W.; et al. Genetic parameters for predicted methane production and potential for reducing enteric emissions through genomic selection. J. Dairy Sci. 2011, 94, 6122–6134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, D.J.; Cánovas, A.; Chud, T.C.S.; Cant, J.P.; Osborne, V.R.; Baes, C.F.; Schenkel, F.S.; Miglior, F.M. The dynamic behavior of feed efficiency in primiparous dairy cattle. J. Dairy Sci. 2020, 103, 1528–1540. [Google Scholar] [CrossRef]
- Gerrits, W.; Labussière, E.; Dijkstra, J.; Reynolds, C.; Metges, C.; Kuhla, B.; Lund, P.; Weisbjerg, M.R. Letter to the editor: Recovery test results as a prerequisite for publication of gaseous exchange measurements. J. Anim. Sci. 2017, 95, 5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailemariam, D.; Manafiazar, G.; Basarab, J.; Stothard, P.; Miglior, F.; Plastow, G.; Wang, Z.; Hailemariam, D.; Stothard, P.; Plastow, G.; et al. Comparative analyses of enteric methane emissions, dry matter intake, and milk somatic cell count in different residual feed intake categories of dairy cows. Can. J. Anim. Sci. 2021, 101, 177–190. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 30 May 2018).
- Herd, R.M.; Bird, S.H.; Donoghue, K.A.; Arthur, P.F.; Hegarty, R.F. Phenotypic associations between methane productions traits, volatile fatty acids and animal breeding traits. In Proceedings of the Twentieth Conference of the Association for the Advancement of Animal Breeding and Genetics, Translating Science into Action, Napier, New Zealand, 20–23 October 2013. [Google Scholar]
- Brito, L.F.; Schenkel, F.S.; Oliveira, H.R.; Cánovas, A.; Miglior, F. Meta-analysis of heritability estimates for methane emission indicator traits in cattle and sheep. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 7–11 February 2018. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Welham, S.J.; Thompson, R. ASReml User Guide Release 4.1 Structural Specification; VSN International Ltd.: Indore, India, 2015; pp. 1–375. [Google Scholar]
- Roso, V.M.; Schenkel, F.S. AMC—A computer program to assess the degree of connectedness among contemporary groups. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Brazil, 13–18 August 2006; pp. 26–27. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman Group Limited: Essex, England, 1996. [Google Scholar]
- Meuwissen, T.H.E.; Luo, Z. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 1992, 24, 305–313. [Google Scholar] [CrossRef]
- Ghavi Hossein-Zadeh, N. Estimates of the genetic contribution to methane emission in dairy cows: A meta-analysis. Sci. Rep. 2022, 12, 12352. [Google Scholar] [CrossRef]
- Grainger, C.; Clarke, T.; McGinn, S.M.; Auldist, M.J.; Beauchemin, K.A.; Hannah, M.C.; Waghorn, G.C.; Clark, H.; Eckard, R.J. Methane emissions from dairy cows measured using the sulfur hexafluoride (SF6) tracer and chamber techniques. J. Dairy Sci. 2007, 90, 2755–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denninger, T.M.; Schwarm, A.; Dohme-Meier, F.; Münger, A.; Bapst, B.; Wegmann, S.; Grandl, F.; Vanlierde, A.; Sorg, D.; Ortmann, S.; et al. Accuracy of methane emissions predicted from milk mid-infrared spectra and measured by laser methane detectors in Brown Swiss dairy cows. J. Dairy Sci. 2020, 103, 2024–2039. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Kebreab, E.; Hristov, A.N.; Oh, J.; Arndt, C.; Bannink, A.; Bayat, A.R.; Brito, A.F.; Boland, T.; Casper, D.; et al. Prediction of enteric methane production, yield, and intensity in dairy cattle using an intercontinental database. Glob. Change Biol. 2018, 24, 3368–3389. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.; Weeks, H.; Zimmerman, P.R.; Harper, M.T.; Hristova, R.A.; Zimmerman, R.; Branco, A.F. The Use of an Automated System (GreenFeed) to Monitor Enteric Methane and Carbon Dioxide Emissions from Ruminant Animals. J. Vis. Exp. 2015, 103, e52904. [Google Scholar] [CrossRef] [Green Version]
