
Occurrence of Aeromonas Species in the Cutaneous Mucus of Barbour’s Seahorses (Hippocampus barbouri) as Revealed by High-Throughput Sequencing

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
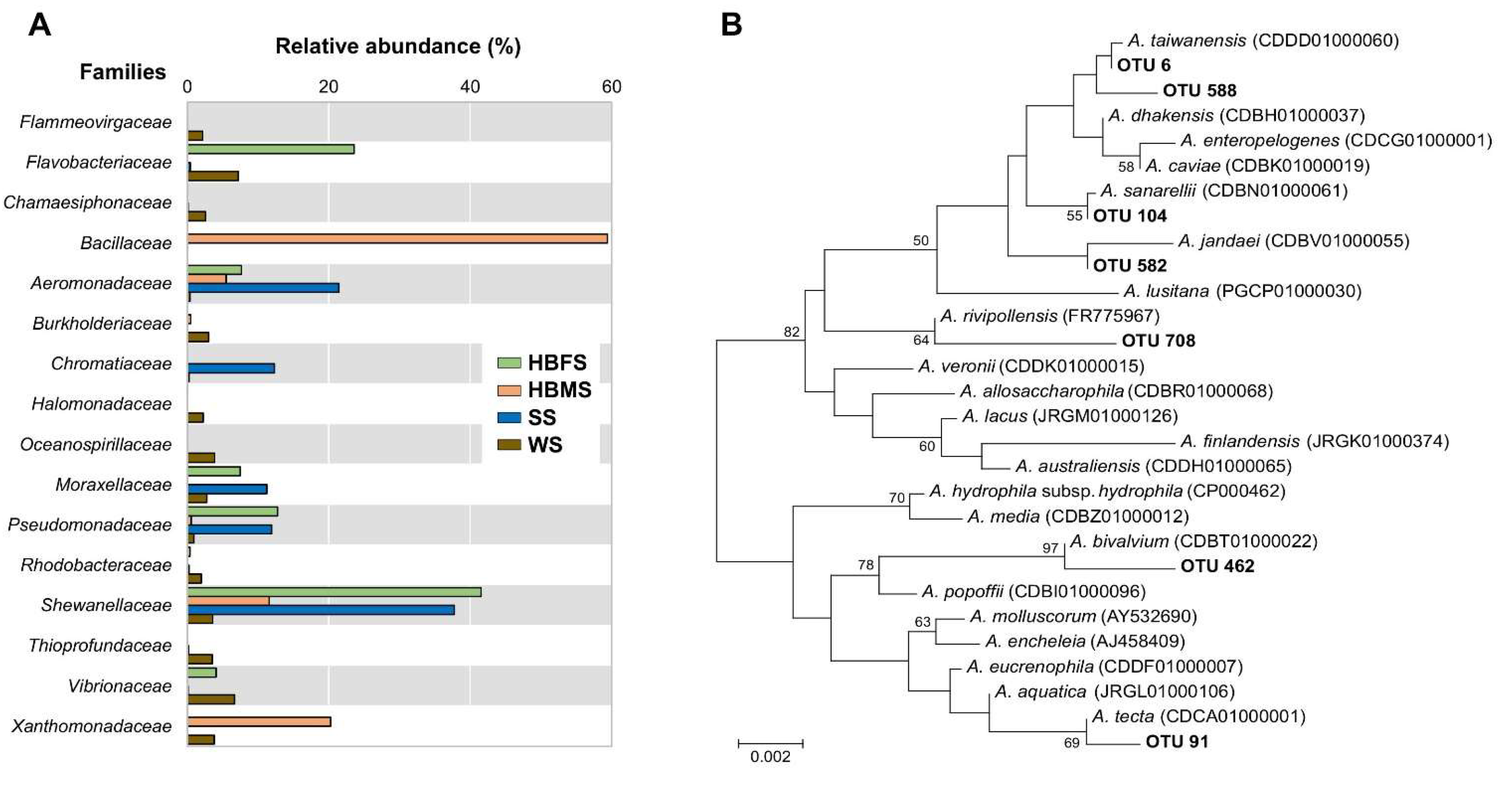
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seahorses in the Philippines. Why Care about Seahorses? Available online: https://static1.squarespace.com/static/5734f2a086db43e3f6fb1bbc/t/5d97c107cfcd403eddf462c1/1570226441051/Why+care+about+seahorses_Philippines_Final+copy.pdf (accessed on 28 May 2021).
