
Effects of Essential Oil and/or Encapsulated Butyrate on Fecal Microflora in Neonatal Holstein Calves

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Materials
2.1.1. Test Animals
2.1.2. Feeding Procedure
2.2. Main Reagents and Instruments
2.3. Sample Collection
2.4. Sample DNA Extraction
2.5. PCR Amplification and Sequencing Library Construction
2.6. High-Throughput Sequencing Data Analysis
2.7. Volatile Fatty Acid Detection
2.8. Statistical Analysis
3. Results
3.1. Sequencing Results and Diversity Analysis
3.1.1. Basic Sequencing Results
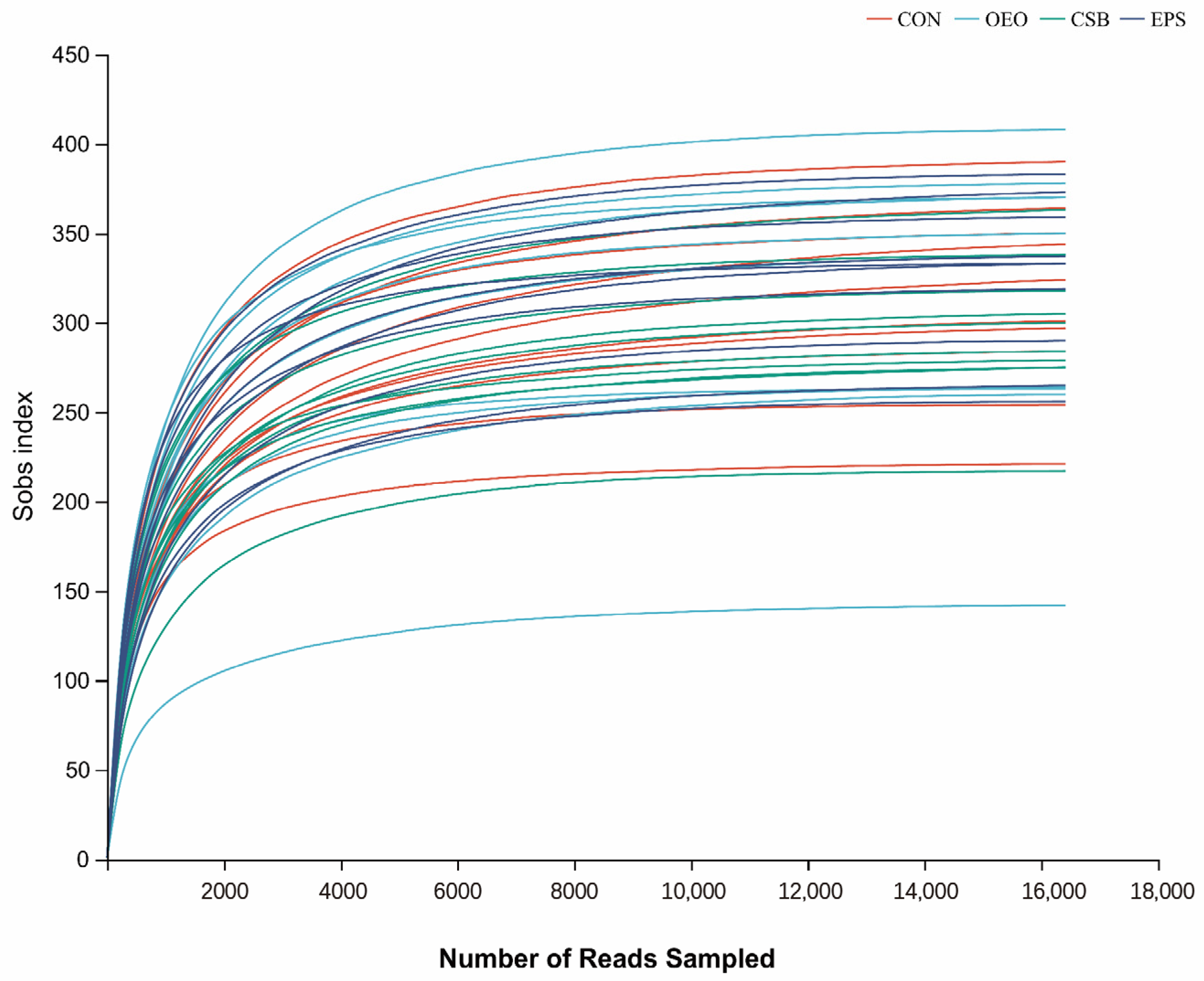
