
Review: Recent Applications of Gene Editing in Fish Species and Aquatic Medicine


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Fish Industry
1.2. Gene Editing
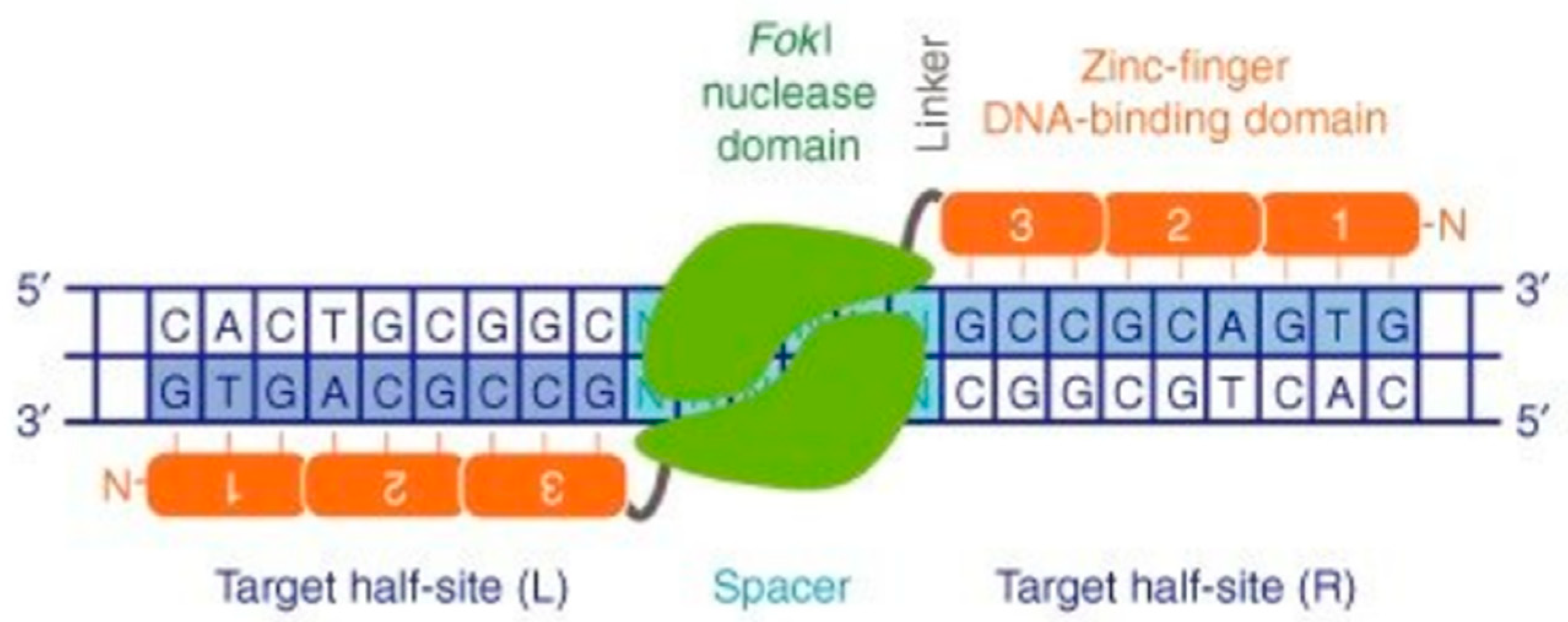
1.2.1. Zinc Finger Nucleases (ZFNs)
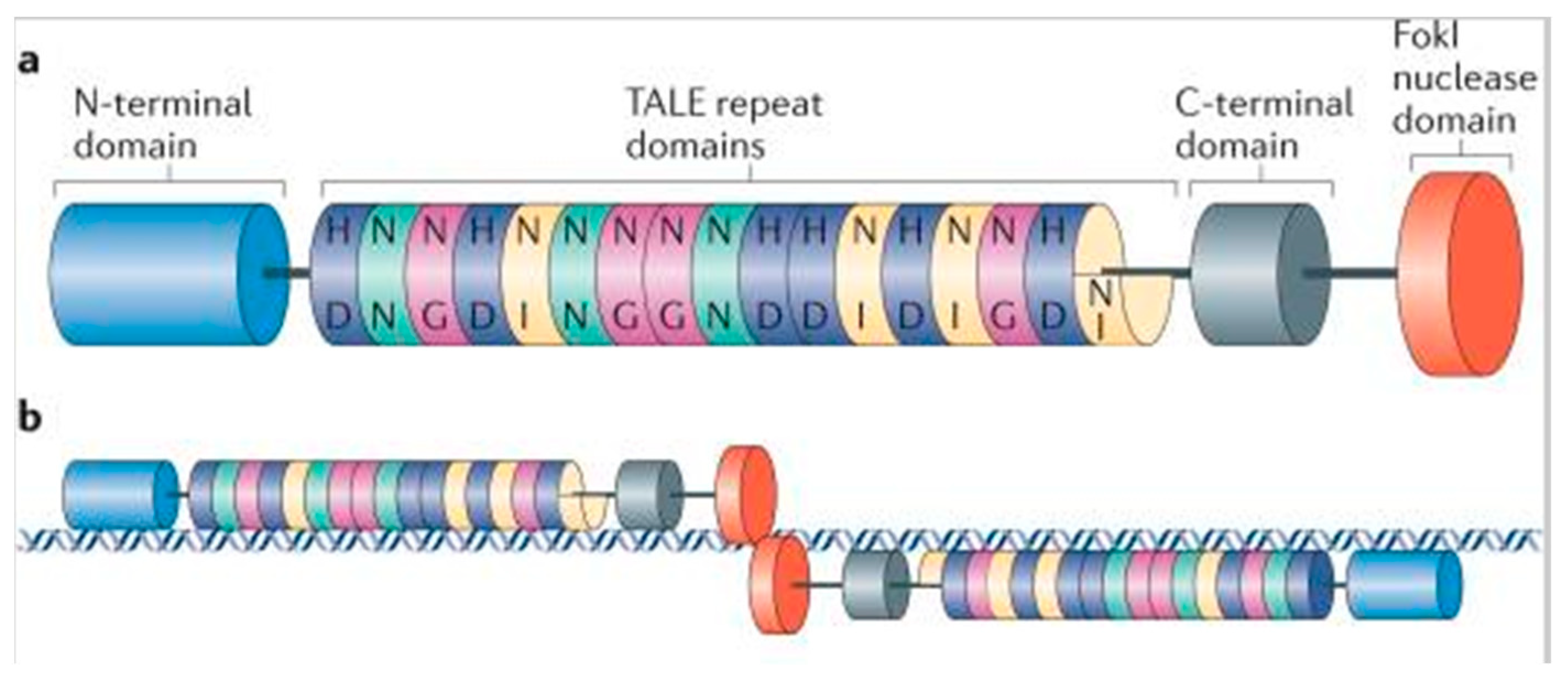
1.2.2. Transcriptional Activator-Like Effector Nucleases (TALENs)
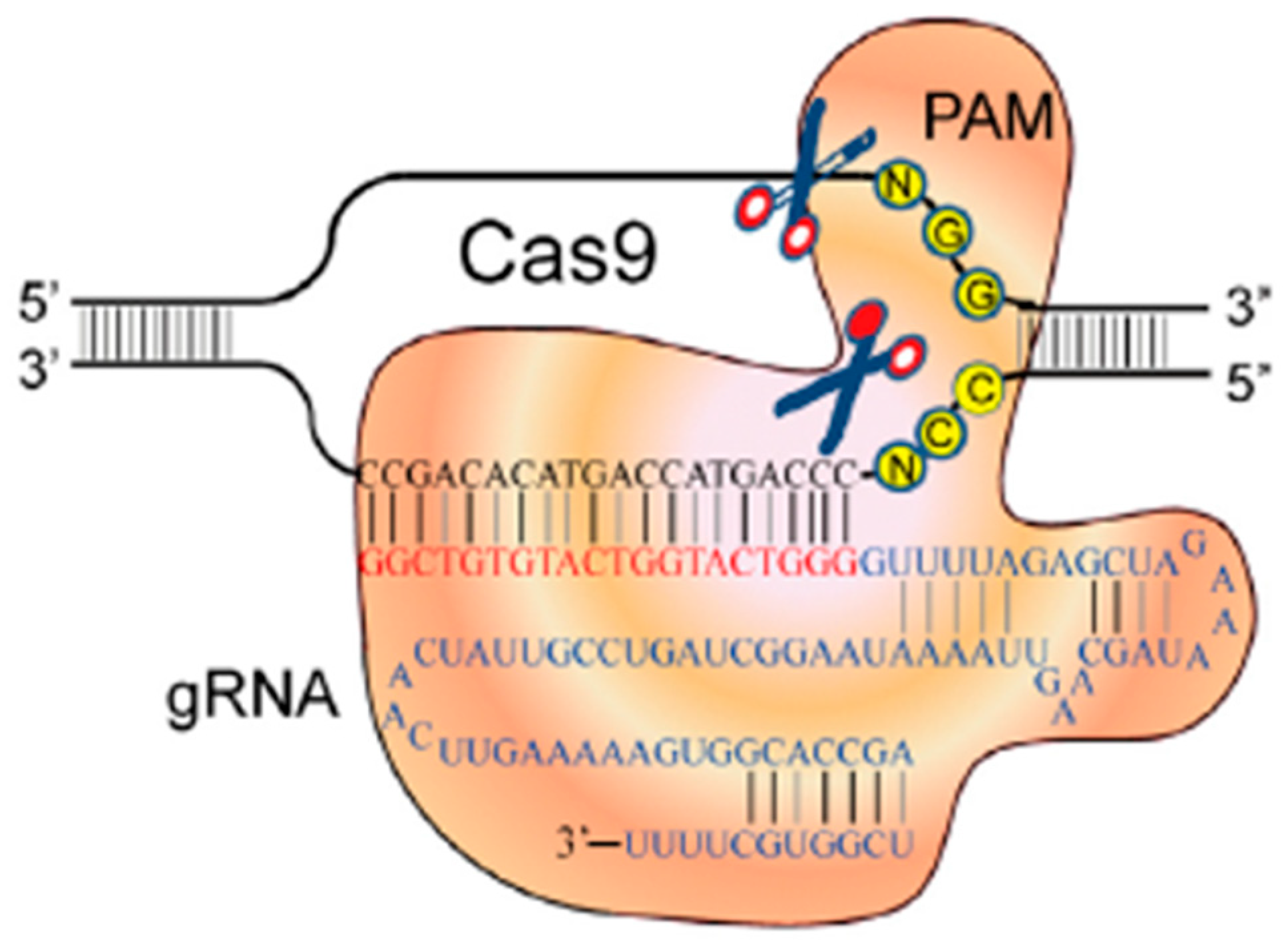
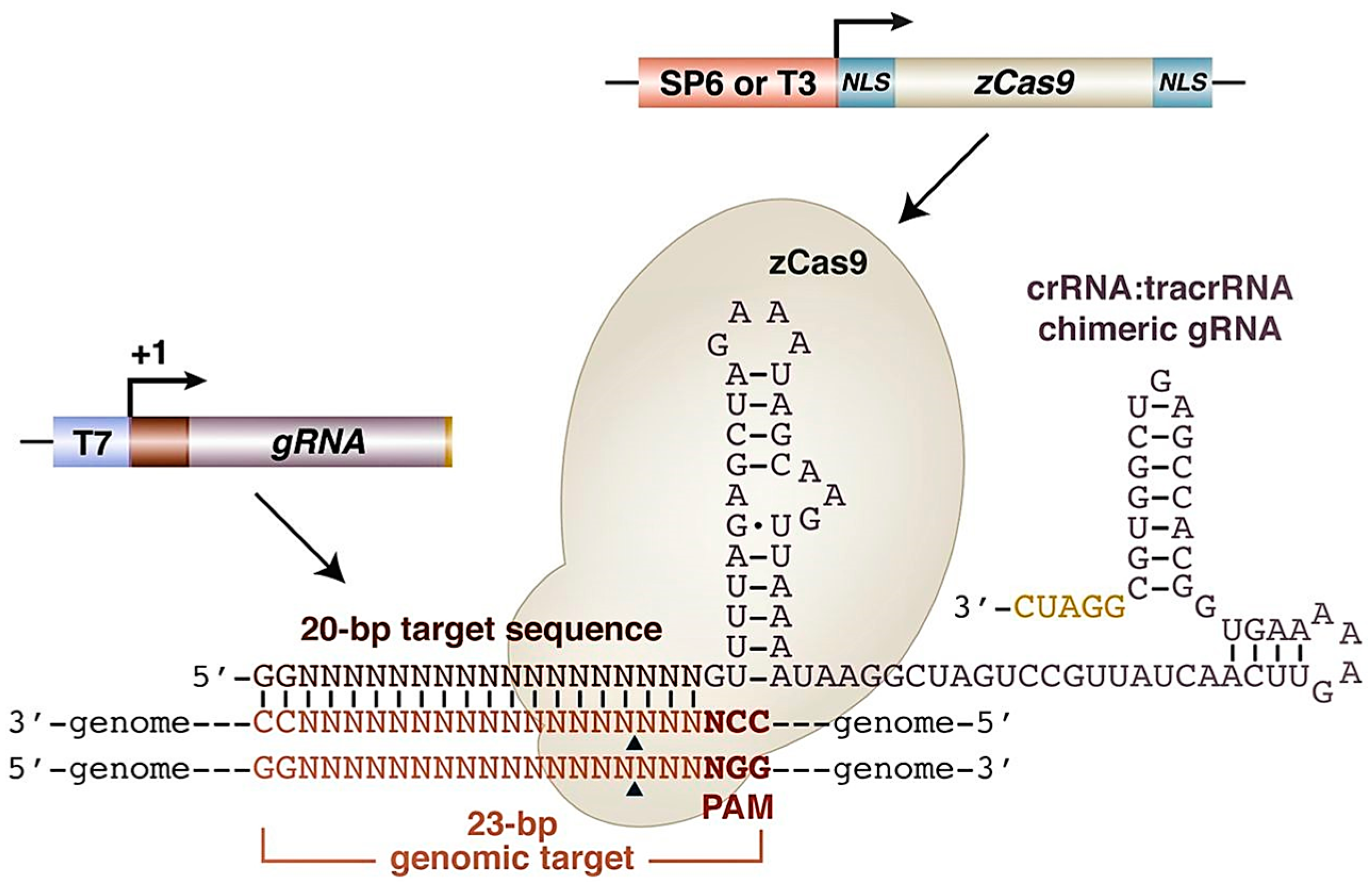
1.2.3. Clustered Regularly Interspaced Short Palindromic Repeats—Cas 9 System (CRISPR/Cas9)
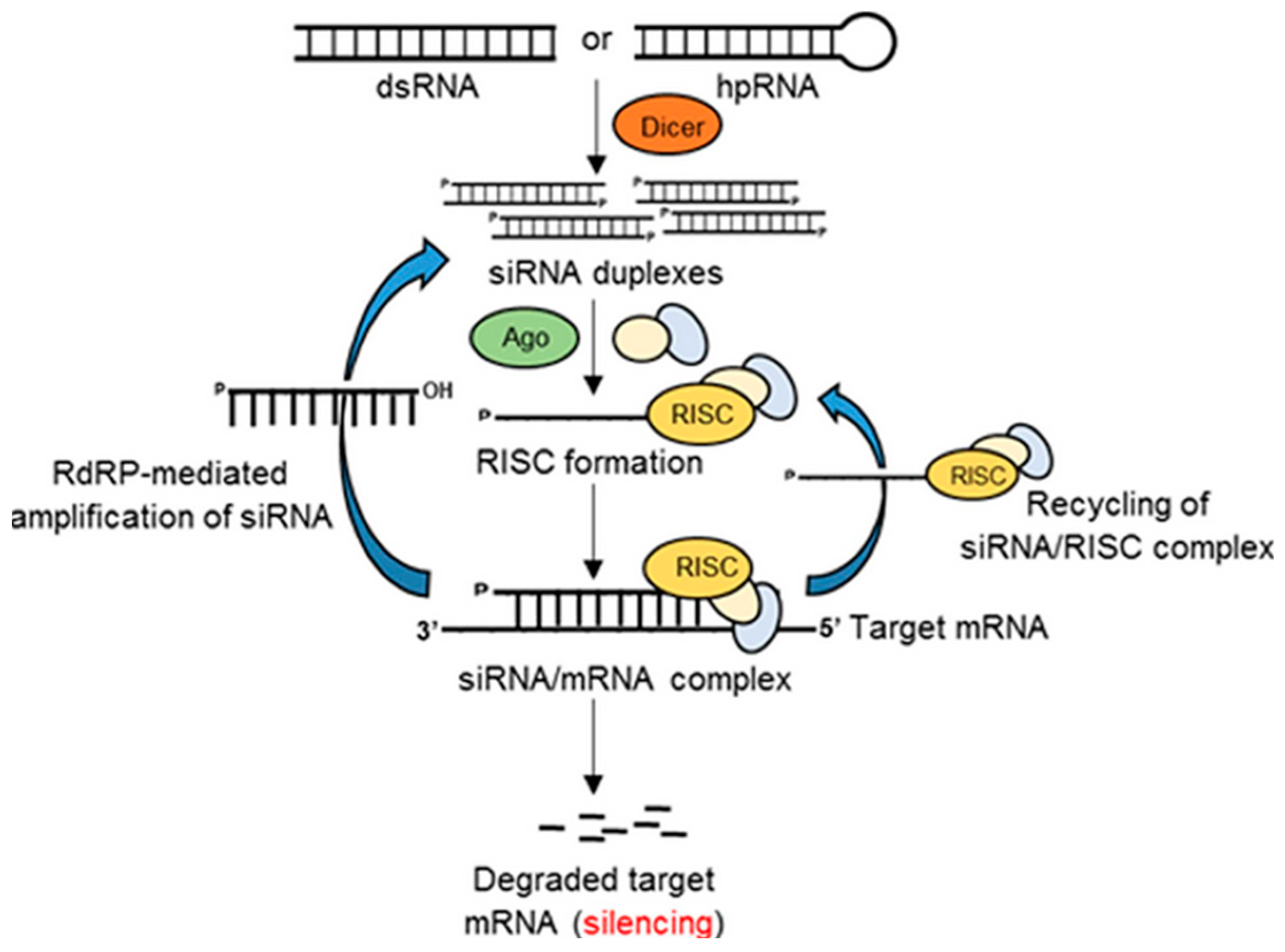
1.2.4. Gene Silencing
1.2.5. Applications (Targeted Gene Modification in Aquatic Animals)
2. Gene Editing in Fish Farm Species Using CRISPR/Cas9 and other Gene Editing Tools
2.1. Gene Editing in Fishery Science
2.2. Gene Editing in Mono-Sex Population
2.3. Gene Editing in Sterility of Fish
2.4. Gene Editing in Reproduction
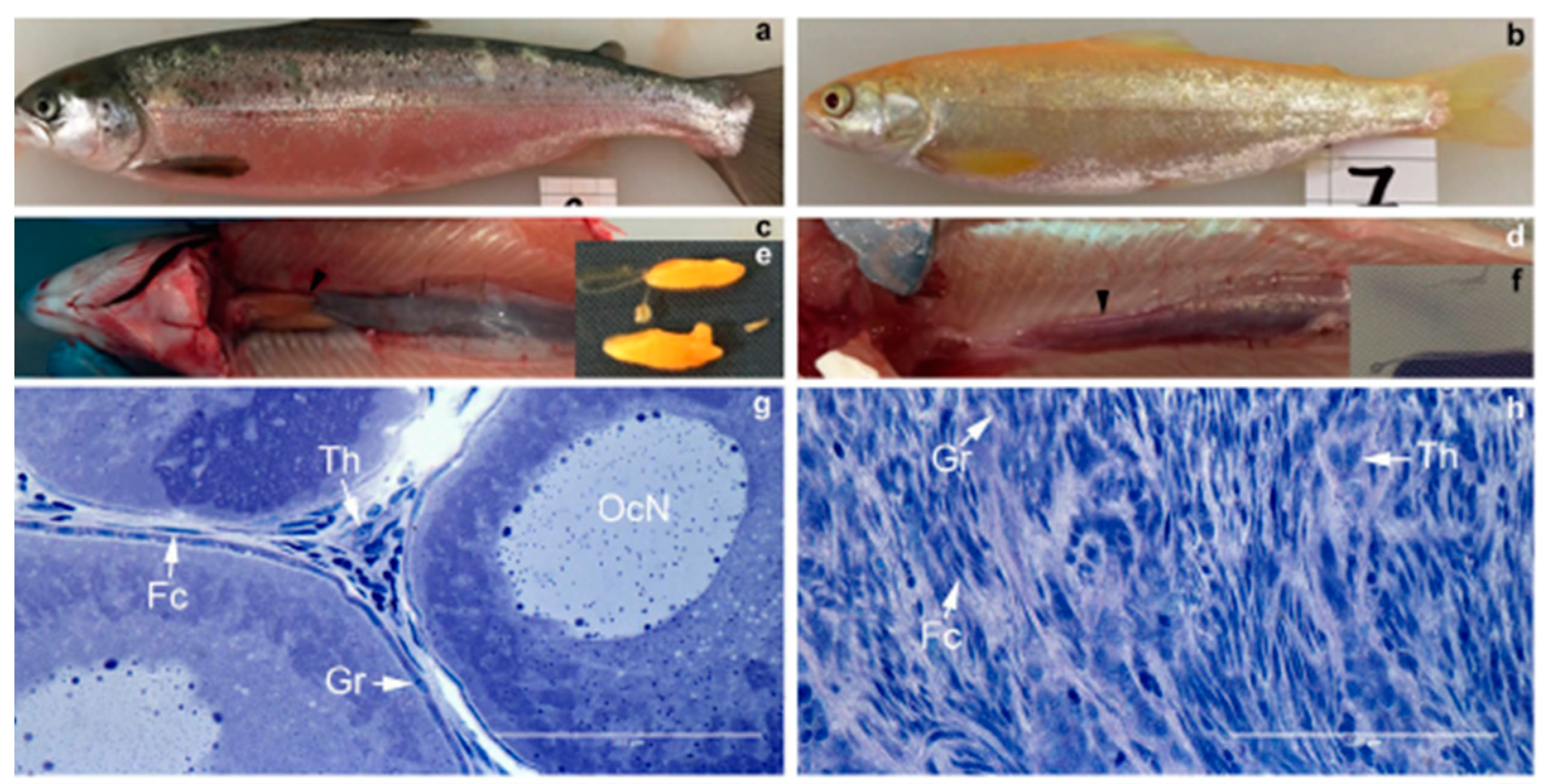
2.5. Gene Editing in Fast-Growing Fishes
2.6. Gene Editing in Ornamental Fishes
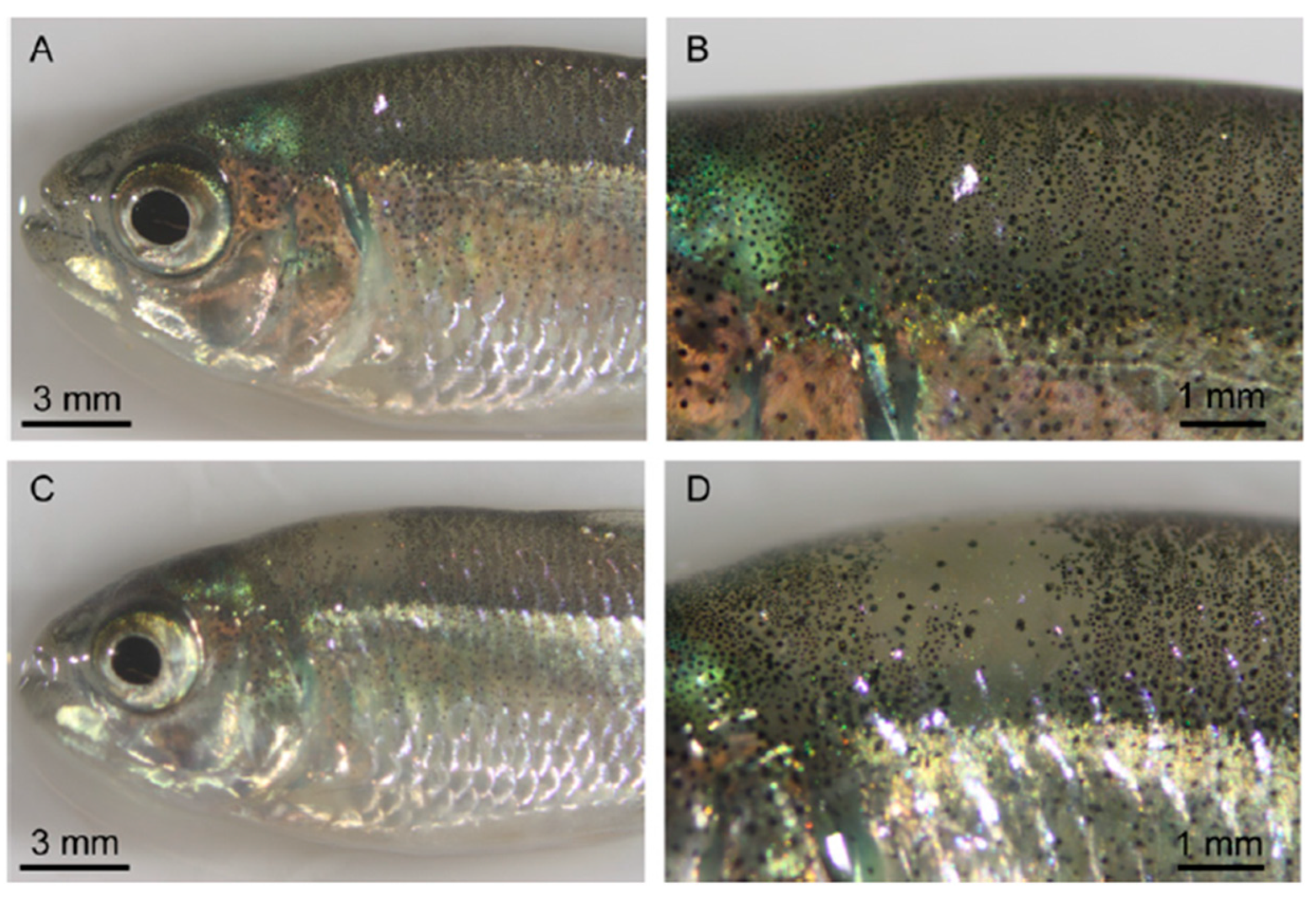
2.7. Gene Editing in Pigmentation
2.8. Gene Editing in Growth
2.9. Gene Editing in Body Configuration
2.10. Gene Editing in Oomycetes
3. Gene Silencing in Fish Medicine
3.1. Gene Silencing in Viral Disease of Fish Medicine
3.2. Gene Silencing in Parasitic Disease of Fish Medicine
3.3. Gene Silencing for Gene Function Studies in Fish Medicine
3.4. Gene Silencing in Oomycetes
3.5. Gene Silencing in Crustaceans
3.5.1. Gene Silencing in Viral Diseases of Crustaceans
3.5.2. Gene Silencing in Bacterial Disease of Crustaceans
3.5.3. Gene Silencing in Decreased Glucose Levels by Crustaceans
3.5.4. Gene Silencing in Pleiotropic Effect by Crustaceans
3.5.5. Gene Silencing in Reproduction of Crustaceans
