
Effect of Dietary Manganese on the Growth Performance, Lipid Metabolism, and Antioxidant Capacity in the Post-Larval Coho Salmon (Oncorhynchus kisutch)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Formulations
2.2. Feeding Trial
2.3. Sampling Procedures
2.4. Whole-Body Composition Analysis
2.5. Hepatic Enzyme Activity Analysis
2.6. Real-Time Quantitative Polymerase Chain Reaction
2.7. Calculations and Statistical Analysis
3. Results
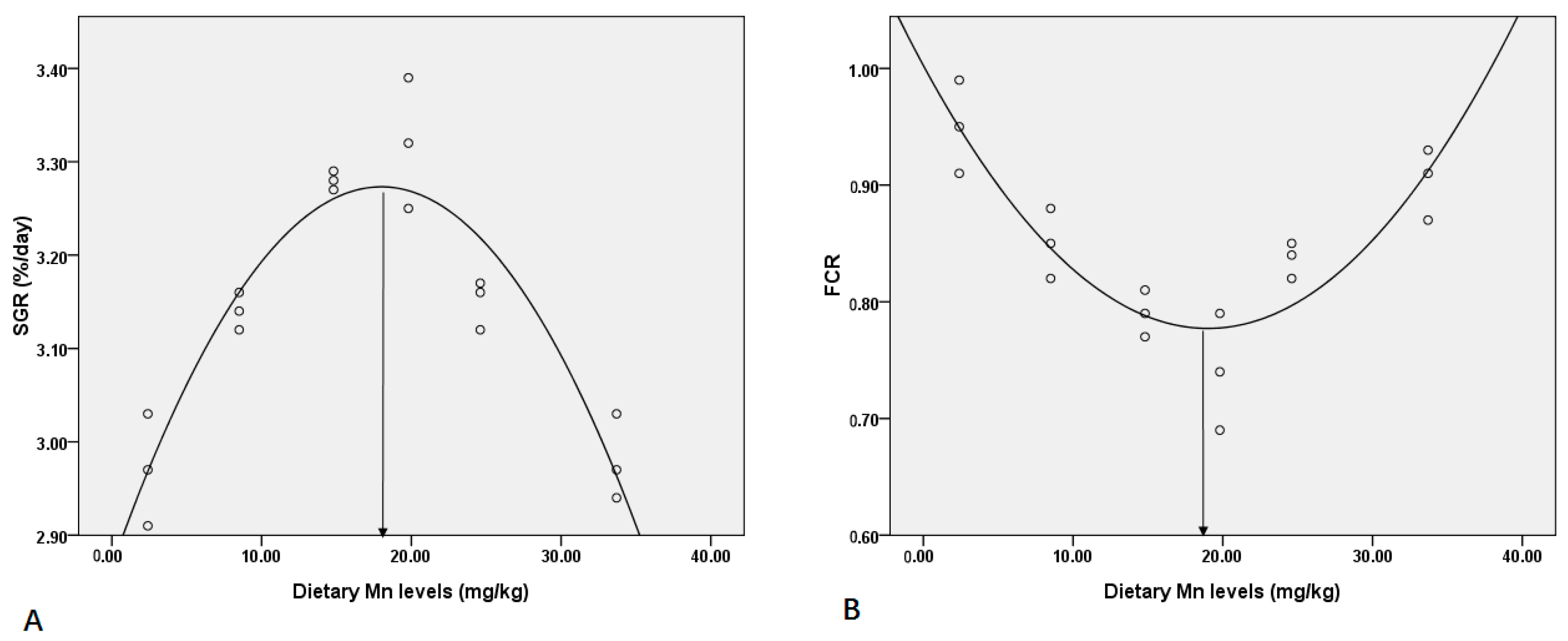
3.1. Effect of Dietary Mn on Feed Utilization and Growth Performance
3.2. Effect of Dietary Mn on the Whole-Body Biochemical Composition
3.3. Effect of Dietary Mn on the Vertebral and Whole-Body Mn Content
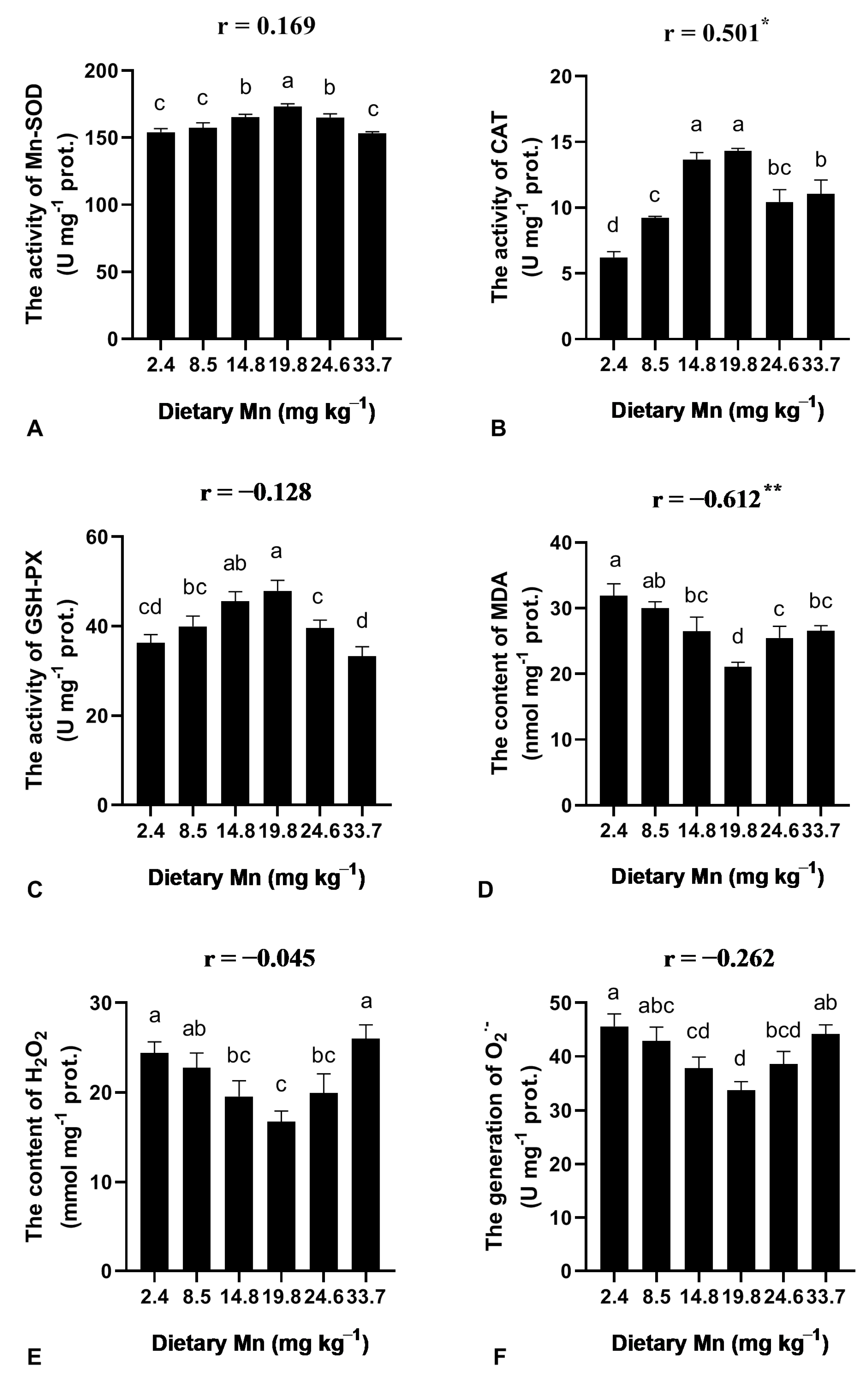
3.4. Effect of Dietary Mn on the Activities of Antioxidant Enzymes in the Liver
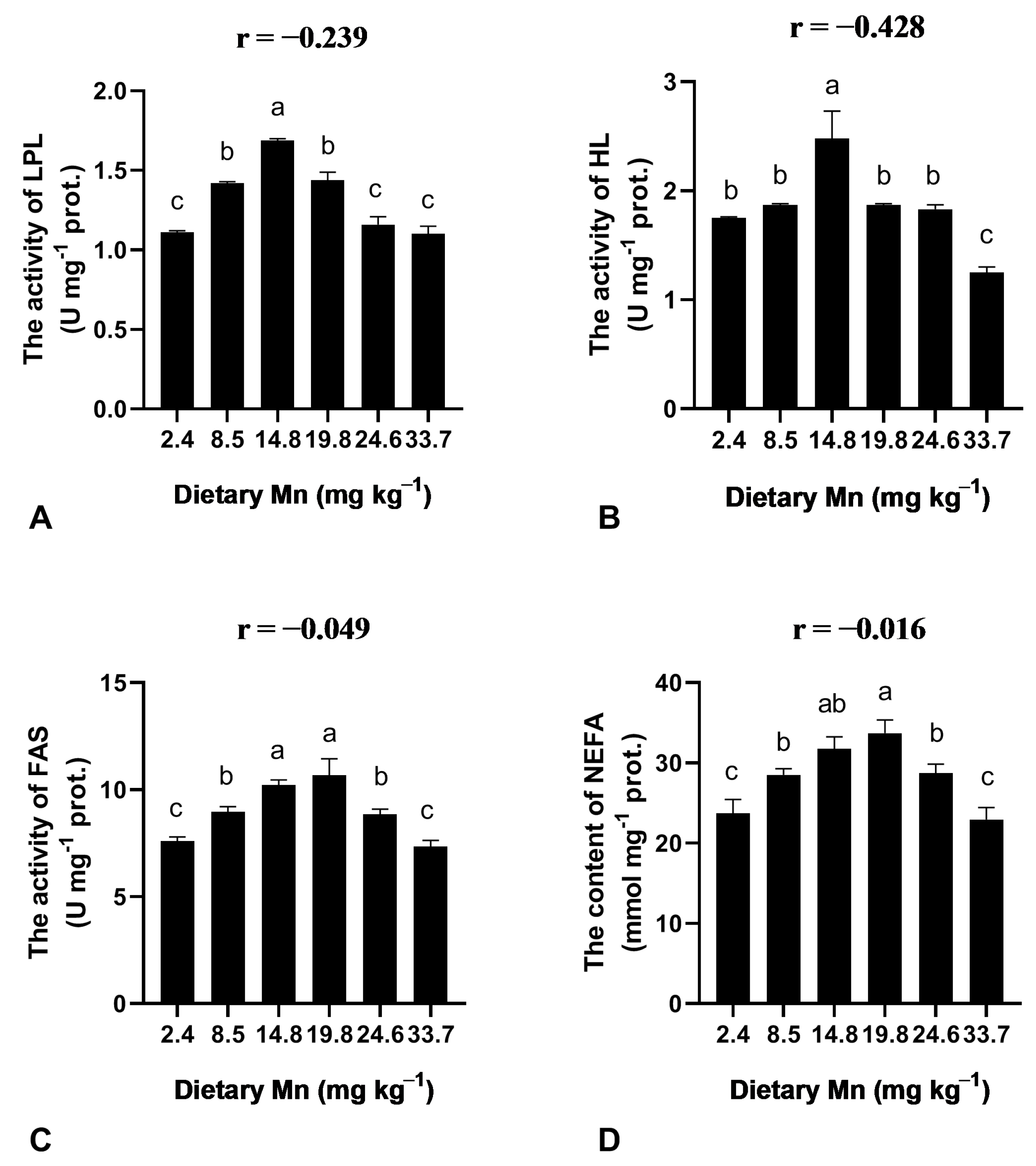
3.5. Effect of Dietary Mn on the Activities of Lipid Metabolism Enzymes in the Liver
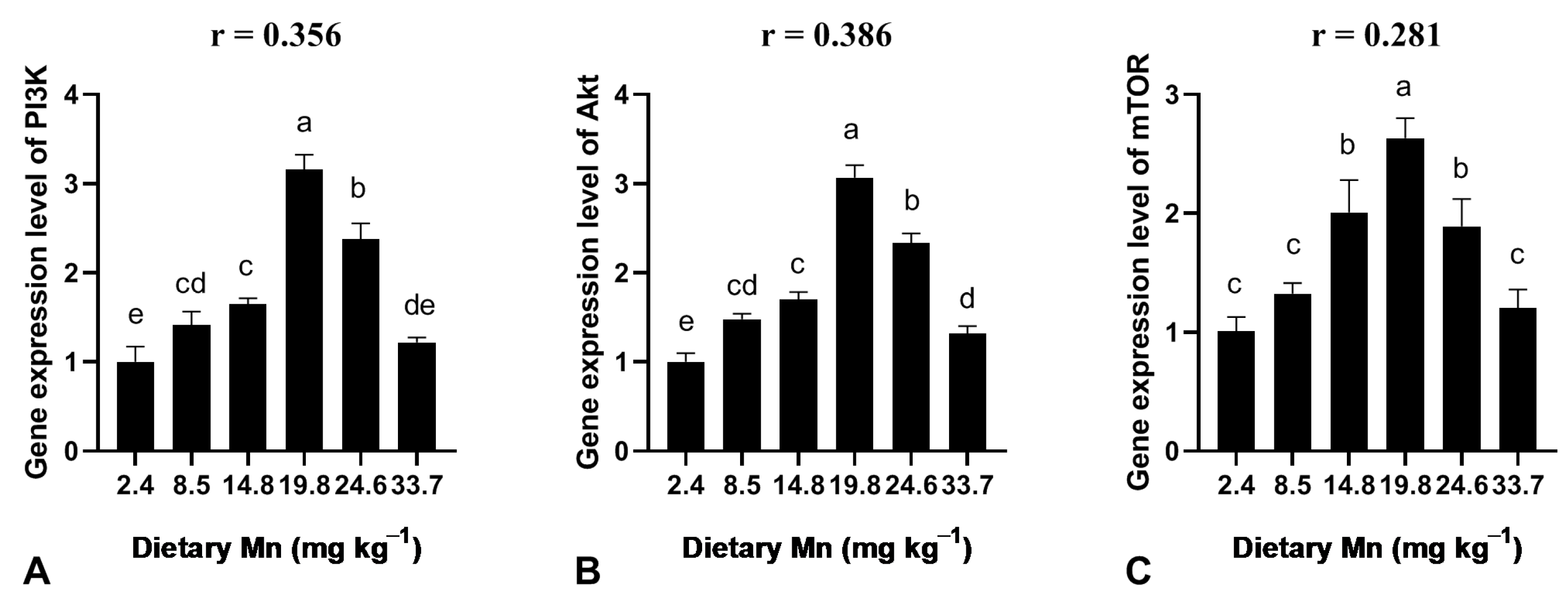
3.6. Effect of Dietary Mn on the Gene Expression Level in the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Viegas, M.N.; Salgado, M.A.; Aguiar, C.; Almeida, A.; Gavaia, P.; Dias, J. Effect of Dietary Manganese and Zinc Levels on Growth and Bone Status of Senegalese Sole (Solea senegalensis) Post-Larvae. Biol. Trace Elem. Res. 2021, 199, 2012–2021. [Google Scholar] [CrossRef]
- Palmieri, G.; Arciello, S.; Bimonte, M.; Carola, A.; Tito, A.; Gogliettino, M.; Cocca, E.; Fusco, C.; Balestrieri, M.; Colucci, M.G.; et al. The extraordinary resistance to UV radiations of a manganese superoxide dismutase of Deinococcus radiodurans offers promising potentialities in skin care applications. J. Biotechnol. 2019, 302, 101–111. [Google Scholar] [CrossRef]
