
Heat Inactivation of Methicillin-Resistant Staphylococcus aureus Strains from German Dairy farms in Colostrum and Raw Milk
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Strains and Preparation of the Inoculum
2.2. Estimation of the Detection Limit of the Experimental Set Up
2.3. Thermal Challenge
2.4. Quantification of Thermal Inactivation
2.5. Statistical Analysis
3. Results
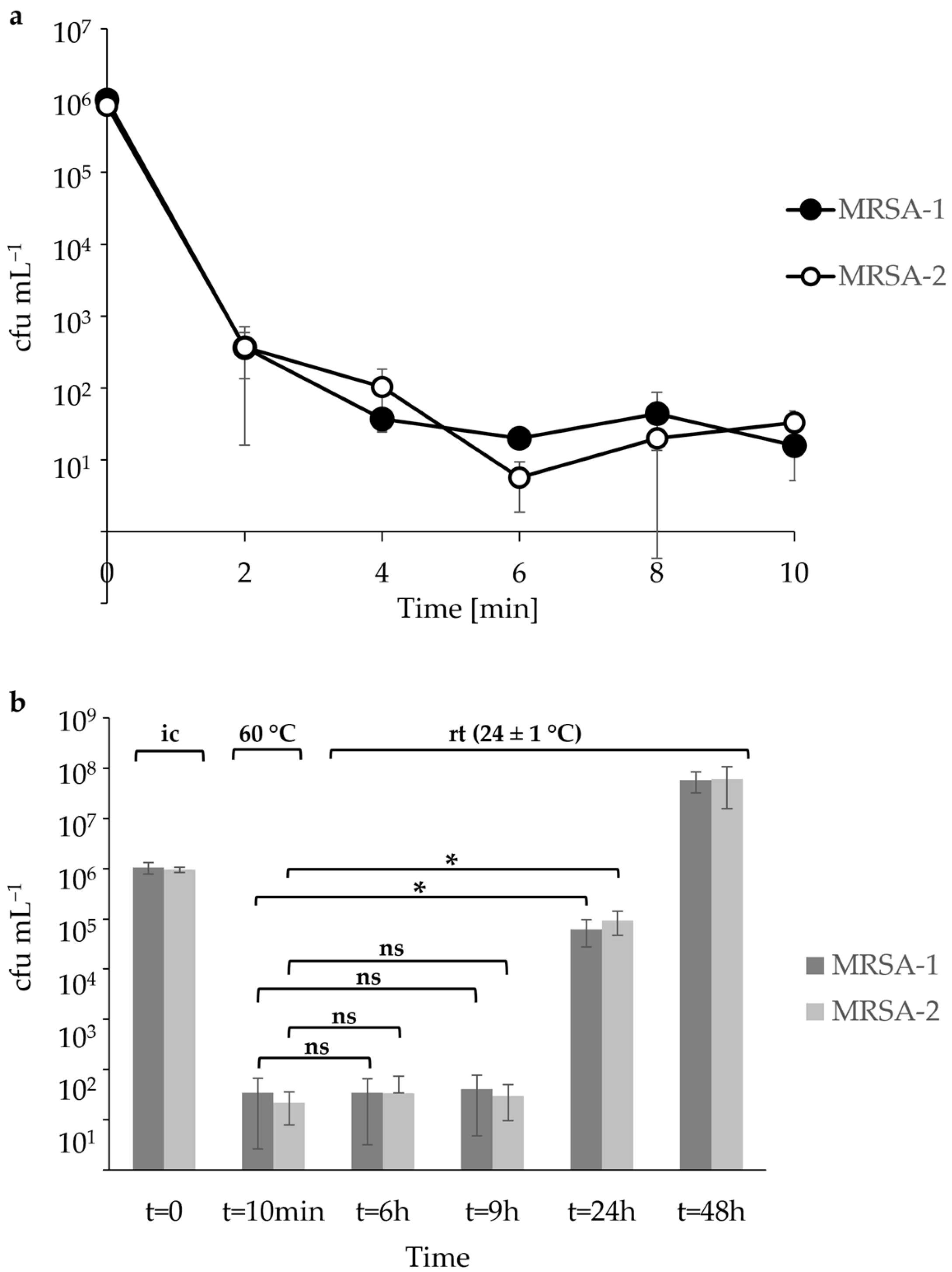
3.1. Heat Inactivation of Low MRSA Levels in Raw Milk and Colostrum
3.2. Heat Inactivation of High MRSA Levels in Raw Milk
3.3. Quantifcation of MRSA Heat Inactivation in Highly Inoculated Raw Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schnitt, A.; Tenhagen, B.-A. Risk Factors for the Occurrence of Methicillin-Resistant Staphylococcus aureus in Dairy Herds: An Update. Foodborne Pathog. Dis. 2019, 17, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Tenhagen, B.-A.; Alt, K.; Pfefferkorn, B.; Wiehle, L.; Käsbohrer, A.; Fetsch, A. Short communication: Methicillin-resistant Staphylococcus aureus in conventional and organic dairy herds in Germany. J. Dairy Sci. 2018, 101, 3380–3386. [Google Scholar] [CrossRef] [PubMed]
- Cuny, C.; Abdelbary, M.; Layer, F.; Werner, G.; Witte, W. Prevalence of the immune evasion gene cluster in Staphylococcus aureus CC398. Vet. Microbiol. 2015, 177, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Goerge, T.; Lorenz, M.B.; van Alen, S.; Hübner, N.-O.; Becker, K.; Köck, R. MRSA colonization and infection among persons with occupational livestock exposure in Europe: Prevalence, preventive options and evidence. Vet. Microbiol. 2017, 200, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-K.; Nam, H.-M.; Jang, G.-C.; Lee, H.-S.; Jung, S.-C.; Kim, T.-S.; Basanisi, M.; La Bella, G.; Nobili, G.; Franconieri, I.; et al. Transmission and Persistence of Methicillin-Resistant Staphylococcus aureus in Milk, Environment, and Workers in Dairy Cattle Farms. Foodborne Pathog. Dis. 2013, 10, 731–736. [Google Scholar] [CrossRef]
- Cuny, C.; Wieler, L.H.; Witte, W. Livestock-Associated MRSA: The Impact on Humans. Antibiotics 2015, 4, 521–543. [Google Scholar] [CrossRef]
- Lienen, T.; Schnitt, A.; Hammerl, J.A.; Maurischat, S.; Tenhagen, B.-A. Genomic Distinctions of LA-MRSA ST398 on Dairy Farms from Different German Federal States With a Low Risk of Severe Human Infections. Front. Microbiol. 2020, 11, 575321. [Google Scholar] [CrossRef]
- Holmes, M.A.; Zadoks, R.N. Methicillin Resistant S. aureus in Human and Bovine Mastitis. J. Mammary Gland. Biol. Neoplasia 2011, 16, 373–382. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef]
- Zigo, F.; Farkašová, Z.; Výrostková, J.; Regecová, I.; Ondrašovičová, S.; Vargová, M.; Sasáková, N.; Pecka-Kielb, E.; Bursová, Š.; Kiss, D.S. Dairy Cows’ Udder Pathogens and Occurrence of Virulence Factors in Staphylococci. Animals 2022, 12, 470. [Google Scholar] [CrossRef]