4. Ethical Issues and Future of Gene Editing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ZFN | Zinc finger nuclease |
| TALEN | Transcriptional activator-like effector nuclease |
| HDR | Homology-directed repair |
| NHEJ | Nonhomologous end joining |
| DSB | Double-strand break |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| sgRNA/gRNA | Single guide RNA |
| crRNA | CRISPR RNA |
| tracrRNA | Trans-activating crRNA |
| PAM | Protospacer adjacent motif |
| RNAi | RNA interference |
| dsRNA | Double-stranded RNA |
| siRNA | Small interfering RNA |
| hpRNA | Hairpin RNA |
| shRNA | Short-hairpin RNA |
| Ago | Argonaute protein |
| RISC | RNA-induced silencing complex |
| RdRp | RNA-dependent RNA polymerase |
| SVCV | Spring viraemia of carp virus |
| EPC | Epithelioma papulosum cyprinid |
| CyHV | 3-cyprinid herpesvirus-3 |
| TK | Thymidine kinase |
| DP | DNA polymerase |
| shRNA | Short hairpin RNA |
| MSTN | Myostatin |
| KO | Knockout |
| GM | Genetically modified |
| GH | Growth hormone |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| CCB | Common carp brain |
| ALF | Anti-lipopolysaccharide factor |
| WSSV | White spot syndrome virus |
| CHH | Crustacean hyperglycemic hormone |
| PO | Phenoloxidase |
| proPO | Prophenoloxidase |
| SEM | Scanning electron microscopy |
| YHV | Yellow head virus |
| TSV | Taura syndrome virus |
| GIH | Gonad-inhibiting hormone |
| Vg | Vitellogenin |
| CHH | Crustacean hyperglycemic hormone |
| MIH | Molt-inhibiting hormone |
| GIH | Gonad-inhibiting hormone |
| eGFP | Exogenous GFP |
References