- Bereiter-Hahn, J.; Richards, K.S.; Elsner, L.; Voth, M. Composition and formation of flame cell caps: A substratum for the attachment of micro-organisms to sea horse epidermis. Proc. R. Soc. Edinb. B Biol. Sci. 1980, 79, 105–112. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Gallo-Bueno, A.; Planas, M.; Pintado, J. Isolation of Vibrio alginolyticus and Vibrio splendidus from captive-bred seahorses with disease symptoms. Antonie Van Leeuwenhoek 2010, 97, 207–210. [Google Scholar] [CrossRef]
- LePage, V.; Young, J.; Dutton, C.J.; Crawshaw, G.; Paré, J.A.; Kummrow, M.; McLelland, D.J.; Huber, P.; Young, K.; Russell, S.; et al. Diseases of captive yellow seahorse Hippocampus kuda Bleeker, pot-bellied seahorse Hippocampus abdominalis Lesson and weedy seadragon Phyllopteryx taeniolatus (Lacépède). J. Fish Dis. 2015, 38, 439–450. [Google Scholar] [CrossRef]
- Kelly, C.; Salinas, I. Under pressure: Interactions between commensal microbiota and the teleost immune system. Front. Immunol. 2017, 8, 559. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pascual, D.; Vendrell-Fernández, S.; Audrain, B.; Bernal-Bayard, J.; Patiño-Navarrete, R.; Petit, V.; Rigaudeau, D.; Ghigo, J.M. Gnotobiotic rainbow trout (Oncorhynchus mykiss) model reveals endogenous bacteria that protect against Flavobacterium columnare infection. PLoS Pathog. 2021, 17, e1009302. [Google Scholar] [CrossRef]
- Hapfelmeier, S.; Lawson, M.A.; Slack, E.; Kirundi, J.K.; Stoel, M.; Heikenwalder, M.; Cahenzli, J.; Velykoredko, Y.; Balmer, M.L.; Endt, K.; et al. Reversible microbial colonization of germ-free mice reveals the dynamics of IgA immune responses. Science 2010, 328, 1705–1709. [Google Scholar] [CrossRef] [Green Version]
- Gomez de Agüero, M.; Ganal-Vonarburg, S.C.; Fuhrer, T.; Rupp, S.; Uchimura, Y.; Li, H.; Steinert, A.; Heikenwalder, M.; Hapfelmeier, S.; Sauer, U.; et al. The maternal microbiota drives early postnatal innate immune development. Science 2016, 351, 1296–1302. [Google Scholar] [CrossRef]
- Ortega, R.C.M.H.; Tabugo, S.R.M.; Martinez, J.G.T.; Padasas, C.S.; Balolong, M.P.; Balcázar, J.L. High-throughput sequencing-based analysis of bacterial communities associated with Barbour’s seahorses (Hippocampus barbouri) from Surigao del Norte, Philippines. Lett. Appl. Microbiol. 2021, 73, 280–285. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Chaix, G.; Roger, F.; Berthe, T.; Lamy, B.; Jumas-Bilak, E.; Lafite, R.; Forget-Leray, J.; Petit, F. Distinct Aeromonas populations in water column and associated with copepods from estuarine environment (Seine, France). Front. Microbiol. 2017, 8, 1259. [Google Scholar] [CrossRef] [Green Version]
- Chenia, H.Y.; Duma, S. Characterization of virulence, cell surface characteristics and biofilm-forming ability of Aeromonas spp. isolates from fish and sea water. J. Fish Dis. 2017, 40, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Cutuli, M.T.; Gibello, A.; Rodriguez-Bertos, A.; Blanco, M.M.; Villarroel, M.; Giraldo, A.; Guarro, J. Skin and subcutaneous mycoses in tilapia (Oreochromis niloticus) caused by Fusarium oxysporum in coinfection with Aeromonas hydrophila. Med. Mycol. Case Rep. 2015, 9, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Yi, M.; Xiao, P.; Meng, L.; Li, X.; Sun, G.; Liu, Y. The impact of Aeromonas salmonicida infection on innate immune parameters of Atlantic salmon (Salmo salar L). Fish Shellfish Immunol. 