- Gatlin, D.M., III; Wilson, R.P. Studies on the manganese requirement of fingerling channel catfish. Aquaculture 1984, 41, 85–92. [Google Scholar] [CrossRef]
- de Rosa, G.; Keen, C.L.; Leach, R.M.; Hurley, L.S. Regulation of superoxide dismutase activity by dietary manganese. J. Nutr. 1980, 110, 795–804. [Google Scholar] [CrossRef]
- Pan, L.; Zhu, X.; Xie, S.; Lei, W.; Han, D.; Yang, Y. Effects of dietary manganese on growth and tissue manganese concentrations of juvenile gibel carp, Carassius auratus gibelio. Aquac. Nutr. 2008, 14, 459–463. [Google Scholar] [CrossRef]
- Liang, J.J.; Wang, S.; Han, B.; Tian, L.X.; Yang, H.J.; Liu, Y.J. Dietary manganese requirement of juvenile grass carp (C tenopharyngodon idella V al.) based on growth and tissue manganese concentration. Aquac. Res. 2015, 46, 2991–2998. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Li, B.; Qiao, H.; Liu, X.; Hao, T.; Wang, X. Dietary manganese requirement of juvenile hybrid grouper, Epinephelus lanceolatus× E. fuscoguttatus. Aquac. Nutr. 2018, 24, 215–223. [Google Scholar] [CrossRef]
- Ye, C.X.; Tian, L.X.; Yang, H.J.; Liang, J.J.; Niu, J.; Liu, Y.J. Growth performance and tissue mineral content of juvenile grouper (Epinephelus coioides) fed diets supplemented with various levels of manganese. Aquac. Nutr. 2009, 15, 608–614. [Google Scholar] [CrossRef]
- Li, A.; Zhang, D.; Wei, W. A study on the nutritional requirement for juvenile flounder Paralichthys olivaceus. J. Zhejiang Ocean Univ. 2001, 20, 6–10. [Google Scholar]
- Tan, L.N.; Feng, L.; Liu, Y.; Jiang, J.; Jiang, W.D.; Hu, K.; Li, S.H.; Zhou, X.Q. Growth, body composition and intestinal enzyme activities of juvenile Jian carp (Cyprinus carpio var. Jian) fed graded levels of dietary zinc. Aquac. Nutr. 2011, 17, 338–345. [Google Scholar] [CrossRef]