- Edwards, P.; Zhang, W.; Belton, B.; Little, D.C. Misunderstandings, myths and mantras in aquaculture: Its contribution to world food supplies has been systematically over reported. Mar. Policy 2019, 106, 103547. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022; Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Food and Agriculture Organization (FAO). FAO: Fisheries and Aquaculture Statistics; FAO: Rome, Italy, 2014. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2012; FAO Fisheries Technical Paper; FAO: Rome, Italy, 2012. [Google Scholar]
- Boyd, C.E.; D’Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso, J.R., Jr.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquac. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- World Bank. Fish to 2030: Prospects for Fisheries and Aquaculture—World Bank Report Number 83177-GLB; Agriculture and Environmental Services Discussion Paper; World Bank: Washington, DC, USA, 2013. [Google Scholar]
- Perota, A.; Lagutina, I.; Quadalti, C.; Lazzari, G.; Cozzi, E.; Galli, C. The Applications of Genome Editing in Xenotransplantation. J. Genet. Genom. 2016, 43, 233–237. [Google Scholar] [CrossRef]
- Cathomen, T.; Joung, J.K. Zinc-finger Nucleases: The Next Generation Emerges. Mol. Ther. 2008, 16, 1200–1207. [Google Scholar] [CrossRef]
- Malik, Y.S.; Barh, D.; Azevedo, V.A.D.C.; Khurana, S.P. Genomics and Biotechnological Advances in Veterinary, Poultry, and Fisheries; Elsevier Inc.: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Gratacap, R.L.; Wargelius, A.; Edvardsen, R.B.; Houston, R.D. Potential of Genome Editing to Improve Aquaculture Breeding and Production. Trends Genet. 2019, 35, 672–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karre, A. Gene Editing Technology. 2020. Available online: https://www.researchgate.net/publication/347442835_GENE_EDITING_TECHNOLOGY (accessed on 22 March 2023).