3.1.2. Dilution Curve
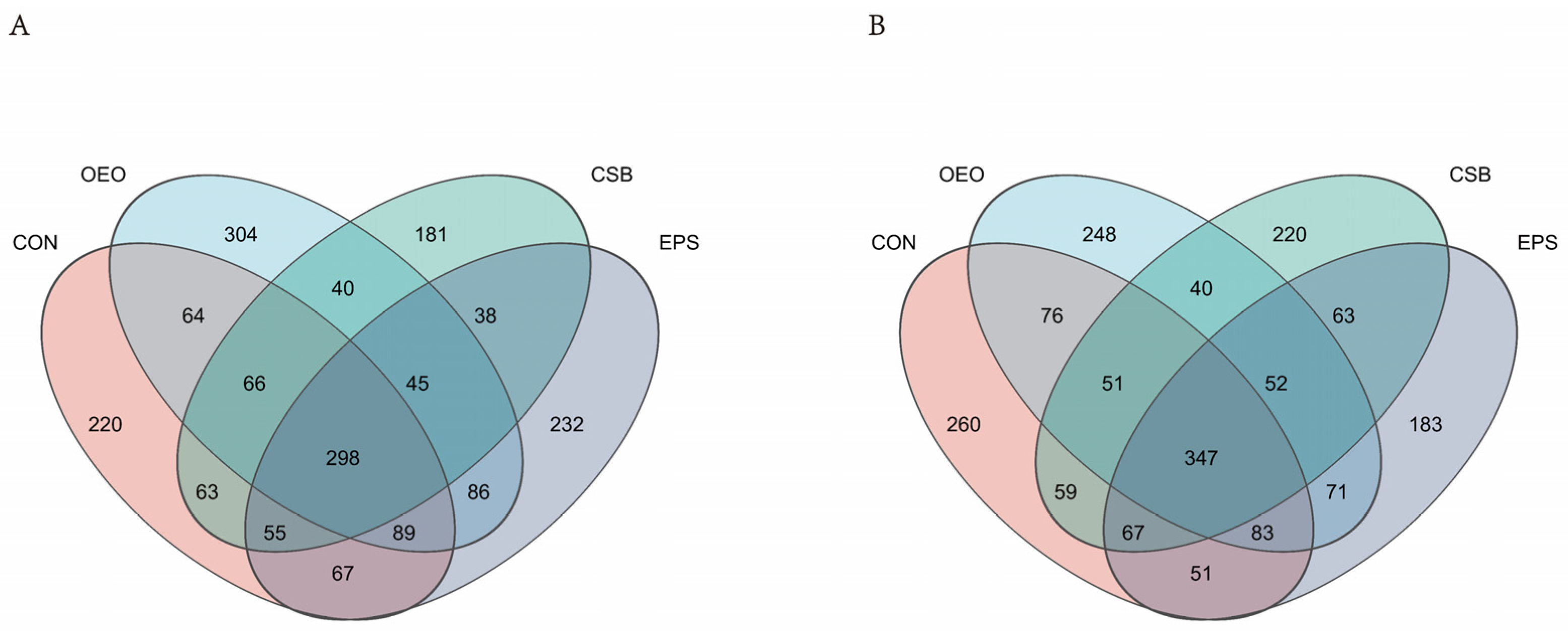
3.1.3. Analysis of Species Composition
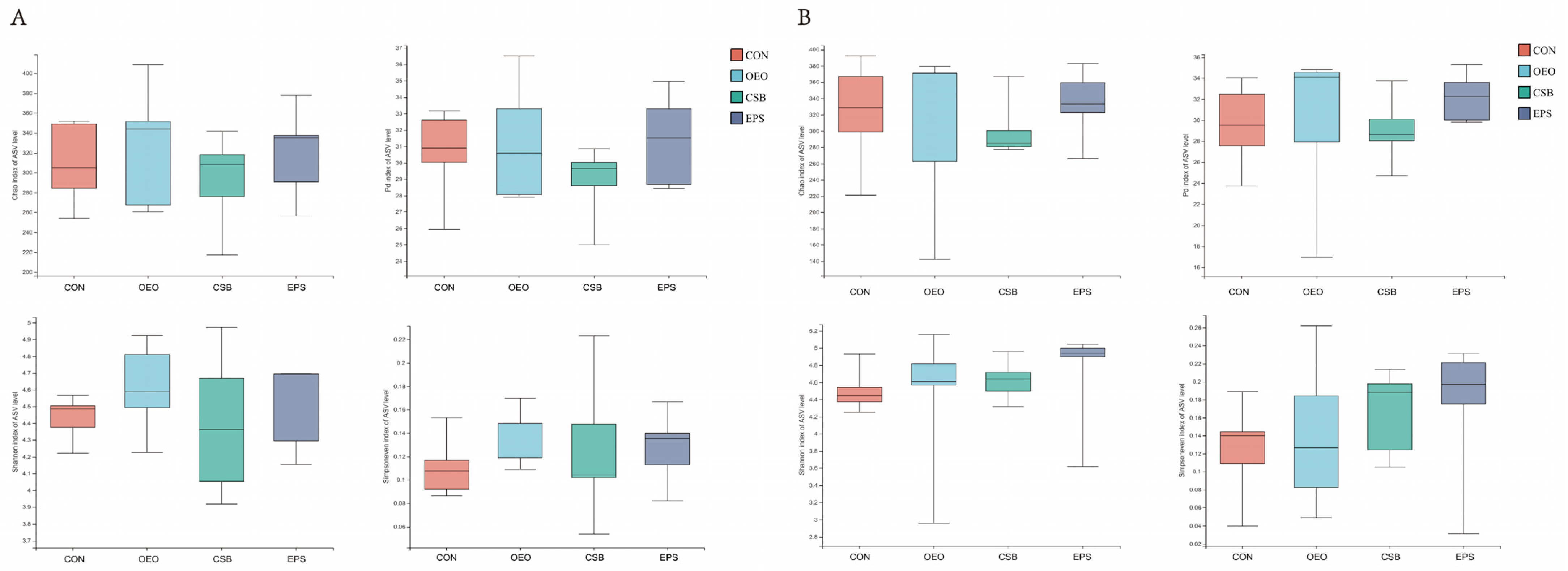