- Gonzalez, R.; Grippo, R.; Dunson, W. The disruption of sodium balance in brook charr, Salvelinus fontinalis (Mitchill), by manganese and iron. J. Fish Biol. 1990, 37, 765–774. [Google Scholar] [CrossRef]
- Barnhoorn, I.; Van Vuren, J.H.J.; du Preez, H.H. Sublethal effects of manganese on the carbohydrate metabolism of Oreochromis mossambicus after acute and chronic exposure. S. Afr. J. Zool. 1999, 34, 102–107. [Google Scholar] [CrossRef]
- Friesen, E.N.; Higgs, D.A.; Devlin, R.H. Flesh nutritional content of growth hormone transgenic and non-transgenic coho salmon compared to various species of farmed and wild salmon. Aquaculture 2015, 437, 318–326. [Google Scholar] [CrossRef]
- Du, L.-c.; Yu, H.-r.; Li, L.-y.; Zhang, Q.; Tian, Q.; Liu, J.-q.; Shan, L.-l. Dietary selenium requirement of coho salmon (Oncorhynchus kisutch W.) alevins. Aquac. Int. 2021, 29, 2291–2304. [Google Scholar] [CrossRef]
- Song, J.; Li, L.-Y.; Chen, B.-B.; Shan, L.-L.; Yuan, S.-Y.; Yu, H.-R. Dietary copper requirements of postlarval coho salmon (Oncorhynchus kisutch). Aquac. Nutr. 2021, 27, 2084–2092. [Google Scholar] [CrossRef]
- Yu, H.; Chen, B.; Li, L.; Zhang, Q.; Liu, D.; Wang, Z.; Shan, L. Dietary iron (Fe) requirement of coho salmon (Oncorhynchus kisutch) alevins assessed using growth, whole body and hepatic Fe concentrations and hepatic antioxidant enzymes activities. Aquac. Res. 2021, 52, 4489–4497. [Google Scholar] [CrossRef]
- Liu, D.D.; Han, C.C.; Wan, H.F.; He, F.; Xu, H.Y.; Wei, S.H.; Du, X.H.; Xu, F. Effects of inhibiting PI3K-Akt-mTOR pathway on lipid metabolism homeostasis in goose primary hepatocytes. Animal 2016, 10, 1319–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chen, J.; Huang, J.; Li, Z.; Gong, Y.; Zou, B.; Liu, X.; Ding, L.; Li, P.; Zhu, Z.; et al. HIF-2α upregulation mediated by hypoxia promotes NAFLD-HCC progression by activating lipid synthesis via the PI3K-AKT-mTOR pathway. Aging 2019, 11, 10839–10860. [Google Scholar] [CrossRef]