- Cobirka, M.; Tancin, V.; Slama, P. Epidemiology and Classification of Mastitis. Animals 2020, 10, 2212. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.; McClure, J.; Scholl, D.; DeVries, T.; Barkema, H. Herd-level association between antimicrobial use and antimicrobial resistance in bovine mastitis Staphylococcus aureus isolates on Canadian dairy farms. J. Dairy Sci. 2012, 95, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Zigo, F.; Vasiľ, M.; Elečko, J.; Zigová, M.; Farkašová, Z. Mastitis pathogens and their resistance against antimicrobial agents in herds of dairy cows situated in marginal parts of Slovakia. Potravin. Slovak J. Food Sci. 2018, 12, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Tenhagen, B.A.; Vossenkuhl, B.; Käsbohrer, A.; Alt, K.; Kraushaar, B.; Guerra, B.; Schroeter, A.; Fetsch, A. Methicillin-resistant Staphylococcus aureus in cattle food chains—Prevalence, diversity, and antimicrobial resistance in Germany. J. Anim. Sci. 2014, 92, 2741–2751. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaeghen, W.; Cerpentier, T.; Adriaensen, C.; Vicca, J.; Hermans, K.; Butaye, P. Methicillin-resistant Staphylococcus aureus (MRSA) ST398 associated with clinical and subclinical mastitis in Belgian cows. Vet. Microbiol. 2010, 144, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.N.; Han, S.G. Bovine mastitis: Risk factors, therapeutic strategies, and alternative treatments—A review. Asian-Australas. J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef]
- Moore, D.; Taylor, J.; Hartman, M.; Sischo, W. Quality assessments of waste milk at a calf ranch. J. Dairy Sci. 2009, 92, 3503–3509. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Escámez, P.S.F.; Girones, R.; Koutsoumanis, K.; Lindqvist, R.; et al. Risk for the development of Antimicrobial Resistance (AMR) due to feeding of calves with milk containing residues of antibiotics. EFSA J. 2017, 15, e04665. [Google Scholar] [CrossRef]
- Spohr, M.; Rau, J.; Friedrich, A.; Klittich, G.; Fetsch, A.; Guerra, B.; Hammerl, J.A.; Tenhagen, B.-A. Methicillin-Resistant Staphylococcus aureus (MRSA) in Three Dairy Herds in Southwest Germany. Zoonoses Public Health 2011, 58, 252–261. [Google Scholar] [CrossRef]
- Schnitt, A.; Lienen, T.; Wichmann-Schauer, H.; Cuny, C.; Tenhagen, B.-A. The occurrence and distribution of livestock-associated methicillin-resistant Staphylococcus aureus ST398 on German dairy farms. J. Dairy Sci. 2020, 103, 11806–11819. [Google Scholar] [CrossRef]
- Lienen, T.; Schnitt, A.; Cuny, C.; Maurischat, S.; Tenhagen, B.-A. Phylogenetic Tracking of LA-MRSA ST398 Intra-Farm Transmission among Animals, Humans and the Environment on German Dairy Farms. Microorganisms 2021, 9, 1119. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.; Hurley, D.J.; Reber, A.J. Neonatal Immune Development in the Calf and Its Impact on Vaccine Response. Vet. Clin. North Am. Food Anim. Pract. 2008, 24, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum Management for Dairy Calves. Vet. Clin. North Am. Food Anim. Pract. 2019, 35, 535–556. [Google Scholar] [CrossRef] [PubMed]
