
A Bayesian Multivariate Gametic Model in a Reciprocal Cross with Genomic Information: An Example with Two Iberian Varieties
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serra, X.; Gil, F.; Pérez-Enciso, M.; Oliver, M.A.; Vázquez, J.M.; Gispert, M.; Díaz, I.; Moreno, F.; Latorre, R.; Noguera, J.L. A comparison of carcass, meat quality and histochemical characteristics of Iberian (Guadyerbas line) and Landrace pigs. Livest. Prod. Sci. 1998, 56, 215–223. [Google Scholar] [CrossRef]
- Serrano, M.P.; Valencia, D.G.; Nieto, M.; Lázaro, R.; Mateos, G.G. Influence of sex and terminal sire line on performance and carcass and meat quality of Iberian pigs reared under intensive production systems. Meat Sci. 2008, 78, 420–428. [Google Scholar] [CrossRef]
- Silio, L.; Rodriguez, M.C.; Rodrigañez, J.; Toro, M.A. La selección de cerdos ibéricos. In Porcino Ibérico: Aspectos Claves; Buxade, C., Daza, A., Eds.; Mundi-Press: Madrid, Spain, 2001; pp. 125–149. [Google Scholar]
- Fernández, A.; Rodrigáñez, J.; Zuzúarregui, J.; Rodríguez, M.C.; Silió, L. Genetic parameters for litter size and weight at different parities in Iberian pigs. Span. J. Agric. Res. 2008, 6, 98–106. [Google Scholar] [CrossRef]
- García-Casco, J.M.; Fernández, A.; Rodríguez, M.C.; Silió, L. Heterosis for litter size and growth in crosses of four strains of Iberian pig. Livest. Sci. 2012, 147, 1–8. [Google Scholar] [CrossRef]