- Pszczola, M.; Rzewuska, K.; Mucha, S.; Strabel, T. Heritability of methane emissions from dairy cows over a lactation measured on commercial farms. J. Anim. Sci. 2017, 95, 4813–4819. [Google Scholar] [CrossRef]
- van Engelen, S.; Bovenhuis, H.; van der Tol, P.P.J.; Visker, M.H.P.W. Genetic background of methane emission by Dutch Holstein Friesian cows measured with infrared sensors in automatic milking systems. J. Dairy Sci. 2018, 101, 2226–2234. [Google Scholar] [CrossRef] [Green Version]
- Chagunda, M.G.G.; Ross, D.; Roberts, D.J. On the use of a laser methane detector in dairy cows. Comput. Electron. Agric. 2009, 68, 157–160. [Google Scholar] [CrossRef]
- Manzanilla-Pech, C.I.V.; De Haas, Y.; Hayes, B.J.; Veerkamp, R.F.; Khansefid, M.; Donoghue, K.A.; Arthur, P.F.; Pryce, J.E. Genomewide association study of methane emissions in Angus beef cattle with validation in dairy cattle. J. Anim. Sci. 2016, 94, 4151–4166. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. Repeatability for Gaussian and non-Gaussian data: A practical guide for biologists. Biol. Rev. 2010, 85, 935–956. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.J.; Potterton, S.L.; Craigon, J.; Saunders, N.; Wilcox, R.H.; Hunter, M.; Goodman, J.R.; Garnsworthy, P.C. Variation in enteric methane emissions among cows on commercial dairy farms. Animal 2014, 8, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.N.; Cornou, C.; Madsen, J. Individual variation and repeatability of methane production from dairy cows estimated by the CO2 method in automatic milking system. Animal 2015, 9, 1567–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, M.S. Effects of Diet on Short-Term Regulation of Feed Intake by Lactating Dairy Cattle. J. Dairy Sci. 2000, 83, 1598–1624. [Google Scholar] [CrossRef] [PubMed]
- Michael Forbes, J. A personal view of how ruminant animals control their intake and choice of food: Minimal total discomfort. Nutr. Res. Rev. 2007, 20, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.S.; Bradford, B.J.; Oba, M. Board Invited Review: The hepatic oxidation theory of the control of feed intake and its application to ruminants. J. Anim. Sci. 2009, 87, 3317–3334. [Google Scholar] [CrossRef] [Green Version]
- Negussie, E.; Mehtiö, T.; Mäntysaari, P.; Løvendahl, P.; Mäntysaari, E.A.; Lidauer, M.H. Reliability of breeding values for feed intake and feed efficiency traits in dairy cattle: When dry matter intake recordings are sparse under different scenarios. J. Dairy Sci. 2019, 102, 7248–7262. [Google Scholar] [CrossRef] [Green Version]
- Byskov, M.V.; Fogh, A.; Løvendahl, P. Genetic parameters of rumination time and feed efficiency traits in primiparous Holstein cows under research and commercial conditions. J. Dairy Sci. 2017, 100, 9635–9642. [Google Scholar] [CrossRef]
- Li, B.; Fikse, W.F.; Løvendahl, P.; Lassen, J.; Lidauer, M.H.; Mäntysaari, P.; Berglund, B. Genetic heterogeneity of feed intake, energy-corrected milk, and body weight across lactation in primiparous Holstein, Nordic Red, and Jersey cows. J. Dairy Sci. 2018, 101, 10011–10021. [Google Scholar] [CrossRef] [PubMed]
- Krattenmacher, N.; Thaller, G.; Tetens, J. Analysis of the genetic architecture of energy balance and its major determinants dry matter intake and energy-corrected milk yield in primiparous Holstein cows. J. Dairy Sci. 2019, 102, 3241–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, D.J.; Cánovas, A.; Chud, T.C.S.; Cant, J.P.; Osborne, V.R.; Baes, C.F.; Schenkel, F.S.; Miglior, F. Associations between feed efficiency and aspects of lactation curves in primiparous Holstein dairy cattle. J. Dairy Sci. 2021, 104, 9304–9315. [Google Scholar] [CrossRef] [PubMed]
