
Effect of Diet on Growth Performance of First Crab Stage Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): A Comparison of Three Different Regimens


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection
2.2. Rearing of Larval and First Crabs
2.3. Growth Experiment
2.3.1. Raw Material
2.3.2. Feeding Regimen
2.4. Statistical Analysis
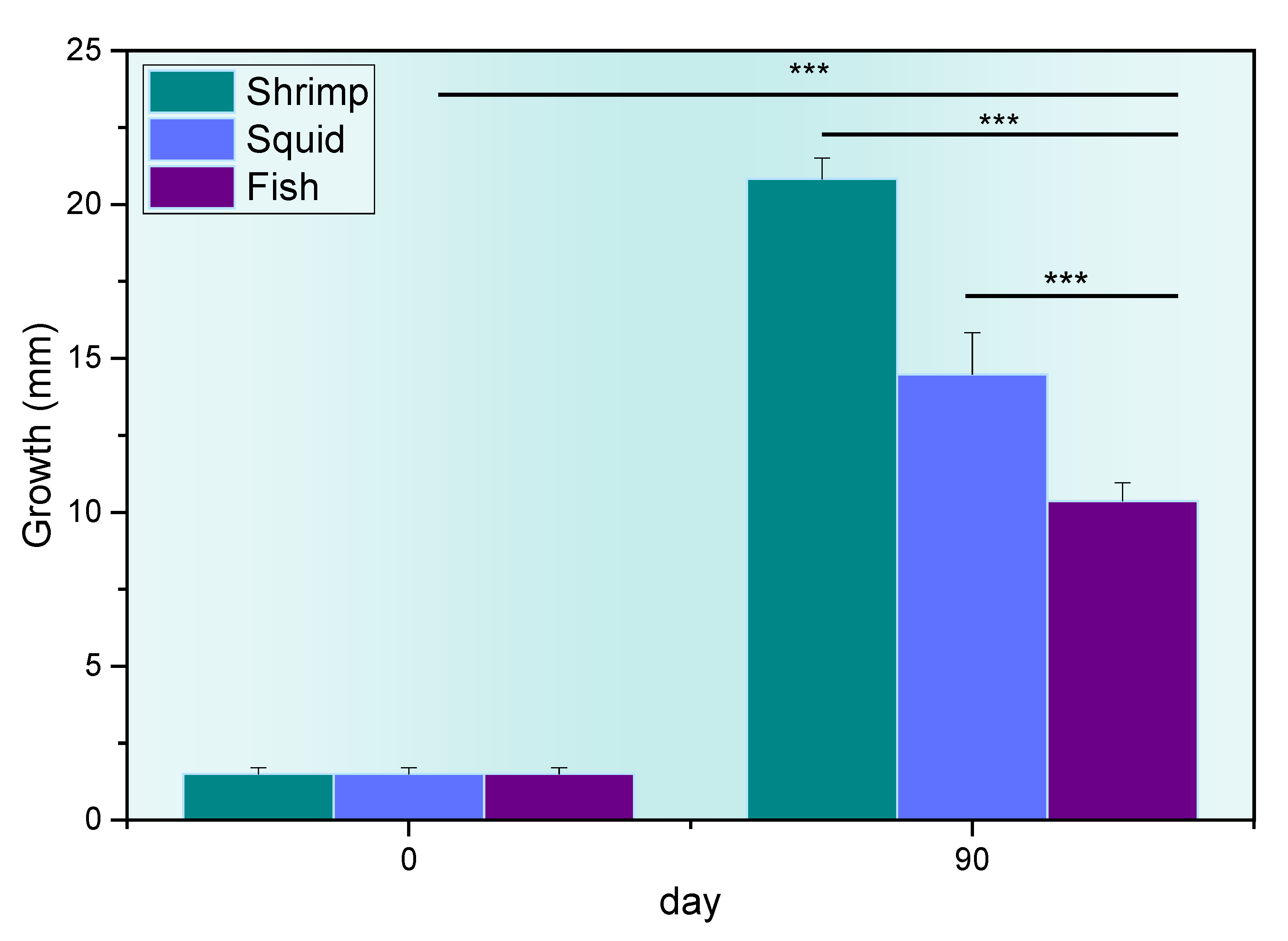
3. Results and Discussions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mancinelli, G.; Bardelli, R.; Zenetos, A. A global occurrence database of the Atlantic blue crab Callinectes sapidus. Sci. Data 2021, 81, 111. [Google Scholar] [CrossRef] [PubMed]
- Overton, A.S.; Margraf, F.J.; May, E.B. Spatial and Temporal Patterns in the Diet of Striped Bass in Chesapeake Bay. Trans. Am. Fish. Soc. 2011, 138, 915–926. [Google Scholar] [CrossRef]
- Weatherall, T.F.; Scheef, L.P.; Buskey, E.J. Spatial and temporal settlement patterns of blue crab (Callinectes sapidus and Callinectes similis) megalopae in a drought-prone Texas estuary. Estuar. Coast. Shelf Sci. 2018, 214, 89–97. [Google Scholar] [CrossRef]
- Moir, F.; Weissburg, M.J. Cautious cannibals: Behavioral responses of juvenile and adult blue crabs to the odor of injured conspecifics. J. Exp. Mar. Biol. Ecol. 2009, 369, 87–92. [Google Scholar] [CrossRef]
- Moksnes, P.O.; Lipcius, R.N.; Pihl, L.; Van Montfrans, J. Cannibal–prey dynamics in young juveniles and postlarvae of the blue crab. J. Exp. Mar. Biol. Ecol. 1997, 215, 157–187. [Google Scholar] [CrossRef]
- Lipcius, R.N.; Seitz, R.D.; Seebo, M.S.; Colón-Carrión, D. Density, abundance and survival of the blue crab in seagrass and unstructured salt marsh nurseries of Chesapeake Bay. J. Exp. Mar. Biol. Ecol. 2005, 319, 69–80. [Google Scholar] [CrossRef]
- Bromilow, M. 2022 Chesapeake Bay Blue Crab Advisory Report CBSAC Meeting July 6; Chesapeake Bay Stock Assessment Committee: Annapolis, MD, USA, 2022; p. 5. [Google Scholar]
- Lipcius, R.N.; Stockhausen, W.T. Concurrent decline of the spawning stock, recruitment, larval abundance, and size of the blue crab Callinectes sapidus in Chesapeake Bay. Mar. Ecol. Prog. Ser. 2002, 226, 45–61. [Google Scholar] [CrossRef]