- Bibikova, M.; Beumer, K.; Trautman, J.K.; Carroll, D. Enhancing Gene Targeting with Designed Zinc Finger Nucleases. Science 2003, 300, 764. [Google Scholar] [CrossRef] [Green Version]
- Moscou, M.J.; Bogdanove, A.J. A Simple Cipher Governs DNA Recognition by TAL Effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Egelie, K.J.; Graff, G.; Strand, S.P.; Johansen, B. The emerging patent landscape of CRISPR–Cas gene editing technology. Nat. Biotechnol. 2016, 34, 1025–1031. [Google Scholar] [CrossRef]
- Xu, X.; Hulshoff, M.S.; Tan, X.; Zeisberg, M.; Zeisberg, E.M. CRISPR/Cas Derivatives as Novel Gene Modulating Tools: Possibilities and In Vivo Applications. Int. J. Mol. Sci. 2020, 21, 3038. [Google Scholar] [CrossRef]
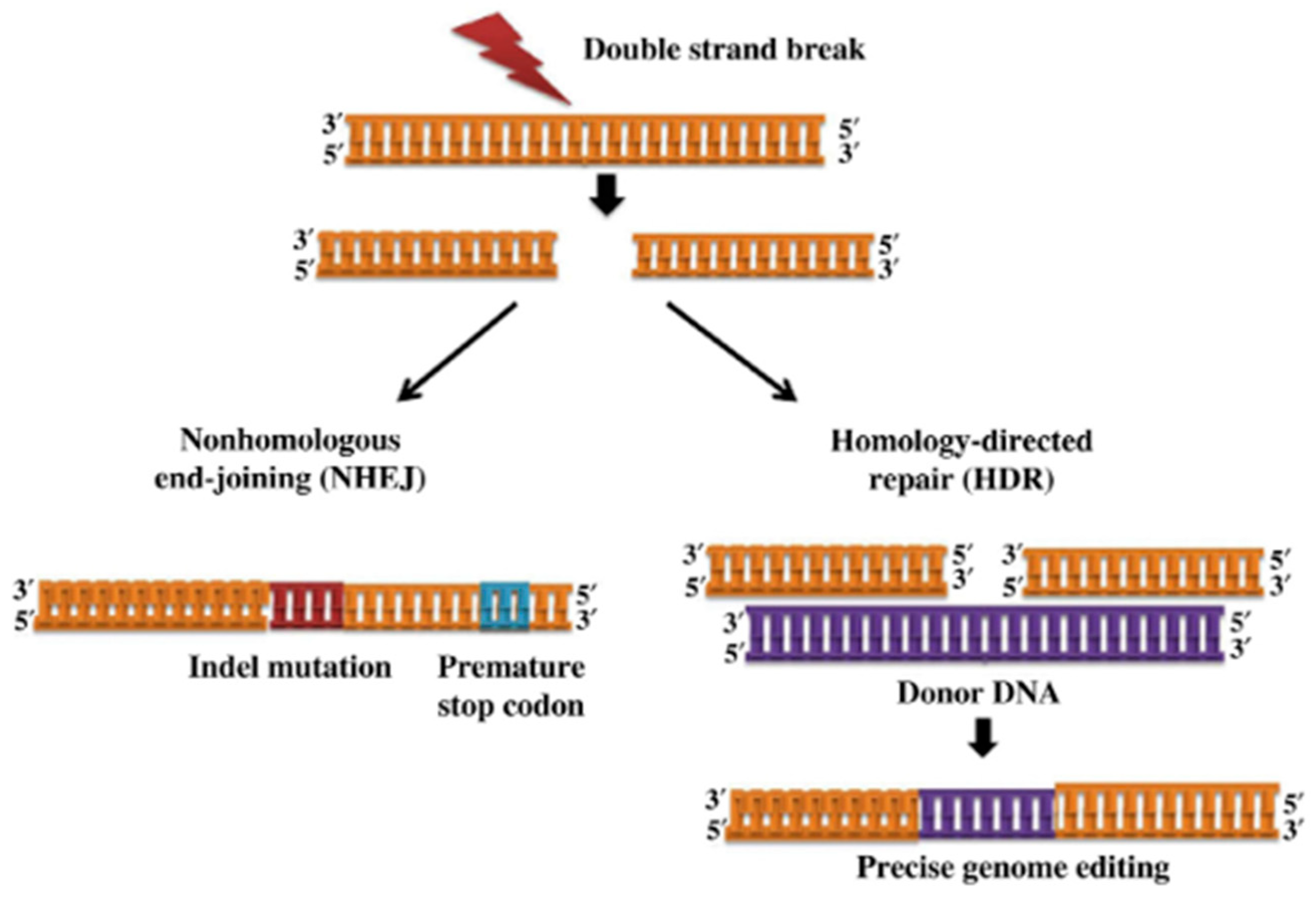
- Takata, M.; Sasaki, M.S.; Sonoda, E.; Morrison, C.; Hashimoto, M.; Utsumi, H.; Yamaguchi-Iwai, Y.; Shinohara, A.; Takeda, S. Homologous recombination and non-homologous end-joining pathways of DNA double-strand break repair have overlapping roles in the maintenance of chromosomal integrity in vertebrate cells. EMBO J. 1998, 17, 5497–5508. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.E. Non-homologous end joining as a mechanism of DNA repair. Curr. Biol. 2001, 11, R455–R457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, M.R. The Mechanism of Double-Strand DNA Break Repair by the Nonhomologous DNA End-Joining Pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lans, H.; Marteijn, J.A.; Vermeulen, W. ATP-dependent chromatin remodeling in the DNA-damage response. Epigenetics Chromatin 2012, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Swiech, L.; Heidenreich, M.; Banerjee, A.; Habib, N.; Li, Y.; Trombetta, J.J.; Sur, M.; Zhang, F. In vivo interrogation of gene function in the mammalian brain using CRISPR-Cas9. Nat. Biotechnol. 2014, 33, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Chen, D.J. DNA double strand break repair via non-homologous end-joining. Transl. Cancer Res. 2013, 2, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Weber, T.; Wefers, B.; Wurst, W.; Sander, S.; Rajewsky, K.; Kühn, R. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nat. Biotechnol. 2015, 33, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huang, J. DNA End Resection: Facts and Mechanisms. Genom. Proteom. Bioinform. 2016, 14, 126–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh-Gohari, N.; Helleday, T. Conservative homologous recombination preferentially repairs DNA double-strand breaks in the S phase of the cell cycle in human cells. Nucleic Acids Res. 2004, 32, 3683–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharati, J.; Punetha, M.; Kumar, B.S.; Vidyalakshmi, G.; Sarkar, M.; D’Occhio, M.J.; Singh, R.K. Genome editing in animals: An overview. In Genomics and Biotechnological Advances in Veterinary, Poultry, and Fisheries; Elsevier Inc.: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Choo, Y.; Sánchez-García, I.; Klug, A. In vivo repression by a site-specific DNA-binding protein designed against an oncogenic sequence. Nature 1994, 372, 642–645. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Urnov, F.D.; Rebar, E.J.; Holmes, M.C.; Zhang, H.S.; Gregory, P.D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 2010, 11, 636–646. [Google Scholar] [CrossRef]
- Tang, L.-M.; Zhou, C.-L.; Guo, Z.-F.; Xiao, L.; Chüeh, A.C. Advances in Zinc Finger Nuclease and Its Applications. Gene Gene Ed. 2015, 1, 3–15. [Google Scholar] [CrossRef]
- Carroll, D. Genome Engineering with Targetable Nucleases. Annu. Rev. Biochem. 2014, 83, 409–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, S.; Larochelle, M.; Turcotte, B. A Fungal Family of Transcriptional Regulators: The Zinc Cluster Proteins. Microbiol. Mol. Biol. Rev. 2006, 70, 583–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Segal, D.J.; Ghiara, J.B.; Barbas, C.F. Design of polydactyl zinc-finger proteins for unique addressing within complex genomes. Proc. Natl. Acad. Sci. USA 1997, 94, 5525–5530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palpant, N.J.; Dudzinski, D.M. Zinc finger nucleases: Looking toward translation. Gene Ther. 2013, 20, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Noyes, M.B.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Urnov, F.D.; Miller, J.C.; Lee, Y.-L.; Beausejour, C.M.; Rock, J.M.; Augustus, S.; Jamieson, A.C.; Porteus, M.H.; Gregory, P.D.; Holmes, M.C. Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature 2005, 435, 646–651. [Google Scholar] [CrossRef]
- Hockemeyer, D.; Soldner, F.; Beard, C.; Gao, Q.; Mitalipova, M.; DeKelver, R.C.; Katibah, G.E.; Amora, R.; Boydston, E.A.; Zeitler, B.; et al. Efficient targeting of expressed and silent genes in human ESCs and iPSCs using zinc-finger nucleases. Nat. Biotechnol. 2009, 27, 851–857. [Google Scholar] [CrossRef] [Green Version]
- Joung, J.K.; Sander, J.D. TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef]
- Boch, J.; Bonas, U. Xanthomonas AvrBs3 Family-Type III Effectors: Discovery and Function. Annu. Rev. Phytopathol. 2010, 48, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Lamb, B.M.; Mercer, A.C.; Barbas, C.F. Directed evolution of the TALE N-terminal domain for recognition of all 5′ bases. Nucleic Acids Res. 2013, 41, 9779–9785. [Google Scholar] [CrossRef] [Green Version]
- Mahfouz, M.M.; Li, L.; Shamimuzzaman; Wibowo, A.; Fang, X.; Zhu, J.-K. De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks. Proc. Natl. Acad. Sci. USA 2011, 108, 2623–2628. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Crudele, J.M.; Chamberlain, J.S. Cas9 immunity creates challenges for CRISPR gene editing therapies. Nat. Commun. 2018, 9, 3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.S.; Southgate, P.C.; Tucker, C.S. Aquaculture: Farming Aquatic Animals and Plants, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Pankaj, C.Y. Cell Stem Cell. Sci Rep. 2014, 331–335. [Google Scholar]