- Schnitt, A.; Lienen, T.; Wichmann-Schauer, H.; Tenhagen, B.-A. The occurrence of methicillin-resistant non-aureus staphylococci in samples from cows, young stock, and the environment on German dairy farms. J. Dairy Sci. 2021, 104, 4604–4614. [Google Scholar] [CrossRef]
- Baird-Parker, T.C. Staphylococcus aureus. In The Microbiological Safety and Quality of Food; Lund, B.M., Baird-Parker, T.C., Gould, G.W., Eds.; Aspen Publishers: Gaithersburg, MA, USA, 2000; pp. 1317–1330. [Google Scholar]
- Gebremedhin, E.Z.; Ararso, A.B.; Borana, B.M.; Kelbesa, K.A.; Tadese, N.D.; Marami, L.M.; Sarba, E.J. Isolation and Identification of Staphylococcus aureus from Milk and Milk Products, Associated Factors for Contamination, and Their Antibiogram in Holeta, Central Ethiopia. Veter-Med. Int. 2022, 2022, 6544705. [Google Scholar] [CrossRef]
- Lucey, J.A. Raw Milk Consumption: Risks and Benefits. Nutr. Today 2015, 50, 189–193. [Google Scholar] [CrossRef]
- Penati, M.; Sala, G.; Biscarini, F.; Boccardo, A.; Bronzo, V.; Castiglioni, B.; Cremonesi, P.; Moroni, P.; Pravettoni, D.; Addis, M.F. Feeding Pre-weaned Calves with Waste Milk Containing Antibiotic Residues Is Related to a Higher Incidence of Diarrhea and Alterations in the Fecal Microbiota. Front. Vet. Sci. 2021, 8, 650150. [Google Scholar] [CrossRef]
- Espinosa, M.F.; Sancho, A.N.; Mendoza, L.M.; Mota, C.R.; Verbyla, M.E. Systematic review and meta-analysis of time-temperature pathogen inactivation. Int. J. Hyg. Environ. Health 2020, 230, 113595. [Google Scholar] [CrossRef]
- Elizondo-Salazar, J.; Jayarao, B.; Heinrichs, A. Effect of heat treatment of bovine colostrum on bacterial counts, viscosity, and immunoglobulin G concentration. J. Dairy Sci. 2010, 93, 961–967. [Google Scholar] [CrossRef]
- Arslan, A.; Kaplan, M.; Duman, H.; Bayraktar, A.; Ertürk, M.; Henrick, B.M.; Frese, S.A.; Karav, S. Bovine Colostrum and Its Potential for Human Health and Nutrition. Front. Nutr. 2021, 8, 651721. [Google Scholar] [CrossRef]
- Weaver, D.M.; Tyler, J.W.; VanMetre, D.C.; Hostetler, D.E.; Barrington, G.M. Passive transfer of colostral immunoglobulins in calves. J. Vet. Intern. Med. 2000, 14, 569–577. [Google Scholar] [CrossRef] [PubMed]
- McMartin, S.; Godden, S.; Metzger, L.; Feirtag, J.; Bey, R.; Stabel, J.; Goyal, S.; Fetrow, J.; Wells, S.; Chester-Jones, H. Heat Treatment of Bovine Colostrum. I: Effects of Temperature on Viscosity and Immunoglobulin G Level. J. Dairy Sci. 2006, 89, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- Donahue, M.; Godden, S.; Bey, R.; Wells, S.; Oakes, J.; Sreevatsan, S.; Stabel, J.; Fetrow, J. Heat treatment of colostrum on commercial dairy farms decreases colostrum microbial counts while maintaining colostrum immunoglobulin G concentrations. J. Dairy Sci. 2012, 95, 2697–2702. [Google Scholar] [CrossRef] [PubMed]
- Mckay, A.M. Antimicrobial Resistance and Heat Sensitivity of Oxacillin-Resistant, mecA-Positive Staphylococcus spp. from Unpasteurized Milk. J. Food Prot. 2008, 71, 186–190. [Google Scholar] [CrossRef]