- Han, C.; Wei, S.; He, F.; Liu, D.; Wan, H.; Liu, H.; Li, L.; Xu, H.; Du, X.; Xu, F. The Regulation of Lipid Deposition by Insulin in Goose Liver Cells Is Mediated by the PI3K-AKT-mTOR Signaling Pathway. PLoS ONE 2015, 10, e0098759. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Xue, Y.; Zheng, B.; Li, L.; Chu, X.; Zhao, Y.; Wu, Y.; Zhang, J.; Han, X.; Wu, Z.; et al. Liquiritigenin protects against arsenic trioxide-induced liver injury by inhibiting oxidative stress and enhancing mTOR-mediated autophagy. Biomed. Pharmacother. 2021, 143, 112167. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemists: Official Methods of Analysis; AOAC: Rockville, MD, USA, 1995. [Google Scholar]
- Tang, R.J.; Feng, L.; Jiang, W.D.; Liu, Y.; Kuang, S.Y.; Jiang, J.; Zhang, Y.A.; Tang, L.; Zhou, X.Q. Growth, digestive and absorptive abilities and antioxidative capacity in the hepatopancreas and intestine of young grass carp (Ctenopharyngodon idellus Val.) fed graded levels of dietary manganese. Aquac. Res. 2016, 47, 1917–1931. [Google Scholar] [CrossRef]
- Tan, X.-Y.; Xie, P.; Luo, Z.; Lin, H.-Z.; Zhao, Y.-H.; Xi, W.-Q. Dietary manganese requirement of juvenile yellow catfish Pelteobagrus fulvidraco, and effects on whole body mineral composition and hepatic intermediary metabolism. Aquaculture 2012, 326, 68–73. [Google Scholar] [CrossRef]
- Liu, K.; Ai, Q.; Mai, K.; Zhang, W.; Zhang, L.; Zheng, S. Dietary manganese requirement for juvenile cobia, Rachycentron canadum L. Aquac. Nutr. 2013, 19, 461–467. [Google Scholar] [CrossRef]
- Maage, A.; Lygren, B.; El-Mowafi, A.F.A. Manganese requirement of Atlantic salmon (Salmo salar) fry. Fish. Sci. 2000, 66, 1–8. [Google Scholar] [CrossRef]