- Mali, P.; Esvelt, K.M.; Church, G. Cas9 as a versatile tool for engineering biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [Green Version]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; Van Der Oost, J. Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR–Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedenheft, B.; Lander, G.C.; Zhou, K.; Jore, M.M.; Brouns, S.J.J.; Van Der Oost, J.; Doudna, J.A.; Nogales, E. Structures of the RNA-guided surveillance complex from a bacterial immune system. Nature 2011, 477, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, D. Gene editing nuclease and its application in tilapia. Sci. Bull. 2017, 62, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 endonukleases reveal RNA-mediated conformational activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [Green Version]
- Gersbach, C.A. Genome engineering: The next genomic revolution. Nat. Methods 2014, 11, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- Lees-Miller, S.P.; Meek, K. Repair of DNA double strand breaks by non-homologous end joining. Biochimie 2003, 85, 1161–1173. [Google Scholar] [CrossRef]
- Singh, I.P.; Hasan, S.; Saxena, P. RNAi: Gene Silencing. Int. J. Res. Eng. Sci. Manag. 2019, 52–54. [Google Scholar]
- Hood, E. RNAi: What’s all the noise about gene silencing? Environ. Health Perspect. 2004, 112, A224–A229. [Google Scholar] [CrossRef]
- Daneholt, B. Advanced Information on the Nobel Prize in Physiology or Medicine 2006; The Nobel Assembly at Karolinska Institute Karolinska Institute: Stockholm, Sweden, 2006. [Google Scholar]
- Estrada, M.P.; Lugo, J.M.; Carpio, Y. RNAi in fish and crustaceans. In RNA Interference: Methods for Plants and Animals; CABI: Wallingford, UK, 2008. [Google Scholar] [CrossRef]
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Chaudhary, S.; Kumar, R.; Sirohi, P.; Mehla, K.; Sirohi, A.; Kumar, S.; Chand, P.; Singh, P.K. RNA Interference Technology—Applications and Limitations. In RNA Interference; Opentech: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, R.; Rajasekaran, K.; Cary, J.W. RNA Interference (RNAi) as a Potential Tool for Control of Mycotoxin Contamination in Crop Plants: Concepts and Considerations. Front. Plant Sci. 2017, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Parveen, P.; Brundavani, K.D.; Mahathi, K.; Bhavani, M.S.; Shaheda Sultana, S.K. Gene Silencing and DNA Methylation. Am. J. Phytomedicine Clin. Therapeutics. 2019, 2, 1–12. [Google Scholar]
- Walsh, A.S.; Yin, H.; Erben, C.M.; Wood, M.J.A.; Turberfield, A.J. DNA Cage Delivery to Mammalian Cells. ACS Nano 2011, 5, 5427–5432. [Google Scholar] [CrossRef] [PubMed]
- Keum, J.-W.; Ahn, J.-H.; Bermudez, H. Design, Assembly, and Activity of Antisense DNA Nanostructures. Small 2011, 7, 3529–3535. [Google Scholar] [CrossRef] [PubMed]
- Tirasophon, W.; Roshorm, Y.; Panyim, S. Silencing of yellow head virus replication in penaeid shrimp cells by dsRNA. Biochem. Biophys. Res. Commun. 2005, 334, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Tiu, S.H.-K.; Chan, S.-M. The use of recombinant protein and RNA interference approaches to study the reproductive functions of a gonad-stimulating hormone from the shrimp Metapenaeus ensis. FEBS J. 2007, 274, 4385–4395. [Google Scholar] [CrossRef]
- Naito, Y.; Yamada, T.; Ui-Tei, K.; Morishita, S.; Saigo, K. siDirect: Highly effective, target-specific siRNA design software for mammalian RNA interference. Nucleic Acids Res. 2004, 32, W124–W129. [Google Scholar] [CrossRef] [Green Version]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef] [PubMed]
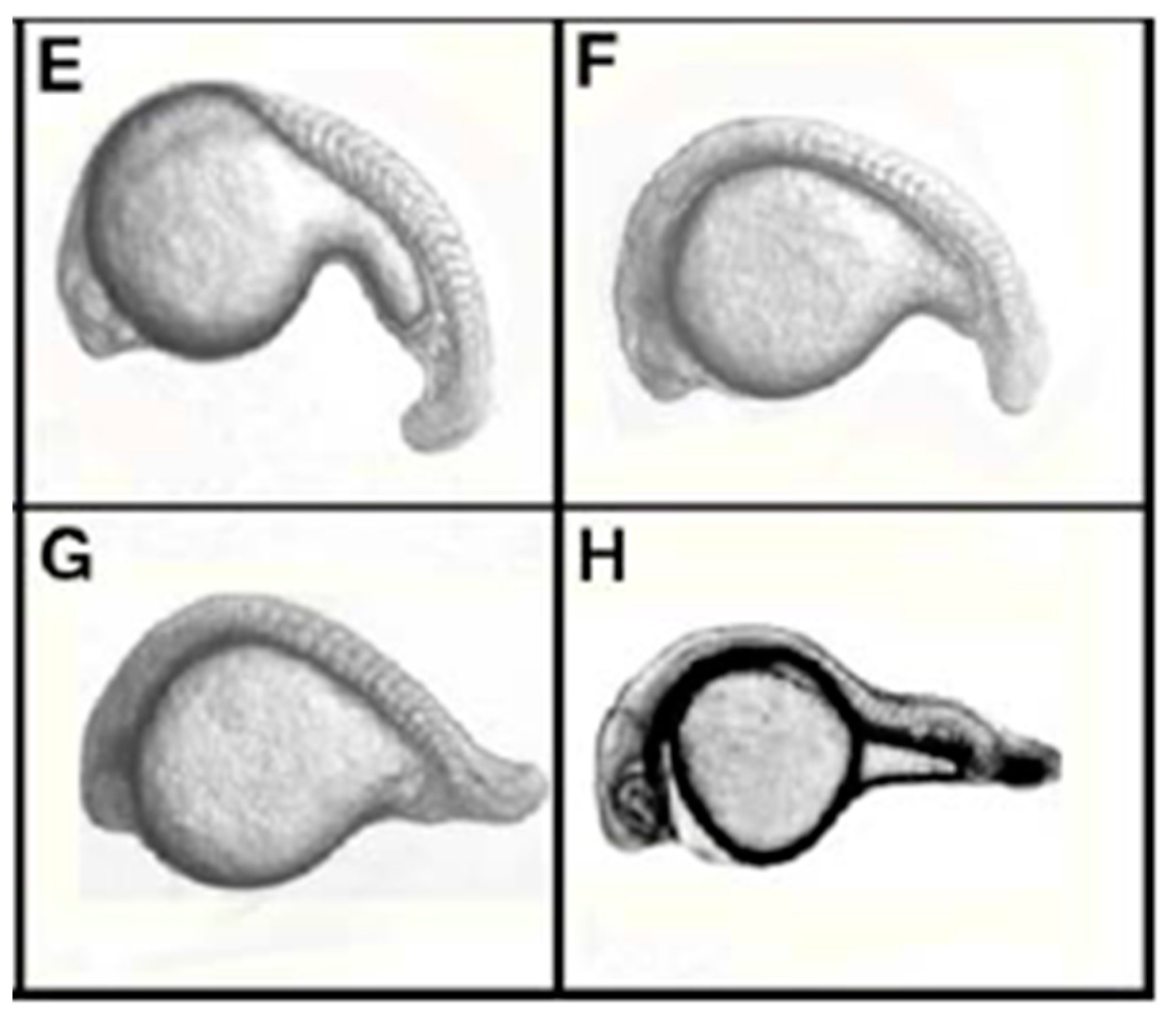
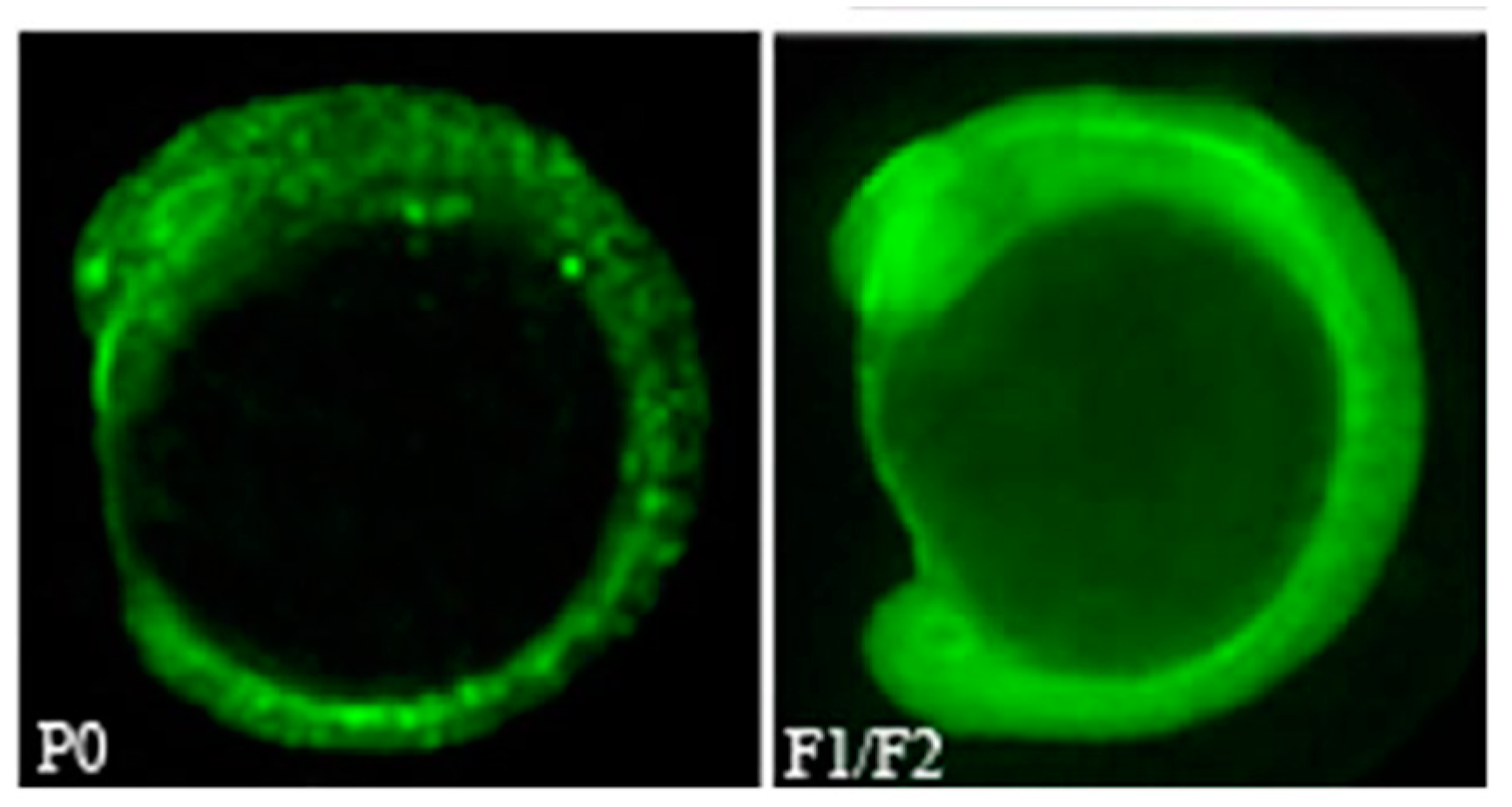
- Wang, N.; Sun, Y.-H.; Liu, J.; Wu, G.; Su, J.-G.; Wang, Y.-P.; Zhu, Z.-Y. Knock down of gfp and no tail expression in zebrafish embryo by in vivo-transcribed short hairpin RNA with T7 plasmid system. J. Biomed. Sci. 2007, 14, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Gotesman, M.; Menanteau-Ledouble, S.; Saleh, M.; Bergmann, S.M.; El-Matbouli, M. A new age in AquaMedicine: Unconventional approach in studying aquatic diseases. BMC Vet. Res. 2018, 14, 178. [Google Scholar] [CrossRef]
- Okoli, A.S.; Blix, T.; Myhr, A.I.; Xu, W.; Xu, X. Sustainable use of CRISPR/Cas in fish aquaculture: The biosafety perspective. Transgenic Res. 2021, 31, 1–21. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shi, Z.; Cui, Y.; Guo, X.; Shi, Y.-B.; Chen, Y. Targeted gene disruption in Xenopus laevis using CRISPR/Cas9. Cell Biosci. 2015, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Doyon, Y.; McCammon, J.M.; Miller, J.C.; Faraji, F.; Ngo, C.; Katibah, G.E.; Amora, R.; Hocking, T.D.; Zhang, L.; Rebar, E.J.; et al. Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 702–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auer, T.O.; Duroure, K.; De Cian, A.; Concordet, J.-P.; Del Bene, F. Highly efficient CRISPR/Cas9-mediated knock-in in zebrafish by homology-independent DNA repair. Genome Res. 2014, 24, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edvardsen, R.; Leininger, S.; Kleppe, L.; Skaftnesmo, K.O.; Wargelius, A. Targeted Mutagenesis in Atlantic Salmon (Salmo salar L.) Using the CRISPR/Cas9 System Induces Complete Knockout Individuals in the F0 Generation. PLoS ONE 2014, 9, e108622. [Google Scholar] [CrossRef]
- Qiu, C.; Cheng, B.; Zhang, Y.; Huang, R.; Liao, L.; Li, Y.; Luo, D.; Hu, W.; Wang, Y. Efficient Knockout of Transplanted Green Fluorescent Protein Gene in Medaka Using TALENs. Mar. Biotechnol. 2014, 16, 674–683. [Google Scholar] [CrossRef]
- Li, M.; Yang, H.; Zhao, J.; Fang, L.; Shi, H.; Li, M.; Sun, Y.; Zhang, X.; Jiang, D.; Zhou, L.; et al. Efficient and Heritable Gene Targeting in Tilapia by CRISPR/Cas9. Genetics 2014, 197, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef]