- Etherington, L.L.; Eggleston, D.B. Large-scale blue crab recruitment: Linking postlarval transport, post-settlement planktonic dispersal, and multiple nursery habitats. Mar. Ecol. Prog. Ser. 2000, 204, 179–198. [Google Scholar] [CrossRef] [Green Version]
- Etherington, L.L.; Eggleston, D.B.; Stockhausen, W.T. Partitioning loss rates of early juvenile blue crabs from seagrass habitats into mortality and emigration. Bull. Mar. Sci. 2003, 72, 371–391. [Google Scholar]
- Secor, D.H.; Mines, A.H.; Place, A.R. Japanese Hatchery-Based Stock Enhancement: Lessons for the Chesapeake Bay Blue Crab. 2002. Available online: https://repository.si.edu/bitstream/handle/10088/18569/serc_Secor_Hines_2004_MDSG_1_44.pdf (accessed on 8 November 2022).
- Munro, J.L.; Bell, J.D. Enhancement of marine fisheries resources. Rev. Fish. Sci. 2008, 5, 185–222. [Google Scholar] [CrossRef]
- Davis, J.L.D.; Young-Williams, A.C.; Hines, A.H.; Zohar, Y. Assessing the potential for stock enhancement in the case of the Chesapeake Bay blue crab (Callinectes sapidus). Can. J. Fish. Aquat. Sci. 2011, 62, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.L.; Young-Williams, A.C.; Aguilar, R.; Carswell, B.L.; Goodison, M.R.; Hines, A.H.; Kramer, M.A.; Zohar, Y.; Zmorer, O. Differences between Hatchery-Raised and Wild Blue Crabs: Implications for Stock Enhancement Potential. Trans. Am. Fish. Soc. 2011, 133, 1–14. [Google Scholar] [CrossRef]
- Zmora, O.; Findiesen, A.; Stubblefield, J.; Frenkel, V.; Zohar, Y. Large-scale juvenile production of the blue crab Callinectes sapidus. Aquaculture 2005, 244, 129–139. [Google Scholar] [CrossRef]
- Quinitio, E.T.; Estepa, F.D.P. Survival and growth of Mud crab, Scylla serrata, juveniles subjected to removal or trimming of chelipeds. Aquaculture 2011, 318, 229–234. [Google Scholar] [CrossRef]
- Josileen, J.; Menon, N.G. Larval Stages of the Blue Swimmer Crab, Portunus pelagicus (Linnaeus, 1758) (Decapoda, Brachyura). Crustaceana 2004, 77, 785–803. Available online: http://www.jstor.org/stable/20105760 (accessed on 1 November 2022).
