Effect of Prepartum Maternal Supplementation with Diphenyl Diselenide on Biochemical, Immunological, and Oxidative Parameters of the Offspring

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethic Committee on Animal Use
2.2. Animals, Experimental Design, and Diet
2.3. Diphenyl Diselenide (PhSe)2
2.4. Execution Period and Location
2.5. Evaluation and Sample Collection
2.6. Blood Tests
2.7. Statistical Analysis
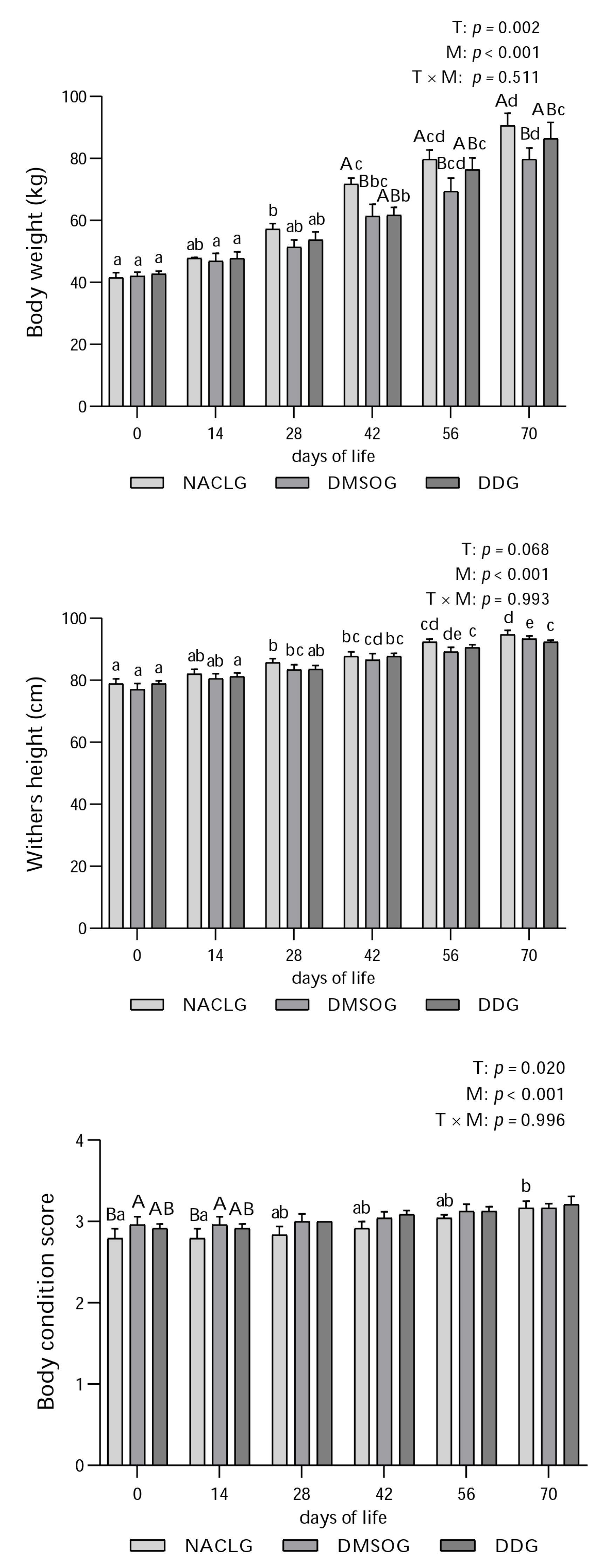
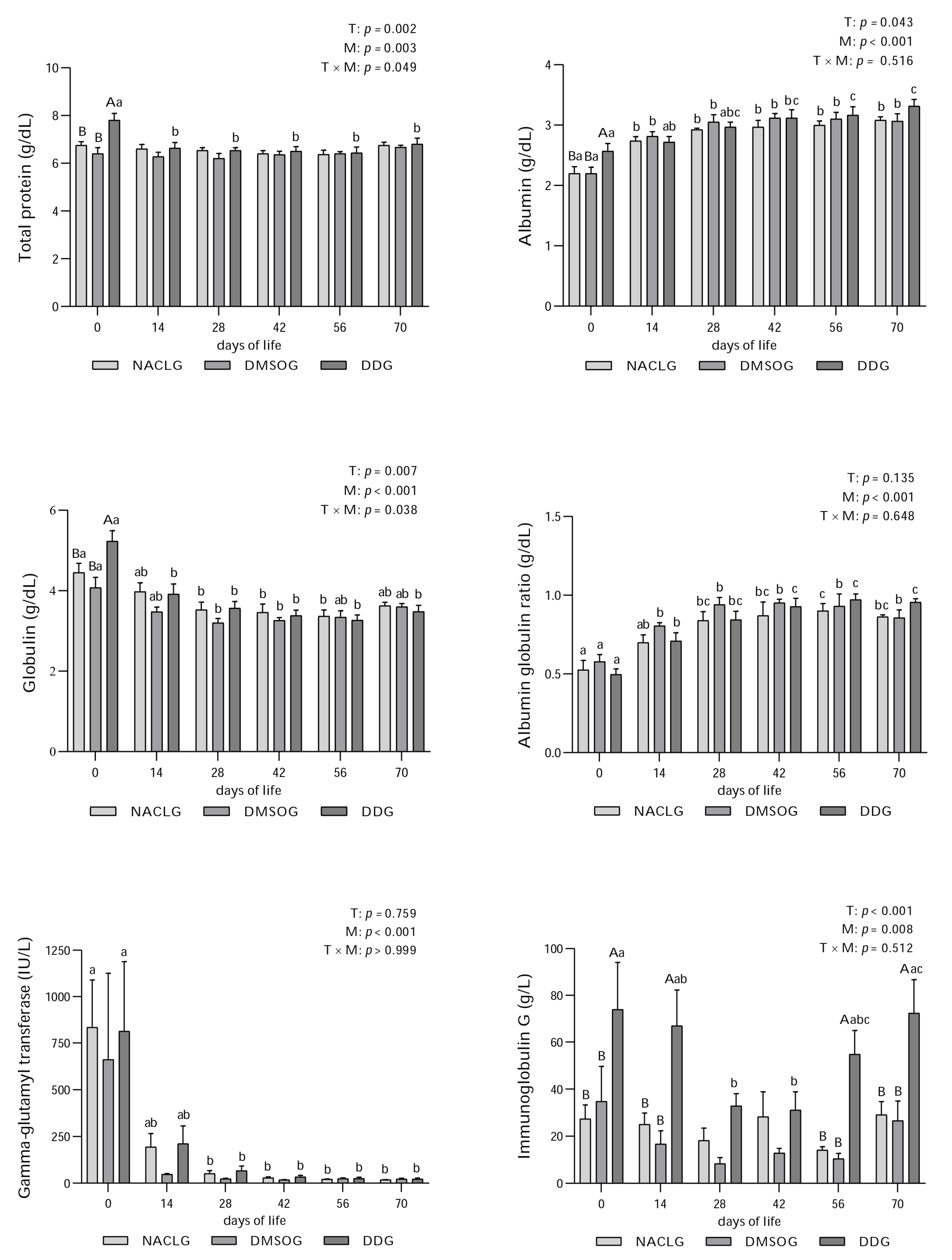
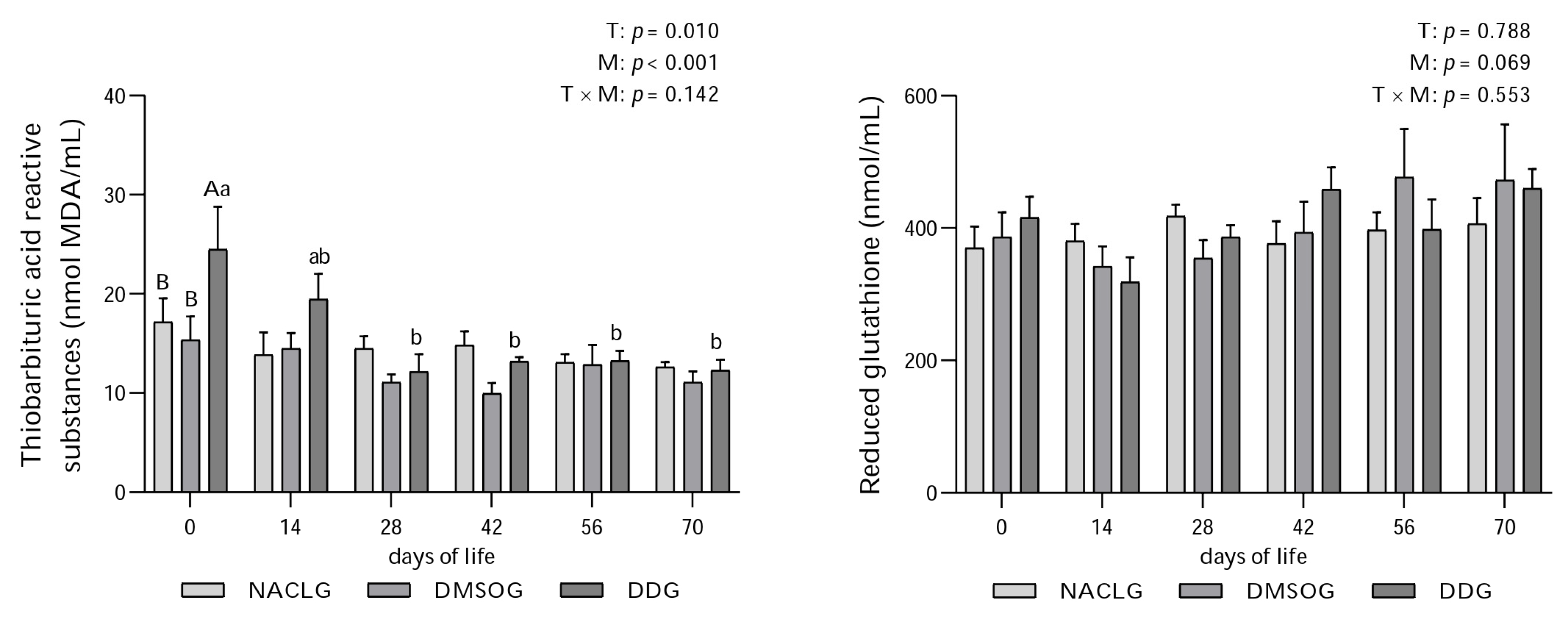
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vogels, Z.; Chuck, G.; Morton, J. Failure of transfer of passive immunity and agammaglobulinaemia in calves in south-west Victorian dairy herds: Prevalence and risk factors. Aust. Vet. J. 2013, 91, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum management for dairy calves. Vet. Clin. N. Am. 2019, 35, 535–556. [Google Scholar] [CrossRef] [PubMed]
- Beam, A.L.; Lombard, J.E.; Kopral, C.A.; Garber, L.P.; Winter, A.L.; Hicks, J.A.; Schlater, J.L. Prevalence of failure of passive transfer of immunity in newborn heifer calves and associated management practices on US dairy operations. J. Dairy Sci. 2009, 92, 3973–3980. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Hernández, J.; Benedito, J.; Castillo, C. Redox biology in transition periods of dairy cattle: Role in the health of periparturient and neonatal animals. Antioxidants 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Castillo, C.; Hernandez, J.; Bravo, A.; Lopez-Alonso, M.; Pereira, V.; Benedito, J.L. Oxidative status during late pregnancy and early lactation in dairy cows. Vet. J. 2005, 169, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, Ö.; Durak, İ. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Saugstad, O.D. Oxygen toxicity at birth: The pieces are put together. Pediatr. Res. 2003, 54, 789. [Google Scholar] [CrossRef]