- Serrini, G.; Zhang, Z.; Wilson, R.P. Dietary riboflavin requirement of fingerling channel catfish (Ictalurus punctatus). Aquaculture 1996, 139, 285–290. [Google Scholar] [CrossRef]
- Lorentzen, M.; Maage, A.; Julshamn, K. Manganese supplementation of a practical, fish meal based diet for Atlantic salmon parr. Aquac. Nutr. 1996, 2, 121–125. [Google Scholar] [CrossRef]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, R.; Xu, W.; Zhou, H.; Zhang, W.; Mai, K. Dietary manganese requirement of juvenile large yellow croaker Larimichthys crocea (Richardson, 1846). Aquaculture 2016, 450, 74–79. [Google Scholar] [CrossRef]
- Ji, H.; Li, J.; Liu, P. Regulation of growth performance and lipid metabolism by dietary n-3 highly unsaturated fatty acids in juvenile grass carp, Ctenopharyngodon idellus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 159, 49–56. [Google Scholar] [CrossRef]
- Mourente, G.; Bell, J.G.; Tocher, D.R. Does dietary tocopherol level affect fatty acid metabolism in fish? Fish Physiol. Biochem. 2007, 33, 269–280. [Google Scholar] [CrossRef]
- Malecki, E.A.; Greger, J.L. Manganese protects against heart mitochondrial lipid peroxidation in rats fed high levels of polyunsaturated fatty acids. J. Nutr. 1996, 126, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, W.-N.; Wang, A.-L.; Wang, J.-M.; Sun, R.-Y. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei (Boone, 1931) exposed to acute salinity changes. Aquaculture 2007, 265, 351–358. [Google Scholar] [CrossRef]
- Zhang, J.; Shen, H.; Wang, X.; Wu, J.; Xue, Y. Effects of chronic exposure of 2,4-dichlorophenol on the antioxidant system in liver of freshwater fish Carassius auratus. Chemosphere 2004, 55, 167–174. [Google Scholar] [CrossRef]