- Costlow, J.D., Jr.; Bookhout, C.G. The Larval Development of Callınectes sapıdus Rathbun Reared ın the Laboratory. Biol. Bull. 1959, 116, 373–396. [Google Scholar] [CrossRef]
- Hasek, B.E.; Rabalais, N.N. A comparison of molt states of blue crab megalopae, Callinectes sapidus (Rathbun), sampled with artificial collectors and plankton nets. J. Exp. Mar. Biol. Ecol. 2001, 265, 15–27. [Google Scholar] [CrossRef]
- Santos, J.; Borges, T. Trophic relationships in deep-water fish communities off Algarve, Portugal. Fish. Res. 2001, 51, 337–341. [Google Scholar] [CrossRef]
- Bryceson, I.; Massinga, A. Coastal resources and management systems influenced by conflict and migration: Mecufi, Mozambique. AMBIO J. Hum. Environ. 2002, 31, 512–517. [Google Scholar] [CrossRef]
- Ebersole, E.L.; Kennedy, V.S. Prey preferences of blue crabs Callinectes sapidus Feeding on Three Bivalve Species. Oceanogr. Lit. Rev. 1995, 10, 878. Available online: https://www.infona.pl//resource/bwmeta1.element.elsevier-097804e9-5c8d-397b-976a-97a6578b6de9 (accessed on 31 January 2023). [CrossRef] [Green Version]
- Møller, H.; Lee, S.Y.; Paterson, B.; Mann, D. Cannibalism contributes significantly to the diet of cultured sand crabs, Portunus pelagicus (L.): A dual stable isotope study. J. Exp. Mar. Biol. Ecol. 2008, 361, 75–82. [Google Scholar] [CrossRef]
- Lovrich, G.A.; Sainte-Marie, B. Cannibalism in the snow crab, Chionoecetes opilio (O. Fabricius) (Brachyura: Majidae), and its potential importance to recruitment. J. Exp. Mar. Biol. Ecol. 1997, 211, 225–245. [Google Scholar] [CrossRef]
- Wahle, R.A. Revealing stock-recruitment relationships in lobsters and crabs: Is experimental ecology the key? Fish. Res. 2003, 65, 3–32. [Google Scholar] [CrossRef]
- Sulkin, S.D. Nutritional requirements during larval development of the portunid crab, Callinectes sapidus Rathbun. J. Exp. Mar. Biol. Ecol. 1978, 34, 29–41. [Google Scholar] [CrossRef]
- Wang, T.; He, K.; Blaney, L.; Chung, J.S. 17β-Estradiol (E2) may be involved in the mode of crustacean female sex hormone (CFSH) action in the blue crab, Callinectes sapidus. Front. Endocrinol. 2022, 13, 1632. [Google Scholar] [CrossRef]
- Bembe, S.; Liang, D.; Chung, J.S. Optimal temperature and photoperiod for the spawning of blue crab, Callinectes sapidus, in captivity. Aquac. Res. 2017, 48, 5498–5505. [Google Scholar] [CrossRef]
- Maurer, L.; Liang, D.; Chung, J.S. Effects of prey densities and dietary supplementation on the larval development of the blue crab Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae). J. Crustac. Biol. 2017, 37, 674–682. [Google Scholar] [CrossRef]
- Ñañez Pacheco, G.K.; Sanabio Maldonado, N.S.; Pastrana Alta, R.Y.; Aguilar Vitorino, H. Short exposure of Artemia salina to group-12 metals: Comparing hatchability, mortality, lipid peroxidation, and swimming speed. Ecotoxicol. Environ. Saf. 2021, 213, 112052. [Google Scholar] [CrossRef] [PubMed]
- Hernández Moresino, R.D.; Helbling, E.W. Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds. Mar. Drugs 2010, 8, 1681. [Google Scholar] [CrossRef] [Green Version]
- Sarower, M.G.; Mahmud-Al-Hasan, M.; Rahman, M.S.; Hasan, M.M.; Ahmmed, M.K.; Ali, M.Y.; Giteru, S.G.; Banu, G.R. Comparative growth and morphometric assessment between cultures of wild and hatchery-produced mud crabs. Heliyon 2021, 7, e07964. [Google Scholar] [CrossRef]
- Vitorino, H.A.; Alta, R.Y.P.; Ortega, P. Lipid peroxidation in hepatopancreas, gill, and hemolymph of male and female crabs Platyxanthus orbignyi after air exposure. J. Mar. Sci. Eng. 2019, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Baylon, A.N.; Failaman, J.C. Larval rearing of the mud crab Scylla serrata in the Philippines. In Mud Crab Aquaculture and Biology: Proceedings of an International Scientific Forum Held in Darwin, Australia, 21–24 April 1997; Australian Centre for International Agricultural Research: Canberra, Australia, 1999; pp. 141–146. Available online: https://ageconsearch.umn.edu/record/135187/files/PR078.pdf#page=143 (accessed on 1 November 2022).