2015, 44, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, G.; Hatha, A.A.M.; Lakshmanaperumalsamy, P. Prevalence of Aeromonas hydrophila in fish and prawns from the seafood market of Coimbatore, South India. Food Microbiol. 2005, 22, 133–137. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The gut microbiota of marine fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Ramírez, C.; Romero, J. Fine flounder (Paralichthys adspersus) microbiome showed important differences between wild and reared specimens. Front. Microbiol. 2017, 8, 271. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.J.; Wu, Y.; Yan, Y.Z.; Zou, W.; Xue, J.; Ma, W.R.; Wang, W.; Tian, G.; Wang, L.Y. High-throughput sequencing of microbial community diversity in soil, grapes, leaves, grape juice and wine of grapevine from China. PLoS ONE 2018, 13, e0193097. [Google Scholar] [CrossRef] [Green Version]
- Leite, A.M.O.; Mayo, B.; Rachid, C.T.C.C.; Peixoto, R.S.; Silva, J.T.; Paschoalin, V.M.F.; Delgado, S. Assessment of the microbial diversity of Brazilian kefir grains by PCR-DGGE and pyrosequencing analysis. Food Microbiol. 2012, 31, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Nurul, A.N.A.; Muhammad, D.D.; Okomoda, V.T.; Nur, A.A.B. 16S rRNA-Based metagenomic analysis of microbial communities associated with wild Labroides dimidiatus from Karah Island, Terengganu, Malaysia. Biotechnol. Rep. 2019, 21, e00303. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Gallo-Bueno, A.; Planas, M.; Pintado, J. Identification and characterization of bacteria with antibacterial activities isolated from seahorses (Hippocampus guttulatus). J. Antibiot. 2010, 63, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Beaz-Hidalgo, R.; Figueras, M.J. Aeromonas spp. whole genomes and virulence factors implicated in fish disease. J. Fish Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef]
- Carnahan, A.; Joseph, S.W. Order XII. Aeromonadales ord. nov. In Bergey’s Manual of Systematic Bacteriology; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Williams & Wilkins: Philadelphia, PA, USA, 2005; pp. 556–578. [Google Scholar]
- Xue, M.; Xiao, Z.; Li, Y.; Jiang, N.; Liu, W.; Meng, Y.; Fan, Y.; Zeng, L.; Zhou, Y. Isolation, identification and characteristics of Aeromonas caviae from diseased largemouth bass (Micropterus salmoides). Fishes 2022, 7, 119. [Google Scholar] [CrossRef]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. Species distribution and prevalence of putative virulence factors in mesophilic Aeromonas spp. isolated from fresh retail sushi. Front. Microbiol. 2017, 8, 931. [Google Scholar] [CrossRef] [Green Version]
- Figueras, M.J.; Beaz-Hidalgo, R. Aeromonas: Introduction. In Encyclopedia of Food Microbiology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 24–30. [Google Scholar]
- Galbis, D.; Farfán, M.; Fusté, M.C.; Lorén, J.G. Aeromonas bivalvium sp. nov., isolated from bivalve molluscs. Int. J. Syst. Evol. Microbiol. 2007, 57, 582–587. [Google Scholar] [CrossRef]
- Carnahan, A.; Fanning, G.R.; Joseph, S.W. Aeromonas jandaei (formerly genospecies DNA group 9 A. sobria), a new sucrose-negative species isolated from clinical specimens. J. Clin. Microbiol. 1991, 29, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Alperi, A.; Martínez-Murcia, A.J.; Ko, W.C.; Monera, A.; Saavedra, M.J.; Figueras, M.J. Aeromonas taiwanensis sp. nov. and Aeromonas sanarellii sp. nov., clinical species from Taiwan. Int. J. Syst. Evol. Microbiol. 2010, 60, 2048–2055. [Google Scholar] [CrossRef] [Green Version]