- Pickering, N.K.; Oddy, V.H.; Basarab, J.; Cammack, K.; Hayes, B.; Hegarty, R.S.; Lassen, J.; McEwan, J.C.; Miller, S.; Pinares-Patino, C.S.; et al. Animal board invited review: Genetic possibilities to reduce enteric methane emissions from ruminants. Animal 2015, 9, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- van Breukelen, A.E.; Aldridge, M.A.; Veerkamp, R.F.; de Haas, Y. Genetic parameters for repeatedly recorded entereic methane concentrations of dairy cows. J. Dairy Sci. 2022, 105, 4256–4271. [Google Scholar] [CrossRef]
- Kandel, P.B.; Vanrobays, M.L.; Vanlierde, A.; Dehareng, F.; Froidmont, E.; Gengler, N.; Soyeurt, H. Genetic parameters of mid-infrared methane predictions and their relationships with milk production traits in Holstein cattle. J. Dairy Sci. 2017, 100, 5578–5591. [Google Scholar] [CrossRef]
- Donoghue, K.A.; Bird-Gardiner, T.; Arthur, P.F.; Herd, R.M.; Hegarty, R.F. Genetic and phenotypic variance and covariance components for methane emission and postweaning traits in Angus cattle. J. Anim. Sci. 2016, 94, 1438. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Herd | Number of Cows | Number of Records | Mean | SD | Min | Max | CV (%) |
|---|---|---|---|---|---|---|---|---|
| MeP (g/day) | DRTC 2 | 58 | 433 | 355.6 | 92.2 | 138 | 674 | 26 |
| ODRC 3 | 272 | 1332 | 498.6 | 99.4 | 224 | 799 | 20 | |
| Total | 330 | 1765 | 463.5 | 115.4 | 135 | 799 | 25 | |
| MeY (g/kg DMI 1) | DRTC 2 | 54 | 342 | 17.2 | 4.5 | 5.1 | 34.4 | 26 |
| ODRC 3 | 233 | 1126 | 25.4 | 5.7 | 11.7 | 75.7 | 22 | |
| Total | 287 | 1468 | 23.5 | 6.4 | 5.1 | 75.7 | 27 | |
| MeI (g/kg milk) | DRTC 2 | 21 | 176 | 10.6 | 3.7 | 4.1 | 26.2 | 35 |
| ODRC 3 | 244 | 1197 | 15.2 | 3.6 | 6.1 | 29.7 | 24 | |
| Total | 265 | 1373 | 14.6 | 3.9 | 4.1 | 29.7 | 27 |
| Herd | DIM | Age | MY | FY | PY | ECM | DMI | BW |
|---|---|---|---|---|---|---|---|---|
| DRTC 1 | 132 (73) | 24.4 (2.0) | 32.65 (6.57) | 1.214 (0.297) | 1.037 (0.204) | 30.96 (6.10) | 20.44 (3.60) | 259 (30) |
| ODRC 2 | 140 (85) | 24.5 (1.6) | 30.85 (5.23) | 1.241 (0.222) | 1.007 (0.158) | 30.62 (4.65) | 19.04 (5.02) | 295 (28) |
| Trait | h2 (SE) | r (SE) | Average EBVAccuracy | |||
|---|---|---|---|---|---|---|
| MeP 1 | 1147.3 | 2735.6 | 3279.6 | 0.16 (0.10) | 0.54 (0.03) | 0.36 |
| MeY 2 | 6.5 | 5.1 | 12.3 | 0.27 (0.12) | 0.49 (0.03) | 0.44 |
| MeI 3 | 2.5 | 6.2 | 3.1 | 0.21 (0.14) | 0.74 (0.02) | 0.32 |
| Trait | MeP | MeY | MeI |
|---|---|---|---|
| MeP | - | 0.73 (0.26) | 0.94 (0.23) |
| MeY | 0.67 (0.02) | - | 0.68 (0.23) |
| MeI | 0.70 (0.03) | 0.63 (0.03) | - |
| Trait | MeP | MeY | MeI |
|---|---|---|---|
| MeP | - | 0.65 | 0.67 |
| MeY | 0.90 | - | 0.64 |
| MeI | 0.88 | 0.88 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamalanathan, S.; Houlahan, K.; Miglior, F.; Chud, T.C.S.; Seymour, D.J.; Hailemariam, D.; Plastow, G.; de Oliveira, H.R.; Baes, C.F.; Schenkel, F.S. Genetic Analysis of Methane Emission Traits in Holstein Dairy Cattle. Animals 2023, 13, 1308. https://doi.org/10.3390/ani13081308
Kamalanathan S, Houlahan K, Miglior F, Chud TCS, Seymour DJ, Hailemariam D, Plastow G, de Oliveira HR, Baes CF, Schenkel FS. Genetic Analysis of Methane Emission Traits in Holstein Dairy Cattle. Animals. 2023; 13(8):1308. https://doi.org/10.3390/ani13081308
Chicago/Turabian StyleKamalanathan, Stephanie, Kerry Houlahan, Filippo Miglior, Tatiane C. S. Chud, Dave J. Seymour, Dagnachew Hailemariam, Graham Plastow, Hinayah R. de Oliveira, Christine F. Baes, and Flavio S. Schenkel. 2023. "Genetic Analysis of Methane Emission Traits in Holstein Dairy Cattle" Animals 13, no. 8: 1308. https://doi.org/10.3390/ani13081308