3.1.4. Alpha Diversity Analysis
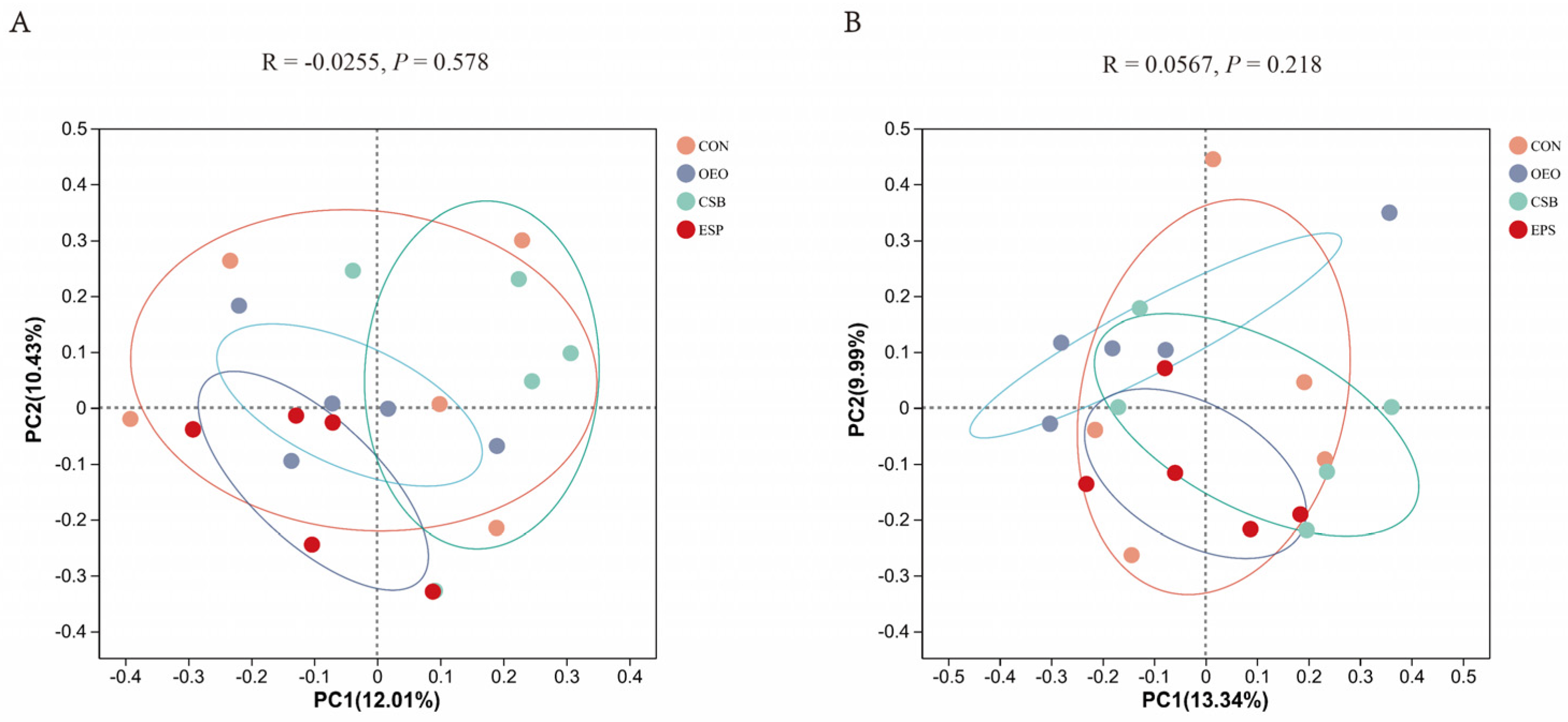
3.1.5. Beta Diversity Analysis
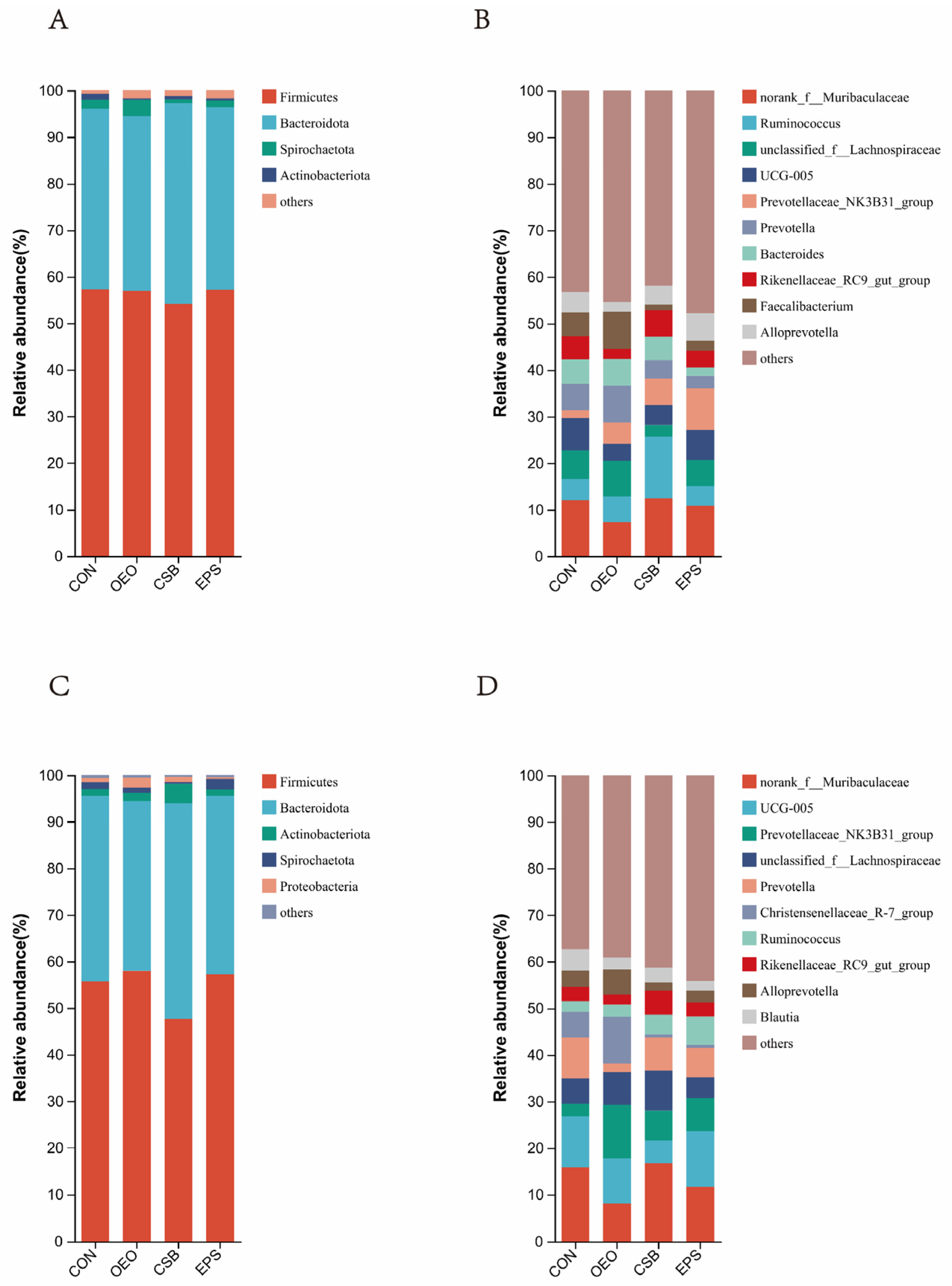