- Tian, J.; Chen, J.; Jiang, D.; Liao, S.; Wang, A. Transcriptional regulation of extracellular copper zinc superoxide dismutase from white shrimp Litopenaeus vannamei following Vibrio alginolyticus and WSSV infection. Fish Shellfish Immunol. 2011, 30, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, W.B.; Li, H.Y.; Xu, W.N.; He, J.X.; Li, X.F.; Jiang, G.Z. Effects of dietary supplementation of fructooligosaccharide on growth performance, body composition, intestinal enzymes activities and histology of blunt snout bream (Megalobrama amblycephala) fingerlings. Aquac. Nutr. 2013, 19, 886–894. [Google Scholar] [CrossRef]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/mTOR interactive pathway. Mol. BioSystems 2015, 11, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g) | Dietary Mn Levels (mg kg−1) | |||||
|---|---|---|---|---|---|---|
| 2.4 | 8.5 | 14.8 | 19.8 | 24.6 | 33.7 | |
| Casein 1 | 400.0 | 400.0 | 400.0 | 400.0 | 400.0 | 400.0 |
| Gelatine 1 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Dextrin 1 | 160.0 | 160.0 | 160.0 | 160.0 | 160.0 | 160.0 |
| α-Cellulose 1 | 95.0 | 95.0 | 95.0 | 95.0 | 95.0 | 95.0 |
| Fish oil 1 | 75.0 | 75.0 | 75.0 | 75.0 | 75.0 | 75.0 |
| Soybean oil 1 | 75.0 | 75.0 | 75.0 | 75.0 | 75.0 | 75.0 |
| Mineral premix, manganese-free 2 | 60.0 | 60.0 | 60.0 | 60.0 | 60.0 | 60.0 |
| Vitamin premix 3 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| L-Arg | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| Ethoxyquin | 6.0 | 6.0 | 6.0 | 6.0 | 6.0 | 6.0 |
| DL-Met | 5.0 | 5.0 | 5.0 | 5.0 | 5.0 | 5.0 |
| Choline chloride | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 |
| Ascorbic acid phosphate | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Glycine betaine | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Manganese sulfate (mg kg−1) | 0.0 | 43.0 | 55.3 | 61.5 | 73.7 | 86.0 |
| Proximate composition | ||||||
| Crude protein (%) | 42.1 | 42.08 | 42.04 | 42.06 | 42.03 | 42.05 |