- Demarta, A.; Küpfer, M.; Riegel, P.; Harf-Monteil, C.; Tonolla, M.; Peduzzi, R.; Monera, A.; José Saavedra, M.; Martínez-Murcia, A. Aeromonas tecta sp. nov., isolated from clinical and environmental sources. Syst. Appl. Microbiol. 2008, 31, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Balcázar, J.L. Aeromonas rivipollensis sp. nov., a novel species isolated from aquatic samples. J. Basic Microbiol. 2015, 55, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Ko, W.C.; Lee, N.Y.; Su, S.L.; Li, C.W.; Li, M.C.; Chen, Y.W.; Su, Y.C.; Shu, C.Y.; Lin, Y.T.; et al. Aeromonas isolates from fish and patients in Tainan City, Taiwan: Genotypic and phenotypic characteristics. Appl. Environ. Microbiol. 2019, 85, e01360-19. [Google Scholar] [CrossRef] [PubMed]
- Figueira, V.; Vaz-Moreira, I.; Silva, M.; Manaia, C.M. Diversity and antibiotic resistance of Aeromonas spp. in drinking and waste water treatment plants. Water Res. 2011, 45, 5599–5611. [Google Scholar] [CrossRef]
- Senderovich, Y.; Ken-Dror, S.; Vainblat, I.; Blau, D.; Izhaki, I.; Halpern, M. A molecular study on the prevalence and virulence potential of Aeromonas spp. recovered from patients suffering from diarrhea in Israel. PLoS ONE 2012, 7, e30070. [Google Scholar] [CrossRef]
- Larsen, A.; Tao, Z.; Bullard, S.A.; Arias, C.R. Diversity of the skin microbiota of fishes: Evidence for host species specificity. FEMS Microbiol. Ecol. 2013, 85, 483–494. [Google Scholar] [CrossRef]
- Chiarello, M.; Villéger, S.; Bouvier, C.; Bettarel, Y.; Bouvier, T. High diversity of skin-associated bacterial communities of marine fishes is promoted by their high variability among body parts, individuals and species. FEMS Microbiol. Ecol. 2015, 91, fiv061. [Google Scholar] [CrossRef] [Green Version]
- Sylvain, F.-É.; Holland, A.; Bouslama, S.; Audet-Gilbert, É.; Lavoie, C.; Luis Val, A.; Derome, N. Fish skin and gut microbiomes show contrasting signatures of host species and habitat. Appl. Environ. Microbiol. 2020, 86, 16. [Google Scholar] [CrossRef]
- Kawamoto, Y.; Kato, H.; Nagata, Y.; Urabe, J. Microbial communities developing within bulk sediments under fish carcasses on a tidal flat. PLoS ONE 2021, 16, e0247220. [Google Scholar] [CrossRef]
- Liu, Q.; Lai, Z.; Gao, Y.; Wang, C.; Zeng, Y.; Liu, E.; Mai, Y.; Yang, W.; Li, H. Connection between the gut microbiota of largemouth bass (Micropterus salmoides) and microbiota of the pond culture environment. Microorganisms 2021, 9, 1770. [Google Scholar] [CrossRef]
- Callewaert, C.; RavardHelffer, K.; Lebaron, P. Skin microbiome and its interplay with the environment. Am. J. Clin. Dermatol. 2020, 21 (Suppl. S1), 4–11. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Lee, N.M.; Pintado, J.; Planas, M. Phylogenetic characterization and in situ detection of bacterial communities associated with seahorses (Hippocampus guttulatus) in captivity. Syst. Appl. Microbiol. 2010, 33, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Tanu; Deobagkar, D.; Khandeparker, R.; Sreepada, R.A.; Sanaye, S.V.; Pawar, H.B. A study on bacteria associated with the intestinal tract of farmed yellow seahorse, Hippocampus kuda (Bleeker, 1852): Characterization and extracellular enzymes. Aquacult. Res. 2011, 43, 386–394. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Qin, G.; Luo, W.; Lin, Q. A novel pathogenic bacteria (Vibrio fortis) causing enteritis in cultured seahorses, Hippocampus erectus Perry, 1810. J. Fish Dis. 2016, 39, 765–769. [Google Scholar] [CrossRef]