- Noguera, J.L.; Ibáñez-Escriche, N.; Casellas, J.; Rosas, J.P.; Varona, L. Genetic parameters and direct, maternal and heterosis effects on litter size in a diallel cross among three commercial varieties of Iberian pig. Animal 2019, 13, 2765–2772. [Google Scholar] [CrossRef] [PubMed]
- Reik, W.; Walter, J. Genomic imprinting: Parental influence on the genome. Nat. Rev. Genet. 2001, 2, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Hanna, C.W. Placental imprinting: Emerging mechanisms and functions. PLoS Genet. 2020, 16, e1008709. [Google Scholar] [CrossRef] [PubMed]
- Coster, A.; Madsen, O.; Heuven, H.C.M.; Dibbits, B.; Groenen, M.A.M.; van Arendonk, J.A.M.; Bovenhuis, H. The imprinted gene DIO3 is a candidate gene for litter size in pigs. PLoS ONE 2012, 7, e31825. [Google Scholar] [CrossRef] [PubMed]
- Nishio, M.; Satoh, M. Genomic best linear unbiased prediction method including imprinting effects for genomic evaluation. Genet. Sel. Evol. 2015, 47, 32. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.M.; Kinghorn, B.P.; Tier, B.; Wilson, J.F.; Dunstan, N.; Van Der Werf, J.H.J. A combined long-range phasing and long haplotype imputation method to impute phase for SNP genotypes. Genet. Sel. Evol. 2011, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Srihi, H.; Ibáñez-Escriche, N.; Casellas, J.; Noguera, J.L.; Hernández, P.; Martín