- Daly, B.; Swingle, J.S.; Eckert, G.L. Increasing hatchery production of juvenile red king crabs (Paralithodes camtschaticus) through size grading. Aquaculture 2012, 364–365, 206–211. [Google Scholar] [CrossRef]
- Baylon, J.C. Appropriate food type, feeding schedule and Artemia density for the zoea larvae of the mud crab, Scylla tranquebarica (Crustacea:Decapoda:Portunidae). Aquaculture 2009, 288, 190–195. [Google Scholar] [CrossRef]
- USDA. FoodData Central. U.S. Department of Agriculture. 2022. Available online: https://fdc.nal.usda.gov/fdc-app.html#/ (accessed on 18 November 2022).
- Islam, T.; Saha, D.; Bhowmik, S.; Nordin, N.; Islam, S.; Nur, A.A.U.; Begum, M. Nutritional properties of wild and fattening mud crab (Scylla serrata) in the south-eastern district of Bangladesh. Heliyon 2022, 8, e09696. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, X.; Wang, X.; Bu, X.; Li, E.; Zhang, M.; Qin, C.; Qin, J.G.; Chen, L. Molting, tissue calcium-phosphorus deposition and immunity of juvenile Chinese mitten crab (Eriocheir sinensis) fed different levels of calcium and vitamin D3. Aquaculture 2022, 554, 738124. [Google Scholar] [CrossRef]
- Zanotto, F.P.; Pinheiro, F.; Sá, M.G. The importance of dietary calcium consumption in two species of semi-terrestrial grapsoid crabs. Iheringia Sér. Zool. 2009, 99, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Belgrad, B.A.; Griffen, B.D. The Influence of Diet Composition on Fitness of the Blue Crab, Callinectes sapidus. PLoS ONE 2016, 11, e0145481. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| n = 270 Animals | Group 1 (n = 90) | Group 2 (n = 90) | Group 3 (n = 90) |
|---|---|---|---|
| Diet (0.001 g/mL) | Shrimp | Squid | Fish |
| Size (day = 0), mm | 1.50 ± 0.20 | ||
| Size (day = 90), mm | |||
| Plate 1 * | 21.0 ± 1.50 | 11.9 ± 1.20 | 11.1 ± 0.40 |
| Plate 2 | 20.4 ± 1.00 | 16.1 ± 1.10 | 10.5 ± 1.40 |
| Plate 3 | 21.2 ± 1.60 | 12.4 ± 1.10 | 9.90 ± 1.60 |
| Plate 4 | 21.2 ± 0.90 | 14.5 ± 0.40 | 11.1 ± 0.40 |
| Plate 5 | 22.1 ± 1.20 | 15.5 ± 2.20 | 10.6 ± 0.90 |
| Plate 6 | 20.0 ± 2.10 | 15.3 ± 0.70 | 9.50 ± 1.40 |
| Plate 7 | 20.1 ± 1.00 | 14.4 ± 1.30 | 11.0 ± 2.20 |
| Plate 8 | 21.4 ± 1.30 | 15.2 ± 2.10 | 9.70 ± 1.20 |
| Plate 9 | 20.1 ± 1.40 | 15.1 ± 1.80 | 9.90 ± 1.30 |
| Water Quality/Physicochemical Variables | Optimal Level |
|---|---|
| Ammonia (NH3, NH4+) and nitrite (NO2−) | <1 ppm |
| Dissolved oxygen (DO) | 90% saturation (approx. 6.7 ppm) |
| pH | 7.7–8.0 |
| Salinity | 30 ppt |
| Temperature | 22 ± 1 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gencer, Ö.; Aguilar Vitorino, H. Effect of Diet on Growth Performance of First Crab Stage Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): A Comparison of Three Different Regimens. Animals 2023, 13, 1242. https://doi.org/10.3390/ani13071242
Gencer Ö, Aguilar Vitorino H. Effect of Diet on Growth Performance of First Crab Stage Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): A Comparison of Three Different Regimens. Animals. 2023; 13(7):1242. https://doi.org/10.3390/ani13071242
Chicago/Turabian StyleGencer, Övgü, and Hector Aguilar Vitorino. 2023. "Effect of Diet on Growth Performance of First Crab Stage Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): A Comparison of Three Different Regimens" Animals 13, no. 7: 1242. https://doi.org/10.3390/ani13071242