- Kang, G.; Choi, K.M.; Joo, M.S.; Woo, W.S.; Park, C. A case report of secondary infection by Vibrio splendidus associated with gas bubble disease in syngnathid fishes (Syngnathus schlegeli and Hippocampus haema). Fish Aquat. Sci. 2022, 25, 40–48. [Google Scholar] [CrossRef]
- Abu-Elala, N.; Abdelsalam, M.; Marouf, S.; Setta, A. Comparative analysis of virulence genes, antibiotic resistance and gyrB-based phylogeny of motile Aeromonas species isolates from Nile tilapia and domestic fowl. Lett. Appl. Microbiol. 2015, 61, 429–436. [Google Scholar] [CrossRef]
- Fernández-Bravo, A.; Figueras, M.J. An update on the genus Aeromonas: Taxonomy, epidemiology, and pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Aeromonas. Global Water Pathogen Project. Available online: https://www.waterpathogens.org/book/aeromonas (accessed on 28 March 2023).
- Pessoa, R.B.; de Oliveira, W.F.; Marques, D.S.C.; dos Santos Correia, M.T.; de Carvalho, E.V.M.M.; Coelho, L.C.B.B. Thegenus Aeromonas: A general approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef]
- Borella, L.; Salogni, C.; Vitale, N.; Scali, F.; Moretti, V.; Pasquali, P.; Alborali, G.L. Motile aeromonads from farmed and wild freshwater fish in northern Italy: An evaluation of antimicrobial activity and multidrug resistance during 2013 and 2016. Acta Vet. Scand. 2020, 62, 6. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Alperi, A.; Buján, N.; Romalde, J.L.; Figueras, M.J. Comparison of phenotypical and genetic identification of Aeromonas strains isolated from diseased fish. Syst. Appl. Microbiol. 2010, 33, 149–153. [Google Scholar] [CrossRef]
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas salmonicida: Updates on an old acquaintance. Dis. Aquat. Organ. 2016, 120, 49–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Wang, N.; Pan, Z.H.; Lu, C.P.; Liu, Y.J. Identity and virulence properties of Aeromonas isolates from diseased fish, healthy controls and water environment in China. Lett. Appl. Microbiol. 2012, 55, 224–233. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega, R.C.M.H.; Tabugo, S.R.M.; Martinez, J.G.T.; Padasas, C.S.; Balcázar, J.L. Occurrence of Aeromonas Species in the Cutaneous Mucus of Barbour’s Seahorses (Hippocampus barbouri) as Revealed by High-Throughput Sequencing. Animals 2023, 13, 1241. https://doi.org/10.3390/ani13071241
Ortega RCMH, Tabugo SRM, Martinez JGT, Padasas CS, Balcázar JL. Occurrence of Aeromonas Species in the Cutaneous Mucus of Barbour’s Seahorses (Hippocampus barbouri) as Revealed by High-Throughput Sequencing. Animals. 2023; 13(7):1241. https://doi.org/10.3390/ani13071241
Chicago/Turabian StyleOrtega, Rose Chinly Mae H., Sharon Rose M. Tabugo, Joey Genevieve T. Martinez, Chinee S. Padasas, and José L. Balcázar. 2023. "Occurrence of Aeromonas Species in the Cutaneous Mucus of Barbour’s Seahorses (Hippocampus barbouri) as Revealed by High-Throughput Sequencing" Animals 13, no. 7: 1241. https://doi.org/10.3390/ani13071241