de Hijas, M.; Vázquez-Gómez, M.; Negro, S.; Rosas, J.P.; Varona, L. Bayesian analysis of paternal and maternal gametic effects in a reciprocal cross between two Iberian varieties. In Proceedings of the 12th World Congress on Genetics Applied to Livestock Production, Rotterdam, The Netherlands, 3–8 July 2022; pp. 3180–3183. [Google Scholar]
- Gelfand, A.E.; Smith, A.F.M. Sampling-Based Approaches to Calculating Marginal Densities. J. Am. Stat. Assoc. 1990, 85, 398–409. [Google Scholar] [CrossRef]
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family of Programs; University of Georgia: Athens, GA, USA, 2018; p. 142. [Google Scholar]
- Patten, M.M.; Ross, L.; Curley, J.P.; Queller, D.C.; Bonduriansky, R.; Wolf, J.B. The evolution of genomic imprinting: Theories, predictions and empirical tests. Heredity 2014, 113, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.; Haig, D. Genomic imprinting in mammalian development: A parental tug-of-war. Trends Genet. 1991, 7, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Thamban, T.; Agarwaal, V.; Khosla, S. Role of genomic imprinting in mammalian development. J. Biosci. 2020, 45, 20. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Coan, P.M.; Angiolini, E.; Burton, G.J.; Constancia, M. Imprinted genes and the epigenetic regulation of placental phenotype. Prog. Biophys. Mol. Biol. 2011, 106, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Zhao, H.; Li, Y.J.; Khederzadeh, S.; Wei, H.J.; Zhou, Z.Y.; Zhang, Y.P. Genome-wide identification of imprinted genes in pigs and their different imprinting status compared with other mammals. Zool. Res. 2020, 41, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.W.; Han, Z.B.; Deng, C.Y.; He, H.J.; Wu, Q. Conservation of genomic imprinting at the NDN, MAGEL2 and MEST loci in pigs. Genes Genet. Syst. 2012, 87, 53–58. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
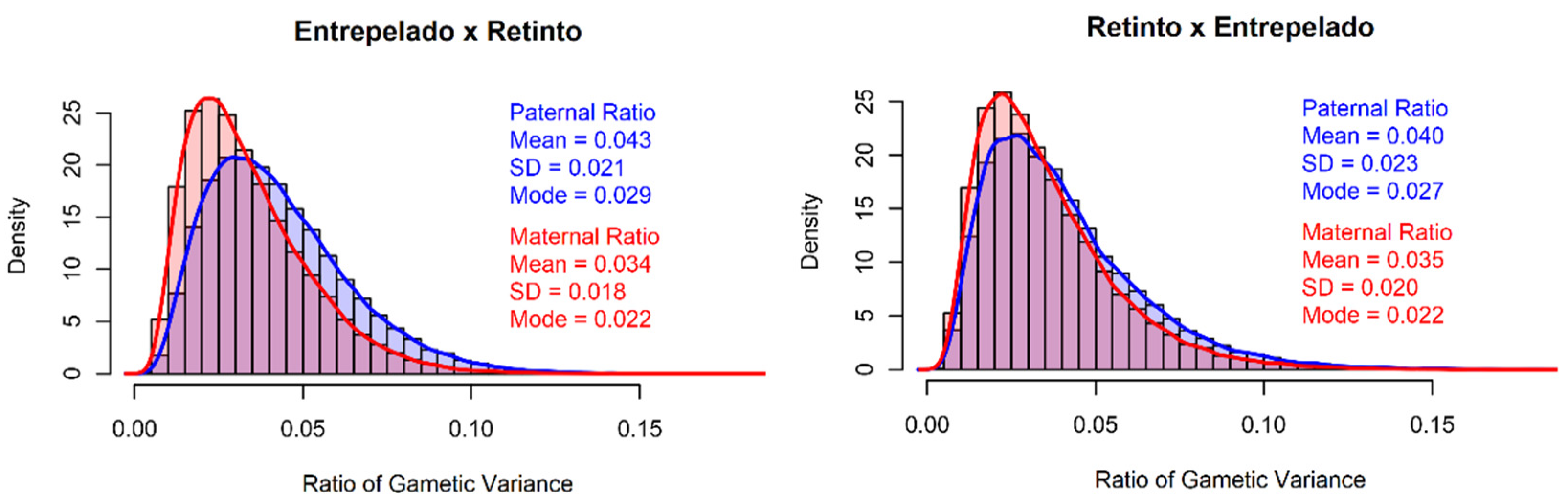
| Entrepelado × Retinto | Retinto × Entrepelado | |
|---|---|---|
| N 1 (NS) 2 | 1258 (203) | 700 (125) |
| TNB 3 | 8.78 ± 2.24 | 8.85 ± 2.37 |
| NBA 4 | 8.55 ± 2.23 | 8.62 ± 2.34 |
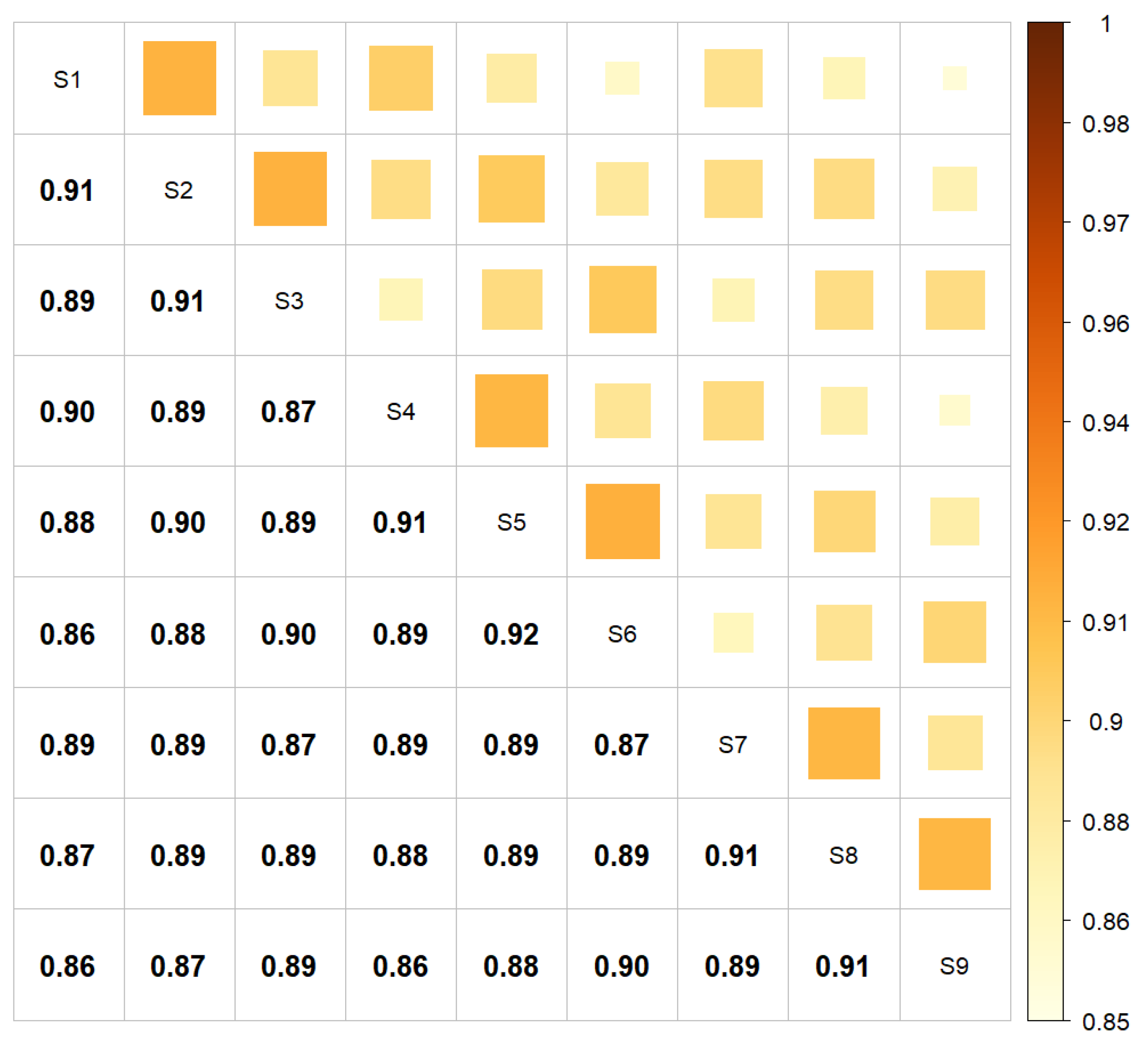
| Scenario | Core Length | Tail Length |
|---|---|---|
| S1 | 75 | 100 |
| S2 | 75 | 150 |
| S3 | 75 | 200 |
| S4 | 100 | 100 |
| S5 | 100 | 150 |
| S6 | 100 | 200 |
| S7 | 125 | 100 |
| S8 | 125 | 150 |
| S9 | 125 | 200 |
| Gametic Relationship | Entrepelado | Retinto |
|---|---|---|
| <0.05 | 92,276 (86.03%) | 94,144 (87.77%) |
| 0.05–0.10 | 8130 (7.58%) | 8900 (8.29%) |
| 0.10–0.20 | 4670 (4.35%) | 3130 (2.92%) |
| 0.20–0.30 | 1076 (1.00%) | 582 (0.54%) |
| 0.30–0.40 | 480 (0.44%) | 252 (0.23%) |
| 0.40–0.50 | 396 (0.36%) | 188 (0.18%) |
| >0.50 | 228 (0.21%) | 60 (0.05%) |