3.2. Species Composition Analysis at the Phylum and Genus Level
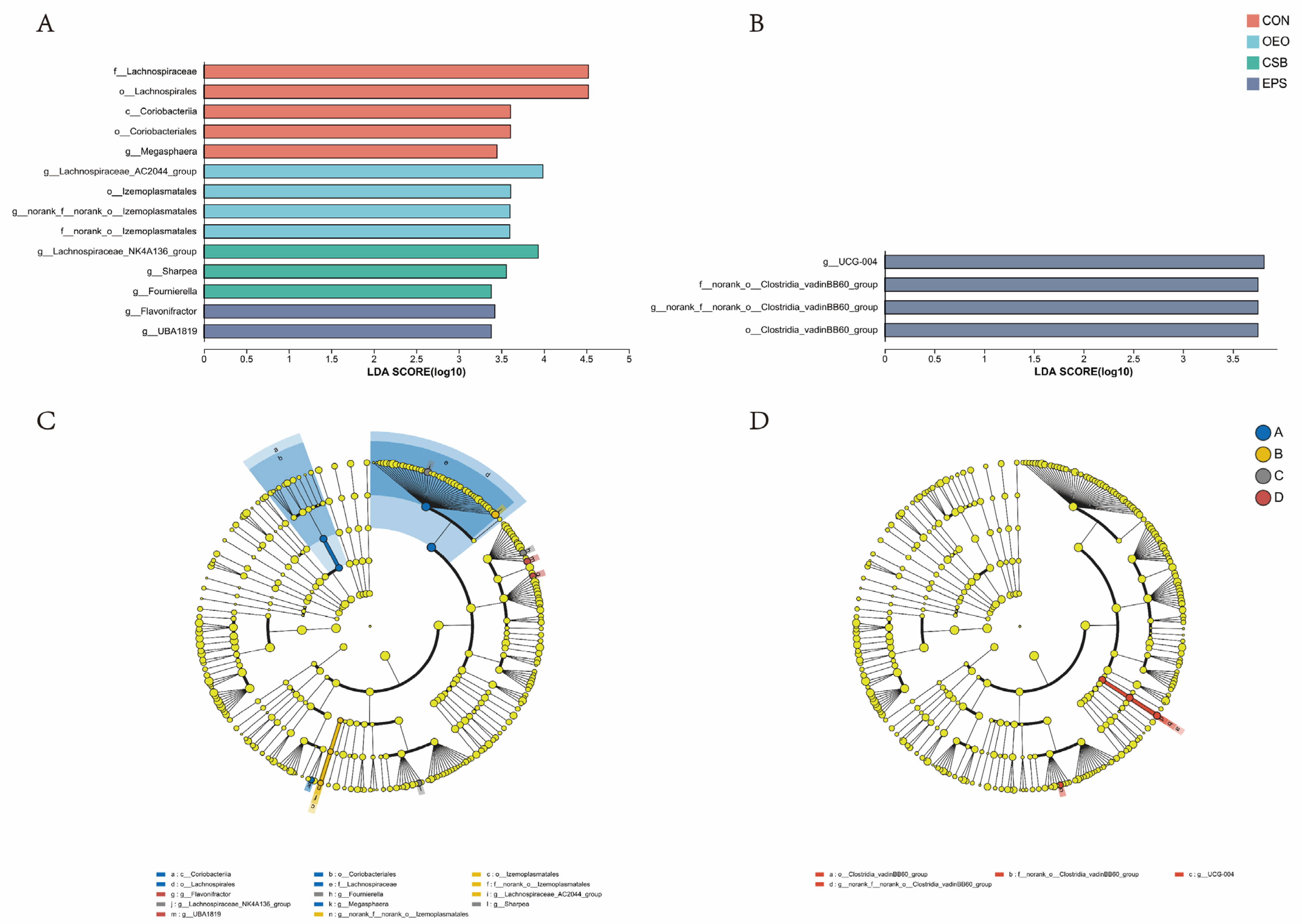
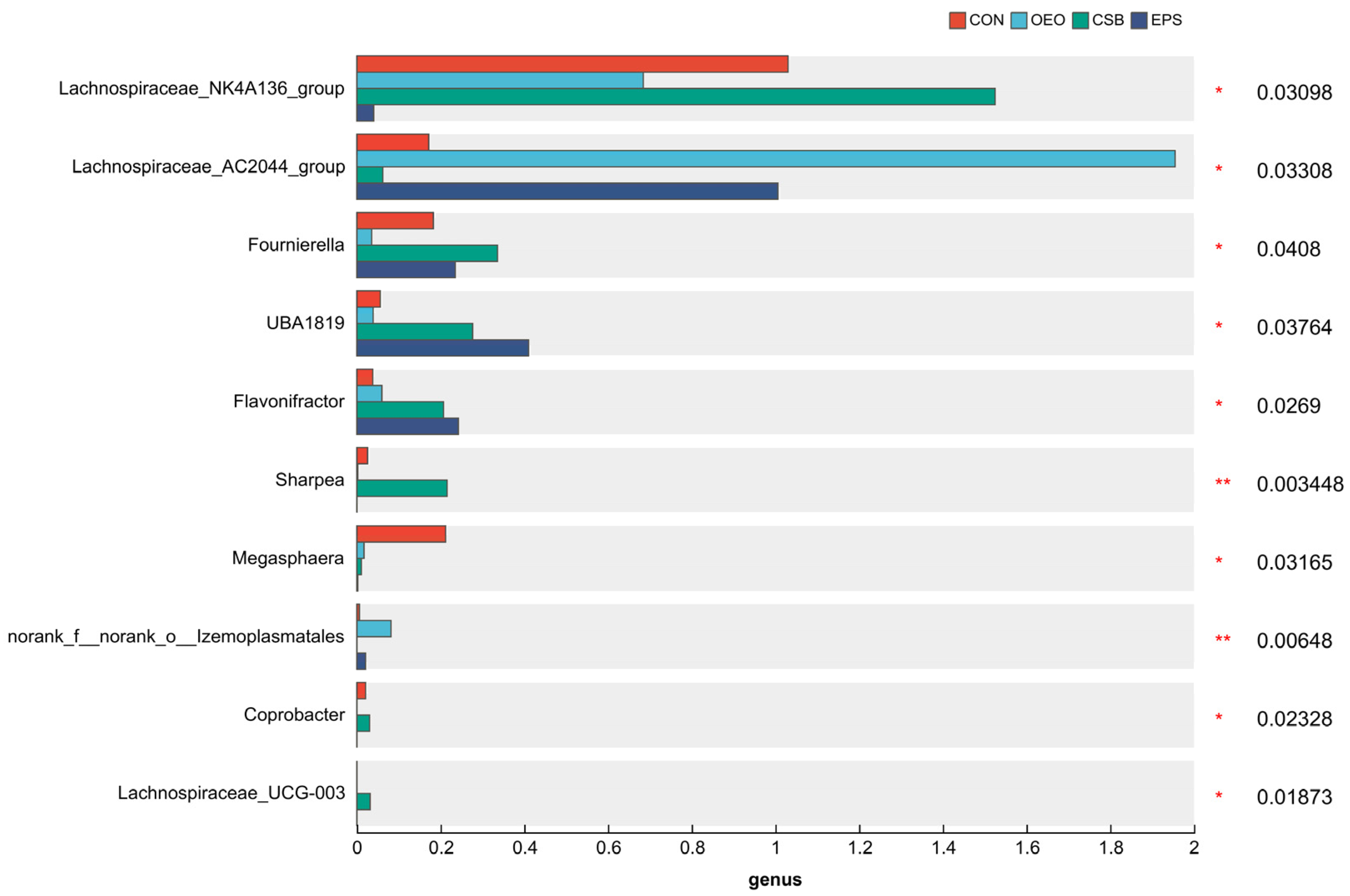