| Crude lipid (%) | 12.57 | 12.53 | 12.55 | 12.57 | 12.59 | 12.51 |
| Ash (%) | 6.12 | 6.18 | 6.13 | 6.17 | 6.15 | 6.19 |
| Moisture (%) | 7.3 | 7.31 | 7.29 | 7.32 | 7.28 | 7.33 |
| Mn (mg kg−1) | 2.4 | 8.5 | 14.8 | 19.8 | 24.6 | 33.7 |
| Target Gene | Forward (5′–3′) | Reverse (5′–3′) | GenBank |
|---|---|---|---|
| PI3K | CCAGTGGCTCAAGGACAAGAACAG | GGATGAAGGTGGCTACGCAGTATC | XM_020466892.1 |
| AKT | GAGTTCACGGCACAGACCATCAC | CGTATGCTGGCGGAGTAAGAGAAC | XM_020503531.1 |
| mTOR | GCAACAGCGACAGCGAGGTAG | TGGAGAGGGAGATTGAGCGGAAG | XM_020506200.1 |
| EF1α | ACCGGCCATCTGATCTACAAATGC | CTCACGCTCAGCCTTCAGCTT | XM_031793751.1 |
| Dietary Mn (mg kg−1) | SGR (% Day−1) | CF (%) | HI (%) | VSI (%) | FCR | PER |
|---|---|---|---|---|---|---|
| 2.4 | 2.97 ± 0.06 c | 0.95 ± 0.05 b | 1.86 ± 0.05 a | 1.43 ± 0.07 a | 0.95 ± 0.04 a | 0.025 ± 0.001 d |
| 8.5 | 3.14 ± 0.02 b | 1.02 ± 0.02 ab | 1.66 ± 0.02 a | 1.48 ± 0.03 a | 0.85 ± 0.03 bc | 0.029 ± 0.001 cd |
| 14.8 | 3.28 ± 0.01 a | 1.02 ± 0.04 ab | 1.64 ± 0.15 a | 1.49 ± 0.38 a | 0.79 ± 0.02 cd | 0.031 ± 0.001 bc |
| 19.8 | 3.32 ± 0.07 a | 1.08 ± 0.08 ab | 1.56 ± 0.01 a | 1.49 ± 0.05 a | 0.74 ± 0.05 d | 0.034±0.001 ab |
| 24.6 | 3.15 ± 0.03 b | 1.11 ± 0.17 ab | 1.52 ± 0.28 a | 1.47 ± 0.39 a | 0.84 ± 0.02 bc | 0.034 ± 0.003 ab |
| 33.7 | 2.98 ± 0.05 c | 1.27 ± 0.15 a | 1.71 ± 0.05 a | 1.34 ± 0.15 a | 0.90 ± 0.03 ab | 0.037 ± 0.002 a |
| ANOVA p-value | 0.000 | 0.034 | 0.094 | 0.962 | 0.000 | 0.000 |
| Linear trend | 0.586 | 0.002 | 0.076 | 0.678 | 0.066 | 0.000 |
| Quadratic trend | 0.000 | 0.311 | 0.021 | 0.425 | 0.000 | 0.170 |
| Dietary Mn Levels (mg kg−1) | Moisture (%) | Crude Protein (%) | Crude Lipid (%) | Ash (%) |
|---|---|---|---|---|
| 2.4 | 75.24 ± 0.02 a | 12.00 ± 0.84 b | 8.45 ± 0.44 cd | 3.49 ± 0.03 a |