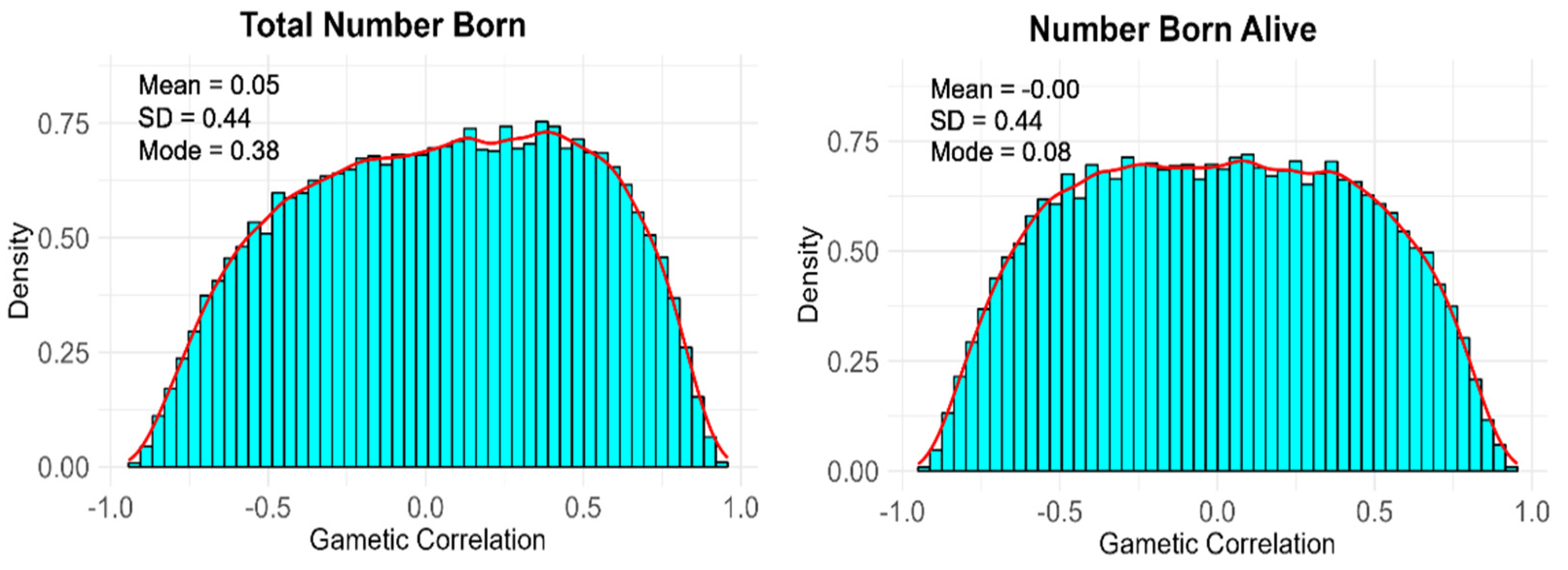
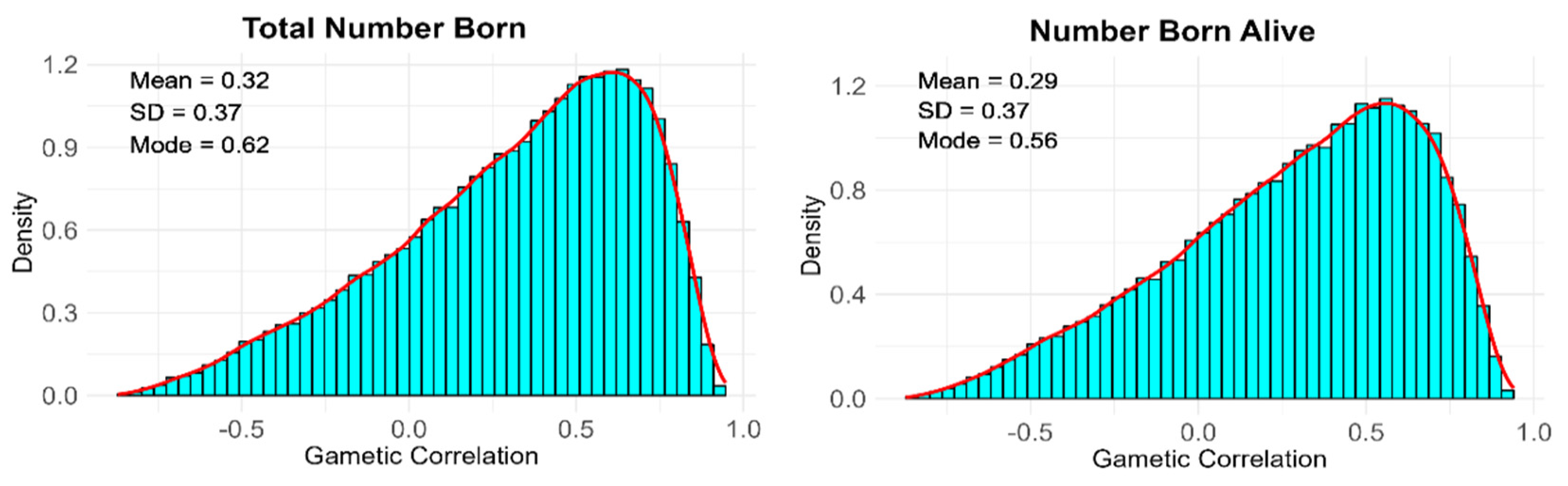
| Variance Component | TNB | NBA |
|---|---|---|
| 0.144 (0.098) | 0.142 (0.097) | |
| 0.357 (0.187) | 0.365 (0.191) | |
| 0.206 (0.103) | 0.199 (0.100) | |
| 0.197 (0.114) | 0.199 (0.115) | |
| 0.224 (0.132) | 0.222 (0.128) | |
| 0.163 (0.087) | 0.151 (0.080) | |
| 4.296 (0.187) | 4.251 (0.185) | |
| 4.795 (0.288) | 4.607 (0.278) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srihi, H.; López-Carbonell, D.; Ibáñez-Escriche, N.; Casellas, J.; Hernández, P.; Negro, S.; Varona, L. A Bayesian Multivariate Gametic Model in a Reciprocal Cross with Genomic Information: An Example with Two Iberian Varieties. Animals 2023, 13, 1648. https://doi.org/10.3390/ani13101648
Srihi H, López-Carbonell D, Ibáñez-Escriche N, Casellas J, Hernández P, Negro S, Varona L. A Bayesian Multivariate Gametic Model in a Reciprocal Cross with Genomic Information: An Example with Two Iberian Varieties. Animals. 2023; 13(10):1648. https://doi.org/10.3390/ani13101648
Chicago/Turabian StyleSrihi, Houssemeddine, David López-Carbonell, Noelia Ibáñez-Escriche, Joaquim Casellas, Pilar Hernández, Sara Negro, and Luis Varona. 2023. "A Bayesian Multivariate Gametic Model in a Reciprocal Cross with Genomic Information: An Example with Two Iberian Varieties" Animals 13, no. 10: 1648. https://doi.org/10.3390/ani13101648