- Inanami, O.; Shiga, A.; Okada, K.; Sato, R.; Miyake, Y.; Kuwabara, M. Lipid peroxides and antioxidants in serum of neonatal calves. Am. J. Vet. Res. 1999, 60, 452–457. [Google Scholar]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2010, 33, 233–240. [Google Scholar] [CrossRef]
- Rock, M.J.; Kincaid, R.L.; Carstens, G.E. Effects of prenatal source and level of dietary selenium on passive immunity and thermometabolism of newborn lambs. Small Rumin. Res. 2001, 40, 129–138. [Google Scholar] [CrossRef]
- Ghany-Hefnawy, A.E.; López-Arellano, R.; Revilla-Vázquez, A.; Ramírez-Bribiesca, E.; Tórtora-Pérez, J. The relationship between fetal and maternal selenium concentrations in sheep and goats. Small Rumin. Res. 2007, 73, 174–180. [Google Scholar] [CrossRef]
- Stewart, W.C.; Bobe, G.; Vorachek, W.R.; Stang, B.V.; Pirelli, G.J.; Mosher, W.D.; Hall, J.A. Organic and inorganic selenium: IV. Passive transfer of immunoglobulin from ewe to lamb. J. Anim. Sci. 2013, 91, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Schweizer, U.; Köhrle, J. Selenium and selenoproteins in mammals: Extraordinary, essential, enigmatic. Cell. Mol. Life Sci. 2004, 61, 1988–1995. [Google Scholar] [CrossRef] [PubMed]
- Apperson, K.D.; Vorachek, W.R.; Dolan, B.P.; Bobe, G.; Pirelli, G.J.; Hall, J.A. Effects of feeding pregnant beef cows selenium-enriched alfalfa hay on passive transfer of ovalbumin in their newborn calves. J. Trace Elem. Med. Biol. 2018, 50, 640–645. [Google Scholar] [CrossRef] [PubMed]