| 8.5 | 74.97 ± 0.03 a | 12.04 ± 0.08 b | 8.22 ± 0.01 d | 3.42 ± 0.01 ab |
| 14.8 | 74.60 ± 0.20 ab | 12.37 ± 0.42 b | 8.93 ± 0.05 bc | 3.37 ± 0.02 b |
| 19.8 | 74.20 ± 0.07 b | 12.23 ± 0.74 b | 9.43 ± 0.15 ab | 3.20 ± 0.04 c |
| 24.6 | 72.17 ± 0.08 c | 14.20 ± 0.01 a | 9.85 ± 0.19 a | 3.20 ± 0.04 c |
| 33.7 | 71.96 ± 0.54 c | 14.41 ± 0.28 a | 9.94 ± 0.15 a | 3.18 ± 0.01 c |
| ANOVA p-value | 0.000 | 0.000 | 0.000 | 0.000 |
| Linear trend | 0.000 | 0.000 | 0.000 | 0.000 |
| Quadratic trend | 0.000 | 0.016 | 0.706 | 0.014 |
| Dietary Mn Levels (mg kg−1) | Whole-Body Mn Concentration (mg kg−1) | Vertebral Mn Concentration (mg kg−1) |
|---|---|---|
| 2.4 | 1.23 ± 0.01 e | 10.56 ± 0.17 d |
| 8.5 | 2.59 ± 0.04 d | 15.63 ± 0.15 c |
| 14.8 | 3.59 ± 0.04 c | 23.12 ± 0.39 b |
| 19.8 | 3.91 ± 0.01 a | 24.78 ± 0.48 a |
| 24.6 | 3.92 ± 0.04 a | 23.79 ± 0.16 b |
| 33.7 | 3.68 ± 0.03 b | 23.68 ± 0.20 b |
| ANOVA p-value | 0.000 | 0.000 |
| Linear trend | 0.000 | 0.000 |
| Quadratic trend | 0.000 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Li, L.; Zhang, Q.; Yu, H. Effect of Dietary Manganese on the Growth Performance, Lipid Metabolism, and Antioxidant Capacity in the Post-Larval Coho Salmon (Oncorhynchus kisutch). Animals 2023, 13, 1310. https://doi.org/10.3390/ani13081310
Liu D, Li L, Zhang Q, Yu H. Effect of Dietary Manganese on the Growth Performance, Lipid Metabolism, and Antioxidant Capacity in the Post-Larval Coho Salmon (Oncorhynchus kisutch). Animals. 2023; 13(8):1310. https://doi.org/10.3390/ani13081310
Chicago/Turabian StyleLiu, Dongwu, Lingyao Li, Qin Zhang, and Hairui Yu. 2023. "Effect of Dietary Manganese on the Growth Performance, Lipid Metabolism, and Antioxidant Capacity in the Post-Larval Coho Salmon (Oncorhynchus kisutch)" Animals 13, no. 8: 1310. https://doi.org/10.3390/ani13081310