- Karigo, T.; Aikawa, M.; Kondo, C.; Abe, H.; Kanda, S.; Oka, Y. Whole Brain-Pituitary In Vitro Preparation of the Transgenic Medaka (Oryzias latipes) as a Tool for Analyzing the Differential Regulatory Mechanisms of LH and FSH Release. Endocrinology 2014, 155, 536–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karigo, T.; Kanda, S.; Takahashi, A.; Abe, H.; Okubo, K.; Oka, Y. Time-of-Day-Dependent Changes in GnRH1 Neuronal Activities and Gonadotropin mRNA Expression in a Daily Spawning Fish, Medaka. Endocrinology 2012, 153, 3394–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattanach, B.M.; Iddon, C.A.; Charlton, H.M.; Chiappa, S.A.; Fink, G. Gonadotrophin-releasing hormone deficiency in a mutant mouse with hypogonadism. Nature 1977, 269, 338–340. [Google Scholar] [CrossRef]
- Takahashi, A.; Kanda, S.; Abe, T.; Oka, Y. Evolution of the Hypothalamic-Pituitary-Gonadal Axis Regulation in Vertebrates Revealed by Knockout Medaka. Endocrinology 2016, 157, 3994–4002. [Google Scholar] [CrossRef]
- Qin, Z.; Li, Y.; Su, B.; Cheng, Q.; Ye, Z.; Perera, D.A.; Fobes, M.; Shang, M.; Dunham, R.A. Editing of the Luteinizing Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Zinc Finger Nuclease Technology with Electroporation. Mar. Biotechnol. 2016, 18, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Taranger, G.L.; Karlsen, Ø.; Bannister, R.J.; Glover, K.A.; Husa, V.; Karlsbakk, E.; Kvamme, B.O.; Boxaspen, K.K.; Bjørn, P.A.; Finstad, B.; et al. Risk assessment of the environmental impact of Norwegian Atlantic salmon farming. ICES J. Mar. Sci. 2015, 72, 997–1021. [Google Scholar] [CrossRef] [Green Version]
- Glover, K.A.; Quintela, M.; Wennevik, V.; Besnier, F.; Sørvik, A.G.E.; Skaala, Ø. Three Decades of Farmed Escapees in the Wild: A Spatio-Temporal Analysis of Atlantic Salmon Population Genetic Structure throughout Norway. PLoS ONE 2012, 7, e43129. [Google Scholar] [CrossRef] [Green Version]
- Wargelius, A.; Leininger, S.; Skaftnesmo, K.O.; Kleppe, L.; Andersson, E.; Taranger, G.L.; Schulz, R.W.; Edvardsen, R.B. Dnd knockout ablates germ cells and demonstrates germ cell independent sex differentiation in Atlantic salmon. Sci. Rep. 2016, 6, 21284. [Google Scholar] [CrossRef] [Green Version]
- Youngren, K.K.; Coveney, D.; Peng, X.; Bhattacharya, C.; Schmidt, L.S.; Nickerson, M.L.; Lamb, B.T.; Deng, J.M.; Behringer, R.R.; Capel, B.; et al. The Ter mutation in the dead end gene causes germ cell loss and testicular germ cell tumours. Nature 2005, 435, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Dungan, H.M.; Clifton, D.K.; Steiner, R.A. Minireview: Kisspeptin Neurons as Central Processors in the Regulation of Gonadotropin-Releasing Hormone Secretion. Endocrinology 2006, 147, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.K.; Nguyen, T.; O’Neill, G.P.; Cheng, R.; Liu, Y.; Howard, A.D.; Coulombe, N.; Tan, C.P.; Tang-Nguyen, A.-T.; George, S.R.; et al. Discovery of a receptor related to the galanin receptors. FEBS Lett. 1999, 446, 103–107. [Google Scholar] [CrossRef]
- Popa, S.M.; Clifton, D.K.; Steiner, R.A. The Role of Kisspeptins and GPR54 in the Neuroendocrine Regulation of Reproduction. Annu. Rev. Physiol. 2008, 70, 213–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, A.E.; Clifton, D.K.; Steiner, R.A. Kisspeptin Signaling in the Brain. Endocr. Rev. 2009, 30, 713–743. [Google Scholar] [CrossRef] [PubMed]
- Roa, J.; Aguilar, E.; Dieguez, C.; Pinilla, L.; Tena-Sempere, M. New frontiers in kisspeptin/GPR54 physiology as fundamental gatekeepers of reproductive function. Front. Neuroendocr. 2008, 29, 48–69. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Liu, Y.; Luo, D.; Ogawa, S.; Yin, Y.; Li, S.; Zhang, Y.; Hu, W.; Parhar, I.S.; Lin, H.; et al. The kiss/kissr Systems Are Dispensable for Zebrafish Reproduction: Evidence From Gene Knockout Studies. Endocrinology 2015, 156, 589–599. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Niu, P.; Wang, M.; Huang, G.; Xu, S.; Sun, Y.; Xu, X.; Hou, Y.; Sun, X.; Yan, Y.; et al. Targeted disruption of sp7 and myostatin with CRISPR-Cas9 results in severe bone defects and more muscular cells in common carp. Sci. Rep. 2016, 6, 22953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlem, T.J.; Hoshijima, K.; Jurynec, M.J.; Gunther, D.; Starker, C.; Locke, A.S.; Weis, A.; Voytas, D.; Grunwald, D.J. Simple Methods for Generating and Detecting Locus-Specific Mutations Induced with TALENs in the Zebrafish Genome. PLOS Genet. 2012, 8, e1002861. [Google Scholar] [CrossRef] [Green Version]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Jeffery, W.R.; Essner, J.J.; Kowalko, J.E. Genome Editing Using TALENs in Blind Mexican Cavefish, Astyanax mexicanus. PLoS ONE 2015, 10, e0119370. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, P.R.; Campbell, J.M.; Clark, K.J.; Ekker, S.C. The CRISPR System—Keeping Zebrafish Gene Targeting Fresh. Zebrafish 2013, 10, 116–118. [Google Scholar] [CrossRef] [Green Version]
- Segev-Hadar, A.; Slosman, T.; Rozen, A.; Sherman, A.; Cnaani, A.; Biran, J. Genome Editing Using the CRISPR-Cas9 System to Generate a Solid-Red Germline of Nile Tilapia (Oreochromis niloticus). CRISPR J. 2021, 4, 583–594. [Google Scholar] [CrossRef]
- Souza, T.A.; Chen, X.; Guo, Y.; Sava, P.; Zhang, J.; Hill, J.J.; Yaworsky, P.J.; Qiu, Y. Proteomic Identification and Functional Validation of Activins and Bone Morphogenetic Protein 11 as Candidate Novel Muscle Mass Regulators. Mol. Endocrinol. 2008, 22, 2689–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, K.; Elayat, M.; Khalifa, E.; Daghash, S.; Elaswad, A.; Miller, M.; Abdelrahman, H.; Ye, Z.; Odin, R.; Drescher, D.; et al. Generation of Myostatin Gene-Edited Channel Catfish (Ictalurus punctatus) via Zygote Injection of CRISPR/Cas9 System. Sci. Rep. 2017, 7, 7301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilley, J.H.; Callinan, R.B.; Chinabut, S.; Kanchanakhan, S.; MacRae, I.H. Epizootic Ulcerative Syndrome (EUS) Technical Handbook; The Aquatic Animal Health Research Institute AARI Department of Fisheries, Kasetsart University: Bangkok, Thailand, 1998; Volume 53. [Google Scholar]
- OIE (World Organisation for Animal Health) Listed Diseases. 2017. Available online: https://www.oie.int/en/what-we-do/animal-health-and-welfare/animal-diseases/ (accessed on 7 May 2022).