- McDowell, L.R. Feeding minerals to cattle on pasture. Anim. Feed Sci. Technol. 1996, 60, 247–271. [Google Scholar] [CrossRef]
- Prauchner, C.A. The Importance of Selenium for Agriculture and Human Health, 1st ed.; Publisher of the Federal University of Santa Maria: Santa Maria, RS, Brazil, 2020; pp. 256–261. [Google Scholar] [CrossRef]
- Leal, M.L.R.; Rocha, J.B.T.; Corte, C.L.D.; Aires, A.R.; Rocha, J.F.X.; Zanatta, R.S.; Carpes, J.L.S.; Szinwelski, G.A.; Stefanello, S.; Pivoto, F.L. Distribution of selenium in sheep treated with diphenyl diselenide. Arq. Bras. Med. Vet. Zootec. 2018, 70, 1017–1022. [Google Scholar] [CrossRef]
- Meotti, F.C.; Stangherlin, E.C.; Zeni, G.; Nogueira, C.W.; Rocha, J.B.T. Protective role of aryl and alkyl diselenides on lipid peroxidation. Environ. Res. 2004, 94, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Zoidis, E.; Chadio, S.E. Maternal selenium and developmental programming. Antioxidants 2019, 8, 145. [Google Scholar] [CrossRef]
- De Bem, A.F.; de Lima Portella, R.; Colpo, E.; Duarte, M.M.M.F.; Frediane, A.; Taube, P.S.; Nogueira, C.W.; Farina, M.; da Silva, E.L.; Teixeira Rocha, J.B. Diphenyl diselenide decreases serum levels of total cholesterol and tissue oxidative stress in cholesterol-fed rabbits. Basic Clin. Pharmacol. Toxicol. 2009, 105, 17–23. [Google Scholar] [CrossRef]
- De Bem, A.F.; Farina, M.; de Lima Portella, R.; Nogueira, C.W.; Dinis, T.C.P.; Laranjinha, J.A.N.; Almeida, L.M.; Rocha, J.B.T. Diphenyl diselenide, a simple glutathione peroxidase mimetic, inhibits human LDL oxidation in vitro. Atherosclerosis 2008, 201, 92–100. [Google Scholar] [CrossRef]