- Ugborogho, T.O.; Ingham, S.C. Increased D-values of Staphylococcus aureus resulting from anaerobic heating and enumeration of survivors. Food Microbiol. 1994, 11, 275–280. [Google Scholar] [CrossRef]
- Kennedy, J.; Blair, I.; McDowell, D.; Bolton, D. An investigation of the thermal inactivation of Staphylococcus aureus and the potential for increased thermotolerance as a result of chilled storage. J. Appl. Microbiol. 2005, 99, 1229–1235. [Google Scholar] [CrossRef]
- Amado, I.R.; A Vázquez, J.A.; Guerra, N.P.; Pastrana, L. Thermal resistance of Salmonella enterica, Escherichia coli and Staphylococcus aureus isolated from vegetable feed ingredients. J. Sci. Food Agric. 2014, 94, 2274–2281. [Google Scholar] [CrossRef]
- Li, X.; Sheldon, B.W.; Ball, H.R. Thermal resistance of Salmonella enterica serotypes, Listeria monocytogenes, and Staphylococcus aureus in high solids liquid egg mixes. J. Food Prot. 2005, 68, 703–710. [Google Scholar] [CrossRef]
- Montanari, C.; Serrazanetti, D.I.; Felis, G.; Torriani, S.; Tabanelli, G.; Lanciotti, R.; Gardini, F. New insights in thermal re-sistance fo staphylococcal strains belonging to the species Staphylococcus epidermidis, Staphylococcus lugdunensis and Staphylococcus aureus. Food Control 2015, 50, 605–612. [Google Scholar] [CrossRef]
- Yehia, H.M.; Ismail, E.A.; Hassan, Z.K.; Al-Masoud, A.H.; Al-Dagal, M.M. Heat resistance and presence of genes encoding staphylococcal enterotoxins evaluated by multiplex-PCR of Staphylococcus aureus isolated from pasteurized camel milk. Biosci. Rep. 2019, 39, BSR20191225. [Google Scholar] [CrossRef]
- Yehia, H.M.; Al-Masoud, A.H.; Alarjani, K.M.; Alamri, M.S. Prevalence of methicillin-resistant (mecA gene) and heat-resistant Staphylococcus aureus strains in pasteurized camel milk. J. Dairy Sci. 2020, 103, 5947–5963. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| 60 °C Raw Milk | 60 °C Colostrum | |||||
|---|---|---|---|---|---|---|
| Time (min) | 10 | 20 | 30 | 10 | 20 | 30 |
| x/y * | 9/17 | 1/17 | 0/17 | 5/12 | 0/12 | 0/12 |
| 60 °C Raw Milk | 65 °C Raw Milk | 70 °C Raw Milk | |||
|---|---|---|---|---|---|
| Time (min) | 30 | 30 | 10 | 20 | 30 |
| x/y * | 2/2 | 2/2 | 12/17 | 3/17 | 0/17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wörmann, M.E.; Bhatte, A.; Wichmann-Schauer, H.; Tenhagen, B.-A.; Lienen, T. Heat Inactivation of Methicillin-Resistant Staphylococcus aureus Strains from German Dairy farms in Colostrum and Raw Milk. Animals 2023, 13, 3549. https://doi.org/10.3390/ani13223549
Wörmann ME, Bhatte A, Wichmann-Schauer H, Tenhagen B-A, Lienen T. Heat Inactivation of Methicillin-Resistant Staphylococcus aureus Strains from German Dairy farms in Colostrum and Raw Milk. Animals. 2023; 13(22):3549. https://doi.org/10.3390/ani13223549
Chicago/Turabian StyleWörmann, Mirka E., Ashwini Bhatte, Heidi Wichmann-Schauer, Bernd-Alois Tenhagen, and Tobias Lienen. 2023. "Heat Inactivation of Methicillin-Resistant Staphylococcus aureus Strains from German Dairy farms in Colostrum and Raw Milk" Animals 13, no. 22: 3549. https://doi.org/10.3390/ani13223549