- Yike, I. Fungal Proteases and Their Pathophysiological Effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef]
- Majeed, M.; Kumar, G.; Schlosser, S.; El-Matbouli, M.; Saleh, M. In vitro investigations on extracellular proteins secreted by Aphanomyces invadans, the causative agent of epizootic ulcerative syndrome. Acta Vet. Scand. 2017, 59, 78. [Google Scholar] [CrossRef] [Green Version]
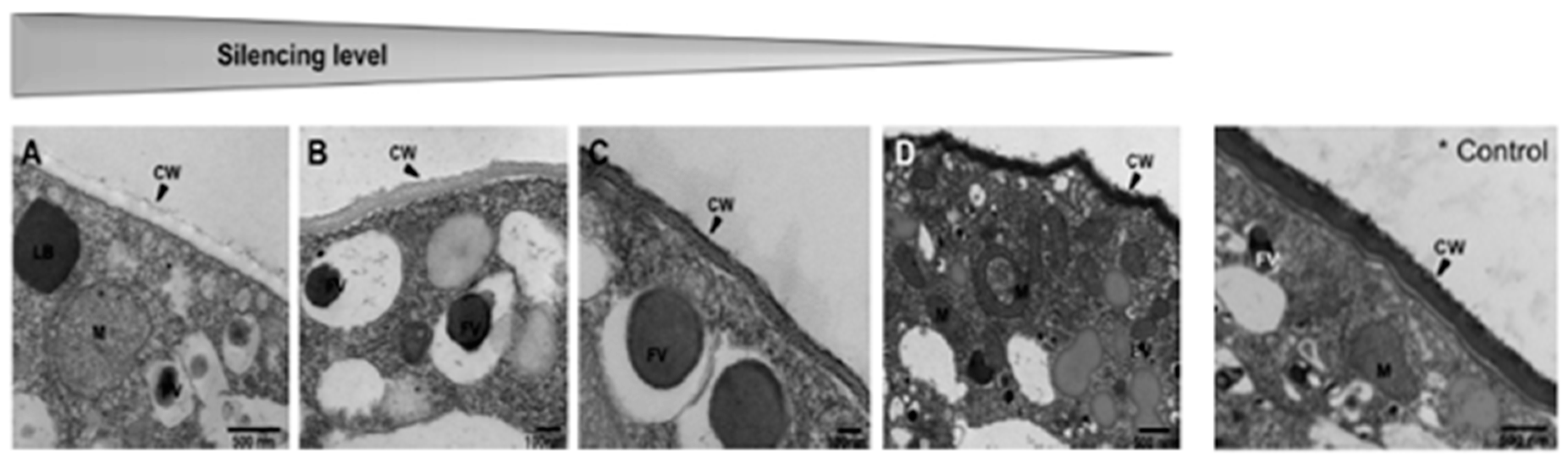
- Majeed, M.; Soliman, H.; Kumar, G.; El-Matbouli, M.; Saleh, M. Editing the genome of Aphanomyces invadans using CRISPR/Cas9. Parasites Vectors 2018, 11, 554. [Google Scholar] [CrossRef]
- Roberts, R.J.; Willoughby, L.G.; Chinabut, S. Mycotic aspects of epizootic ulcerative syndrome (EUS) of Asian fishes. J. Fish Dis. 1993, 16, 169–183. [Google Scholar] [CrossRef]
- Pathiratne, A.; Widanapathirana, G.S.; Chandrakanthi, W.H.S. Association of Aeromonas hydrophila with epizootic ulcerative syndrome (EUS) of freshwater fish in Sri Lanka. J. Appl. Ichthyol. 1994, 10, 204–208. [Google Scholar] [CrossRef]
- Lilley, J.H.; Roberts, R.J. Pathogenicity and culture studies comparing the Aphanomyces involved in epizootic ulcerative syndrome (EUS) with other similar fungi. J. Fish Dis. 1997, 20, 135–144. [Google Scholar] [CrossRef]
- Vishwanath, T.; Mohan, C.; Shankar, K. Epizootic Ulcerative Syndrome (EUS), associated with a fungal pathogen, in Indian fishes: Histopathology—‘A cause for invasiveness’. Aquaculture 1998, 165, 1–9. [Google Scholar] [CrossRef]
- Biacchesi, S. The reverse genetics applied to fish RNA viruses. Vet. Res. 2011, 42, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reshi, M.L.; Wu, J.-L.; Wang, H.-V.; Hong, J.-R. RNA interference technology used for the study of aquatic virus infections. Fish Shellfish. Immunol. 2014, 40, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.C.; Harris, J.O.; Cook, M. Exploring RNAi as a therapeutic strategy for controlling disease in aquaculture. Fish Shellfish. Immunol. 2013, 34, 729–743. [Google Scholar] [CrossRef]
- Carpio, Y.; Estrada, M.P. Zebrafish as a genetic model organism. Biotecnol. Apl. 2006, 23, 265–270. [Google Scholar]
- Hammond, S.M. MicroRNA therapeutics: A new niche for antisense nucleic acids. Trends Mol. Med. 2006, 12, 99–101. [Google Scholar] [CrossRef]
- Gotesman, M.; Soliman, H.; Besch, R.; El-Matbouli, M. Inhibition of spring viraemia of carp virus replication in an E pithelioma papulosum cyprini cell line by RNA i. J. Fish Dis. 2015, 38, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Gotesman, M.; Soliman, H.; Besch, R.; El-Matbouli, M. In vitro inhibition of Cyprinid herpesvirus-3 replication by RNAi. J. Virol. Methods 2014, 206, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Adamek, M.; Rauch, G.; Brogden, G.; Steinhagen, D. Small interfering RNA treatment can inhibit Cyprinid herpesvirus 3 associated cell death in vitro. Pol. J. Vet. Sci. 2014, 17, 733–735. [Google Scholar] [CrossRef]
- Saleh, M.; Kumar, G.; Abdel-Baki, A.-A.; Dkhil, M.A.; El-Matbouli, M.; Al-Quraishy, S. In Vitro Gene Silencing of the Fish Microsporidian Heterosporis saurida by RNA Interference. Nucleic Acid Ther. 2016, 26, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, B. Über die Drehkrankheit der Regenbogenforelle. Allg Fisch-Ztg. 1903, 28, 7–8. [Google Scholar]
- Sarker, S.; El-Matbouli, M. Can RNAi Target Salmonid Whirling Disease In Vivo? Nucleic Acid Ther. 2015, 25, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.; Kallert, D.; Hedrick, R.; El-Matbouli, M. Whirling disease revisited: Pathogenesis, parasite biology and disease intervention. Dis. Aquat. Org. 2015, 114, 155–175. [Google Scholar] [CrossRef]
- Sarker, S.; Menanteau-Ledouble, S.; Kotob, M.H.; El-Matbouli, M. A RNAi-based therapeutic proof of concept targets salmonid whirling disease in vivo. PLoS ONE 2017, 12, e0178687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rienzo, G.; Gutzman, J.H.; Sive, H.; Don, E.K.; Formella, I.; Badrock, A.P.; Hall, T.E.; Morsch, M.; Hortle, E.; Hogan, A.; et al. Efficient shRNA-Mediated Inhibition of Gene Expression in Zebrafish. Zebrafish 2012, 9, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Gruber, J.; Manninga, H.; Tuschl, T.; Osborn, M.; Weber, K. Specific RNAi Mediated Gene Knockdown in Zebrafish Cell Lines. RNA Biol. 2005, 2, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; de Bruijn, I.; Grenville-Briggs, L.; McLaggan, D.; Willems, A.; Bulone, V.; van West, P. Functional characterization of a tyrosinase gene from the oomycete Saprolegnia parasitica by RNAi silencing. Fungal Biol. 2014, 118, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Capodici, J.; Karikó, K.; Weissman, D. Inhibition of HIV-1 Infection by Small Interfering RNA-Mediated RNA Interference. J. Immunol. 2002, 169, 5196–5201. [Google Scholar] [CrossRef] [Green Version]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. elegans Requires the Putative Transmembrane Protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, E.H.; Hunter, C.P. Transport of dsRNA into Cells by the Transmembrane Protein SID-1. Science 2003, 301, 1545–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishok, A. RNAi mechanisms in Caenorhabditis elegans. FEBS Lett. 2005, 579, 5932–5939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flegel, T. Major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microbiol. Biotechnol. 1997, 13, 433–442. [Google Scholar] [CrossRef]
- Lightner, D.V.; Hasson, K.W.; White, B.L.; Redman, R.M. Experimental infection of western hemisphere penaeid shrimp with asian white spot syndrome virus and asian yellow head virus. J. Aquat. Anim. Health 1998, 10, 271–281. [Google Scholar] [CrossRef]
- Assavalapsakul, W.; Smith, D.R.; Panyim, S. Identification and Characterization of a Penaeus monodon Lymphoid Cell-Expressed Receptor for the Yellow Head Virus. J. Virol. 2006, 80, 262–269. [Google Scholar] [CrossRef] [Green Version]
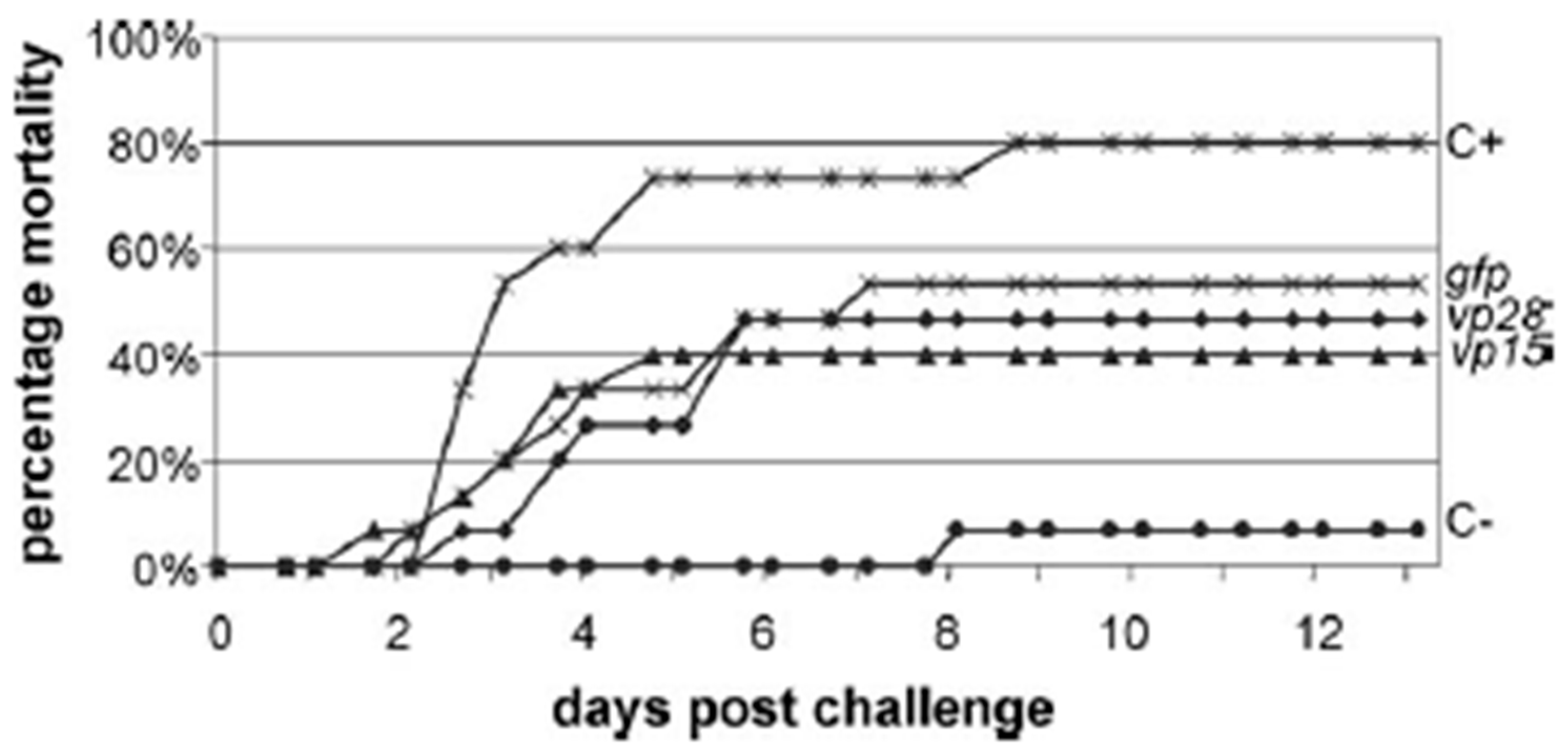
- Westenberg, M.; Heinhuis, B.; Zuidema, D.; Vlak, J.M. siRNA injection induces sequence-independent protection in Penaeus monodon against white spot syndrome virus. Virus Res. 2005, 114, 133–139. [Google Scholar] [CrossRef]
- Witteveldt, J.; Vermeesch, A.M.G.; Langenhof, M.; de Lang, A.; Vlak, J.M.; van Hulten, M.C.W. Nucleocapsid protein VP15 is the basic DNA binding protein of white spot syndrome virus of shrimp. Arch. Virol. 2005, 150, 1121–1133. [Google Scholar] [CrossRef]
- Van Hulten, M.C.; Witteveldt, J.; Snippe, M.; Vlak, J.M. White Spot Syndrome Virus Envelope Protein VP28 Is Involved in the Systemic Infection of Shrimp. Virology 2001, 285, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Jiravanichpaisal, P.; Söderhäll, I.; Cerenius, L.; Söderhäll, K. Antilipopolysaccharide Factor Interferes with White Spot Syndrome Virus Replication In Vitro and In Vivo in the Crayfish Pacifastacus leniusculus. J. Virol. 2006, 80, 10365–10371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Jiravanichpaisal, P.; Cerenius, L.; Lee, B.L.; Söderhäll, I.; Söderhäll, K. Phenoloxidase Is an Important Component of the Defense against Aeromonas hydrophila Infection in a Crustacean, Pacifastacus leniusculus. J. Biol. Chem. 2007, 282, 33593–33598. [Google Scholar] [CrossRef] [Green Version]
- Soyez, D.; Noel, P.; Van Deijnen, J.; Martin, M.; Morel, A.; Payen, G. Neuropeptides from the sinus gland of the lobster Homarus americanus: Characterization of hyperglycemic peptides. Gen. Comp. Endocrinol. 1990, 79, 261–274. [Google Scholar] [CrossRef]