- Costa, M.D.; de Freitas, M.L.; Dalmolin, L.; Oliveira, L.P.; Fleck, M.A.; Pagliarini, P.; Acker, C.; Roman, S.S.; Brandão, R. Diphenyl diselenide prevents hepatic alterations induced by paraquat in rats. Environ. Toxicol. Pharmacol. 2013, 36, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Prauchner, C.A.; Prestes, A.D.S.; da Rocha, J.B.T. Effects of diphenyl diselenide on oxidative stress induced by sepsis in rats. Pathol. Res. Pract. 2011, 207, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Glaser, V.; Moritz, B.; Schmitz, A.; Dafré, A.L.; Nazari, E.M.; Rauh Müller, Y.M.; Feksa, L.; Straliottoa, M.R.; de Bem, A.F.; Farina, M.; et al. Protective effects of diphenyl diselenide in a mouse model of brain toxicity. Chem. Biol. Interact. 2013, 206, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Loreto, É.S.; Alves, S.H.; Santurio, J.M.; Nogueira, C.W.; Zeni, G. Diphenyl diselenide in vitro and in vivo activity against the oomycete Pythium insidiosum. Vet. Microbiol. 2012, 156, 222–226. [Google Scholar] [CrossRef]
- de Menezes, C.C.; Leitemperger, J.; Santi, A.; Lópes, T.; Aline Veiverberg, C.; Peixoto, S.; Bohrer Adaime, M.; Zanella, R.; Vargas Barbosa, N.B.; Lucia Loro, V. The effects of diphenyl diselenide on oxidative stress biomarkers in Cyprinus carpio exposed to herbicide quinclorac (Facet®). Ecotoxicol. Environ. Saf. 2012, 81, 91–97. [Google Scholar] [CrossRef]
- Roza, L.d.F.; da Silva, A.S.; Migliorini, M.J.; Bottari, N.B.; Glombowsky, P.; Baretta, M.; Tavernari, F.d.C.; Stefani, L.M.; Boiago, M. Adding diphenyl diselenide in the diets of quails improves the quality of meat. Rev. MVZ Cordoba 2018, 23, 6964–6973. [Google Scholar] [CrossRef]
- Stefanello, S.T.; da Rosa, E.J.F.; Dobrachinski, F.; Amaral, G.P.; de Carvalho, N.R.; da Luz, S.C.A.; Bender, C.R.; Schwab, R.S.; Dornelles, L.; Soares, F.A.A. Effect of diselenide administration in thioacetamide-induced acute neurological and hepatic failure in mice. Toxicol. Res. 2015, 4, 707–717. [Google Scholar] [CrossRef]
- Doleski, P.H.; Ten Caten, M.V.; Passos, D.F.; Castilhos, L.G.; Leal, D.B.R.; Machado, V.S.; Bottari, N.B.; Vogel, F.F.; Mendes, R.E.; da Silva, A.S. Toxoplasmosis treatment with diphenyl diselenide in infected mice modulates the activity of purinergic enzymes and reduces inflammation in spleen. Exp. Parasitol. 2017, 181, 7–13. [Google Scholar] [CrossRef]
- Dirksen, G.; Gründer, H.D.; Stöber, M. Clinical Examination of Cattle, 3rd ed.; Guanabara Koogan: Rio de Janeiro, RJ, Brazil, 1993; pp. 44–80. [Google Scholar]
- Reber, A.J.; Donovan, D.C.; Gabbard, J.; Galland, K.; Aceves-Avila, M.; Holbert, K.A.; Marshall, L.; Hurley, D.J. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system. Vet. Immunol. Immunopathol. 2008, 123, 305–313. [Google Scholar] [CrossRef]
- Ohkawa, K. Promotion of renewal canes in greenhouse roses by 6-benzylamino purine without cutback. HortScience 1979, 14, 612–613. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Sakia, R.M. The box-cox transformation technique: A review. Statistician 1992, 41, 169–178. [Google Scholar] [CrossRef]
- Gomes, V.; Madureira, K.M.; Borges, J.R.J.; Pinheiro, F.A.; Martin, C.C.; Baccili, C.C.; Santos, J.F.; Yasuoka, M.M.; Decaris, N.; Bombardelli, J.A.; et al. Pre-weaning diseases and calf health management practices. Rev. Bras. Buiat. 2021, 1, 27–62. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Eukaryotic selenoproteins and selenoproteomes. Biochim. Biophys. Acta 2009, 1790, 1424–1428. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef]