- Lugo, J.M.; Morera, Y.; Rodríguez, T.; Huberman, A.; Ramos, L.; Estrada, M.P. Molecular cloning and characterization of the crustacean hyperglycemic hormone cDNA from Litopenaeus schmitti: Functional analysis by double-stranded RNA interference technique. FEBS J. 2006, 273, 5669–5677. [Google Scholar] [CrossRef]
- Copf, T.; Rabet, N.; Averof, M. Knockdown of spalt function by RNAi causes de-repression of Hox genes and homeotic transformations in the crustacean Artemia franciscana. Dev. Biol. 2006, 298, 87–94. [Google Scholar] [CrossRef]
- Treerattrakool, S.; Panyim, S.; Chan, S.-M.; Withyachumnarnkul, B.; Udomkit, A. Molecular characterization of gonad-inhibiting hormone of Penaeus monodon and elucidation of its inhibitory role in vitellogenin expression by RNA interference. FEBS J. 2008, 275, 970–980. [Google Scholar] [CrossRef]
- Keller, R. Crustacean neuropeptides: Structures, functions and comparative aspects. Experientia 1992, 48, 439–448. [Google Scholar] [CrossRef]
- De Kleijn, D.P.; Van Herp, F. Molecular biology of neurohormone precursors in the eyestalk of Crustacea. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 112, 573–579. [Google Scholar] [CrossRef]
- Chan, S.-M.; Gu, P.-L.; Chu, K.H.; Tobe, S.S. Crustacean neuropeptide genes of the CHH/MIH/GIH family: Implications from molecular studies. Gen. Comp. Endocrinol. 2003, 134, 214–219. [Google Scholar] [CrossRef]
- Yang, L.; Güell, M.; Byrne, S.; Yang, J.L.; Angeles, A.D.L.; Mali, P.; Aach, J.; Kim-Kiselak, C.; Briggs, A.W.; Rios, X.; et al. Optimization of scarless human stem cell genome editing. Nucleic Acids Res. 2013, 41, 9049–9061. [Google Scholar] [CrossRef]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef]
- Ledford, H. CRISPR, the disruptor. Nature 2015, 522, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capps, B.; Chadwick, R.; Joly, Y.; Mulvihill, J.J.; Lysaght, T.; Zwart, H. Falling giants and the rise of gene editing: Ethics, private interests and the public good. Hum. Genom. 2017, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oye, K.A.; Esvelt, K.; Appleton, E.; Catteruccia, F.; Church, G.; Kuiken, T.; Lightfoot, S.B.-Y.; McNamara, J.; Smidler, A.; Collins, J.P. Regulating gene drives. Science 2014, 345, 626–628. [Google Scholar] [CrossRef] [Green Version]
- Collins, J.P. Gene drives in our future: Challenges of and opportunities for using a self-sustaining technology in pest and vector management. BMC Proc. 2018, 12, 37–41. [Google Scholar] [CrossRef]
- Gantz, V.M.; Bier, E. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 2015, 348, 442–444. [Google Scholar] [CrossRef] [Green Version]
- Esvelt, K.M.; Smidler, A.L.; Catteruccia, F.; Church, G.M. Concerning RNA-guided gene drives for the alteration of wild populations. eLife 2014, 3, e03401. [Google Scholar] [CrossRef]
- Hackett, P.B.; Fahrenkrug, S.C.; Carlson, D.F. The Promises and Challenges of Precision Gene Editing in Animals of Agricultural Importance; NABC, NABC report; eCommons: New York, NY, USA, 2014. [Google Scholar]
- Zhu, B.; Ge, W. Genome editing in fishes and their applications. Gen. Comp. Endocrinol. 2018, 257, 3–12. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Zinc Finger Nucleases (Zfns) | Transcription Activator-Like Effector Nucleases (Talens) | Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) |
|---|---|---|---|
| Structure | Fusion of zinc finger DNA-binding domain (DBD) with DNA-cleavage domain of Fok I endonuclease | Fusion of transcription activator-like effector DNA (TALEN) repeats with DNA-cleavage domain of Fok I endonuclease | Cas9 endonuclease and guide RNA (gRNA) |
| Size of recognition site | 9–18 bases in DNA | 30–36 bases in DNA | 23 DNA bases in DNA |
| Ease of designing | More difficult than TALENs and CRISPR | Easier than ZFNs | Easier than the other two |
| Multiplexing | No | No | Yes |
| Off target | Same as that of TALENs | Same as that of ZFNs | More than the other two |
| Ease of redesigning/adaptability to target new site | Difficult, require recording of large DNA segments (500–1000 bp) | Easy, only requires a change in 20-bp protospacer of gRNA | |
| Viral delivery | Using lentivirus and adenovirus; needs cotransduction with two lentiviral vectors, each encoding a monomer to form functional heterodimer | Using adenovirus | More difficult than the other two because it requires polyadenylation signal and promoter |
| Efficacy | + + | + + | + + + |
| Application | Indels, obligate ligation-gated recombination (ObLiGaRe); can insert a 15-kb inducible gene expression cassette at a defined locus in human cell lines, tag ligation | Indels | |
| Cost | Higher than CRISPR | Higher than CRISPR | Less than the other two |
| Type of Molecule | Genes Targeted | RNAi | Response | Organism |
|---|---|---|---|---|
| Specific | Nonspecific | |||
| Long dsRNA | ntl, fl h, pax2.1, LacZ | x | x | Zebrafish embryos a |
| Long dsRNA | Gfp, Zf-T, pax6.1 | x | Zebrafish embryos | |
| Long dsRNA | Tbx16/spt, LacZ | x | Zebrafish embryos | |
| Long dsRNA | pouII-1, gfp, terra | x | Zebrafish embryos | |
| Long dsRNA | RanBP1 | x | Zebrafish embryos | |
| Long dsRNA | M2mAchR | x | Zebrafish embryos | |
| siRNA | gfp, tyrA | x | Rainbow trout embryos b | |
| siRNA | Dmd | x | Zebrafish embryos | |
| esiRNA | Ntl | x | Zebrafish embryos | |
| siRNA | Ntl | x | Zebrafish embryos | |
| siRNA | laminA and B2, Eg5, GL2, gfp | x | ZFL, SJD and ZF4 c | |
| siRNA | laminA, GL2, gfp | x | Zebrafish embryos | |
| Long dsRNA | Myostatin | x | Zebrafish embryos | |
| T7RPshRNA | ntl, gfp | x | Zebrafish embryos |
| RNA | Target | Genes | RNAi Response | Organism |
|---|---|---|---|---|
| dsRNA | Endogenous A | Spalt | Pleiotropic effects | Artemia franciscana |
| dsRNA | Chh | Decrease in glucose levels | Litopenaeus schmitti | |
| dsRNA | ALF | Protection against WSSV | Pacifastacus leniusculus | |
| dsRNA | pmYRP65 | Inhibition of YHV cell entry | Penaeus monodon C | |
| dsRNA | proPO | Increased bacterial D growth | Pacifastacus leniusculus | |
| dsRNA | Pacifastin | Decreased bacterial D growth | Pacifastacus leniusculus | |
| dsRNA | Mih-B | Reduction of vitellogenin gene | Metapenaeus ensis | |
| dsRNA | Pem-GIH | Decrease in Pem-GIH transcripts and reduction of vitellogenin gene | Penaeus monodon | |
| dsRNA | Virals B and (unrelated) | hel, pol, pro, gp116, gp64 | Inhibition of YHV replication | Penaeus monodon C |
| dsRNA | (gfp) | Non-specific antiviral immunity | ||
| dsRNA | vp28, vp15 | Non-specific antiviral immunity and lower viral protection | Penaeus monodon | |
| dsRNA | Pro | Inhibition of YHV replication | Penaeus monodon | |
| dsRNA | (gfp, TSV pol) | Partial inhibition of YHV replication | ||
| siRNA | (duck u) | Non-specific antiviral immunity | Litopenaeus vannamei | |
| siRNA | vp28 | Non-specific antiviral immunity | Penaeus japonicus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutási, A.; Hammer, S.E.; El-Matbouli, M.; Saleh, M. Review: Recent Applications of Gene Editing in Fish Species and Aquatic Medicine. Animals 2023, 13, 1250. https://doi.org/10.3390/ani13071250
Gutási A, Hammer SE, El-Matbouli M, Saleh M. Review: Recent Applications of Gene Editing in Fish Species and Aquatic Medicine. Animals. 2023; 13(7):1250. https://doi.org/10.3390/ani13071250
Chicago/Turabian StyleGutási, Anikó, Sabine E. Hammer, Mansour El-Matbouli, and Mona Saleh. 2023. "Review: Recent Applications of Gene Editing in Fish Species and Aquatic Medicine" Animals 13, no. 7: 1250. https://doi.org/10.3390/ani13071250