- Rooke, J.A.; Robinson, J.J.; Arthur, J.R. Effects of vitamin E and selenium on the performance and immune status of ewes and lambs. J. Agric. Sci. 2004, 142, 253–262. [Google Scholar] [CrossRef]
- Ganie, A.A.; Baghel, R.P.S.; Mudgal, V.; Shelkh, G.G. Effect of selenium supplementation on growth and nutrient utilization in buffalo heifers. Anim. Nutr. Feed Technol. 2010, 10, 255–259. Available online: https://www.researchgate.net/publication/326113102_Effect_of_Selenium_Supplementation_on_Growth_and_Nutrient_Utilization_in_Buffalo_Heifers (accessed on 10 October 2023).
- Rodrigues, C.M.; Schneider, M.; de Oliveira, J.S.; Collet, S.G.; Camillo, G.; Duarte, M.M.M.F.; Rocha, J.B.T.d.; Andrade, C.M.d.; Blagitz, M.G.; Leal, M.L.d.R. Does diphenyl diselenide metaphylaxis increase weight gain and immunoglobulin G in holstein calves from the neonatal period to weaning? Agric. Acad. J. 2020, 3, 49–61. [Google Scholar] [CrossRef]
- Santos, D.S.D.; Boito, J.P.; Klauck, V.; Reis, J.H.D.; Gebert, R.R.; Glombowsky, P.; Biazus, A.H.; Bottari, N.B.; Sousa, R.S.; Machado, G.; et al. Health benefits of subcutaneous zinc edetate and diphenyl diselenide in calves during the weaning period. An. Acad. Bras. Cienc. 2019, 91, 1–12. [Google Scholar] [CrossRef]
- Fagliari, J.J.; Santana, A.E.; Lucas, F.A.; Campos Filho, E.; Curi, P.R. Blood constituents of the newborn Nelore cattle (Bos indicus), Holstein cattle (Bos Taurus), and Murrah buffalo (Bubalus bubalis). Arq. Bras. Med. Vet. Zootec. 1998, 50, 253–262. Available online: https://repositorio.unesp.br/server/api/core/bitstreams/b06a7772-8418-4b55-819d-4cee67d1ac57/content (accessed on 15 October 2023).
- Pehrson, B.; Ortman, K.; Madjid, N.; Trafikowska, U. The influence of dietary selenium as selenium yeast or sodium selenite on the concentration of selenium in the milk of Suckler cows and on the selenium status of their calves. J. Anim. Sci. 1999, 77, 3371–3376. [Google Scholar] [CrossRef] [PubMed]
- Gunter, S.A.; Beck, P.A.; Phillips, J.M. Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 2003, 81, 856–864. [Google Scholar] [CrossRef]
- Guyot, H.; Spring, P.; Andrieu, S.; Rollin, F. Comparative responses to sodium selenite and organic selenium supplements in Belgian Blue cows and calves. Livest. Sci. 2007, 111, 259–263. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, S.; Yang, X.; Han, X.; Liu, Y.; Khan, N.A.; Tan, Z. Effects of organic and inorganic selenium on selenium bioavailability, growth performance, antioxidant status and meat quality of a local beef cattle in China. Front. Vet. Sci. 2023, 10, 1171751. [Google Scholar] [CrossRef]
- Hall, J.A.; Bobe, G.; Vorachek, W.R.; Estill, C.T.; Mosher, W.D.; Pirelli, G.J.; Gamroth, M. Effect of supranutritional maternal and colostral selenium supplementation on passive absorption of immunoglobulin G in selenium-replete dairy calves. J. Dairy Sci. 2014, 97, 4379–4391. [Google Scholar] [CrossRef]
- Kamada, H.; Nonaka, I.; Ueda, Y.; Murai, M. Selenium addition to colostrum increases immunoglobulin G absorption by newborn calves. J. Dairy Sci. 2007, 90, 5665–5670. [Google Scholar] [CrossRef]
- Golin, A.; Tinkov, A.A.; Aschner, M.; Farina, M.; da Rocha, J.B.T. Relationship between selenium status, selenoproteins and COVID-19 and other inflammatory diseases: A critical review. J. Trace Elem. Med. Biol. 2023, 75, 127099. [Google Scholar] [CrossRef]
- Ning, M.; Hua, S.; Ma, Y.; Liu, Y.; Wang, D.; Xu, K.; Yu, H. Microvesicles facilitate the differentiation of mesenchymal stem cells into pancreatic beta-like cells via miR-181a-5p/150-5p. Int. J. Biol. Macromol. 2023, 254, 127719. [Google Scholar] [CrossRef]
- Logan, E.F.; Penhale, W.J.; Jones, R.A. Changes in the serum immunoglobulin levels of colostrum-fed calves during the first 12 weeks postpartum. Res. Vet. Sci. 1973, 14, 394–397. [Google Scholar] [CrossRef]
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: San Diego, CA, USA, 2008; pp. 157–172. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhao, Q.; Zhan, T.; Han, Y.; Tang, C.; Zhang, J. Effect of different selenium sources on growth performance, tissue selenium content, meat quality, and selenoprotein gene expression in finishing pigs. Biol. Trace Elem. Res. 2019, 196, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Bami, M.K.; Afsharmanesh, M.; Espahbodi, M.; Esmaeilzadeh, E. Effects of dietary nano-selenium supplementation on broiler chicken performance, meat selenium content, intestinal microflora, intestinal morphology, and immune response. J. Trace Elem. Med. Biol. 2022, 69, 126897. [Google Scholar] [CrossRef]
- De Bem, A.F.; Fiuza, B.; Calcerrada, P.; Brito, P.M.; Peluffo, G.; Dinis, T.C.; Trujillo, M.; Rocha, J.B.T.; Radi, R.; Almeida, L.M. Protective effect of diphenyl diselenide against peroxynitrite-mediated endothelial cell death: A comparison with ebselen. Nitric Oxide 2013, 31, 20–30. [Google Scholar] [CrossRef]
- Dias, G.R.M.; Golombieski, R.M.; Portella, R.L.P.; Amaral, G.P.; Soares, F.A.; Rocha, J.B.T.; Nogueira, C.W.; Barbosa, N.V. Diphenyl diselenide modulates gene expression of antioxidant enzymes in the cerebral cortex, hippocampus and striatum of female hypothyroid rats. Neuroendocrinology 2014, 100, 45–59. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 8th ed.; National Academies Press: Washington, DC, USA, 2021; pp. 143–146. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredient | Values |
|---|---|
| Corn silage (% DM 2) | 64.07 |
| Ryegrass hay (% DM 2) | 12.00 |
| Soybean meal (% DM 2) | 11.23 |
| Soybean hulls (% DM 2) | 10.63 |
| Premix of vitamins and minerals 1 (% DM 2) | 2.07 |
| Chemical composition | |
| DM 2 (%) | 49.8 |
| CP 3 (% DM 2) | 11.3 |
| ADF 4 (% DM 2) | 26.5 |
| NDF 5 (% DM 2) | 42.6 |
| Starch (% DM 2) | 23.3 |
| Fatty acids (% DM 2) | 3.42 |
| Ca 6 (% DM 2) | 0.51 |
| P 7 (% DM 2) | 0.31 |
| ME 8 (Mcal/kg) | 2.57 |
| NEL 9 (Mcal/kg) | 1.70 |
| Se 10 (mg/kg) | 0.41 |
| Parameters | Group | Values |
|---|---|---|
| Brix index (%) | NACLG 1 | 24.87 ± 1.21 |
| DMSOG 2 | 24.62 ± 0.80 | |
| DDG 3 | 25.50 ± 1.54 | |
| IgG 4 (g/L) | NACLG 1 | 111.67 ± 11.18 |
| DMSOG 2 | 106.70 ± 12.83 | |
| DDG 3 | 113.83 ± 12.29 |
| Ingredient | Quantities/kg * |
|---|---|
| Vitamin A (IU/kg) | 74,806 |
| Vitamin D3 (IU/kg) | 21,210 |
| Vitamin E (IU/kg) | 1090 |
| Calcium (g/kg) | 175.2 |
| Chlorine (g/kg) | 24.6 |
| Crude protein (g/kg) | 119.5 |
| Ethereal extract (g/kg) | 54.2 |
| Lactose (g/kg) | 70.5 |
| Sodium (g/kg) | 16.5 |
| Biotin (mg/kg) | 25 |
| Chromium (mg/kg) | 8.3 |
| Cobalt (mg/kg) | 4.4 |
| Cooper (mg/kg) | 354.8 |
| Fluorine (mg/kg) | 295 |
| Iodine (mg/kg) | 15 |
| Magnesium (mg/kg) | 1500 |
| Manganese (mg/kg) | 1068.2 |
| Monensin (mg/kg) | 700 |
| Phosphorus (mg/kg) | 8000 |
| Potassium (mg/kg) | 2500 |
| Selenium (mg/kg) | 6.5 |
| Sulfur (mg/kg) | 300 |
| Zinc (mg/kg) | 1022.1 |
| Saccharomyces cerevisiae (CFU/kg) | 1.0 × 1011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, C.M.; Parmeggiani, E.B.; Leal, K.W.; Schneider, M.; Collet, S.G.; Cibin, F.W.S.; Gomes, V.; Blagitz, M.G.; Rocha, J.B.T.d.; Leal, M.L.d.R. Effect of Prepartum Maternal Supplementation with Diphenyl Diselenide on Biochemical, Immunological, and Oxidative Parameters of the Offspring. Animals 2024, 14, 10. https://doi.org/10.3390/ani14010010
Rodrigues CM, Parmeggiani EB, Leal KW, Schneider M, Collet SG, Cibin FWS, Gomes V, Blagitz MG, Rocha JBTd, Leal MLdR. Effect of Prepartum Maternal Supplementation with Diphenyl Diselenide on Biochemical, Immunological, and Oxidative Parameters of the Offspring. Animals. 2024; 14(1):10. https://doi.org/10.3390/ani14010010
Chicago/Turabian StyleRodrigues, Cláudia Medeiros, Eliana Burtet Parmeggiani, Karoline Wagner Leal, Marla Schneider, Silvana Giacomini Collet, Francielli Weber Santos Cibin, Viviani Gomes, Maiara Garcia Blagitz, João Batista Teixeira da Rocha, and Marta Lizandra do Rêgo Leal. 2024. "Effect of Prepartum Maternal Supplementation with Diphenyl Diselenide on Biochemical, Immunological, and Oxidative Parameters of the Offspring" Animals 14, no. 1: 10. https://doi.org/10.3390/ani14010010