3.3. Species Difference Analysis
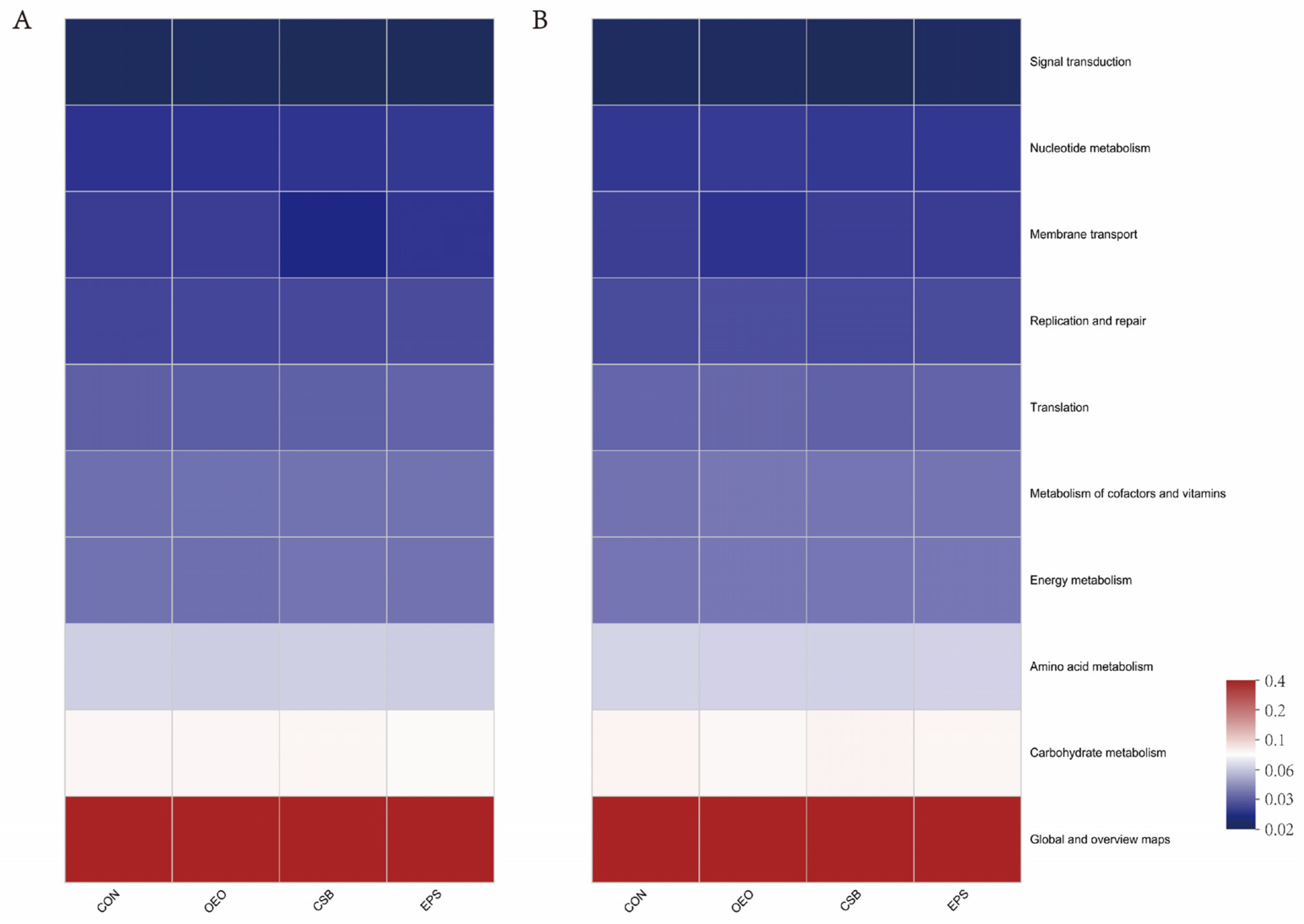
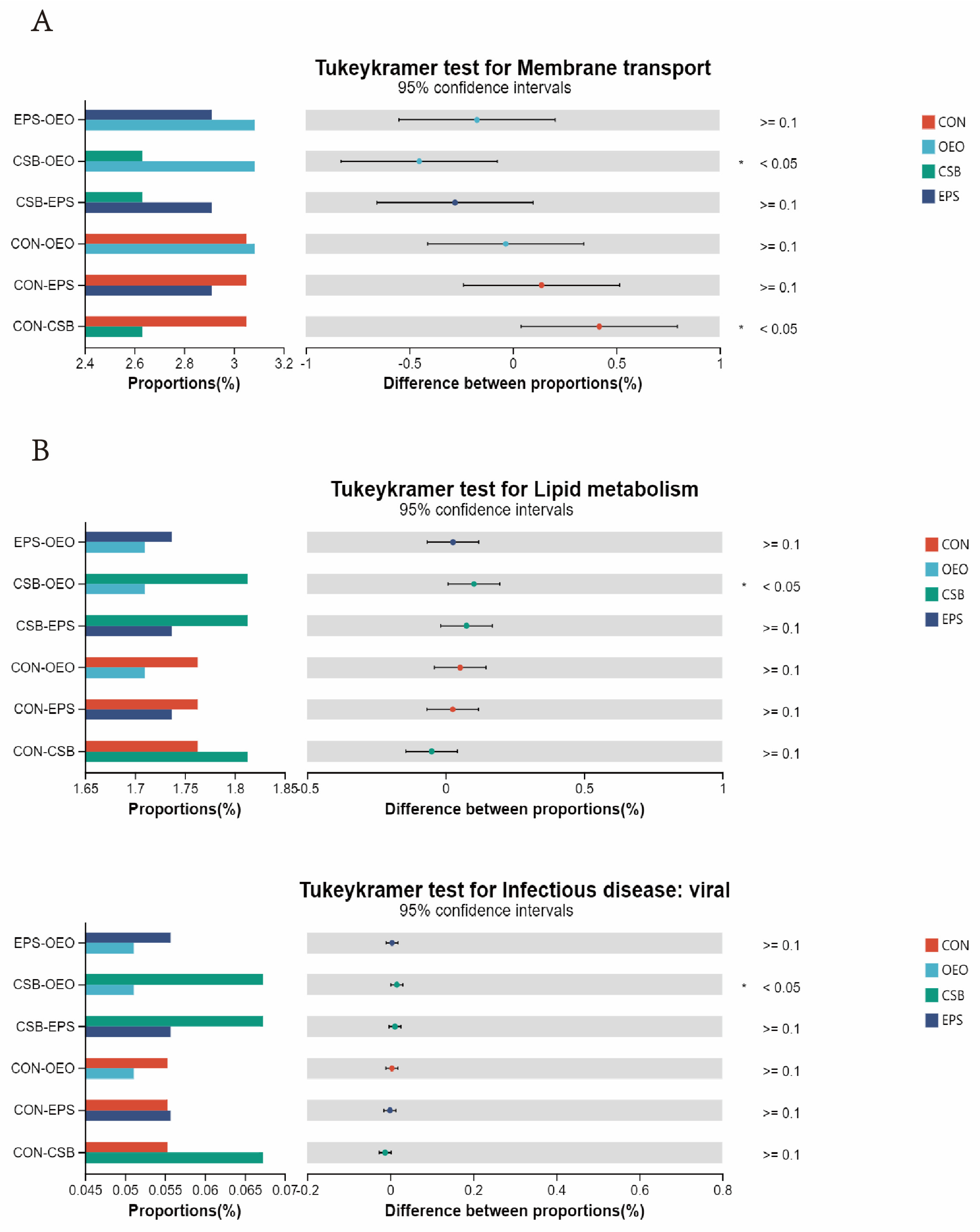
3.4. Microbial Function Prediction Analysis
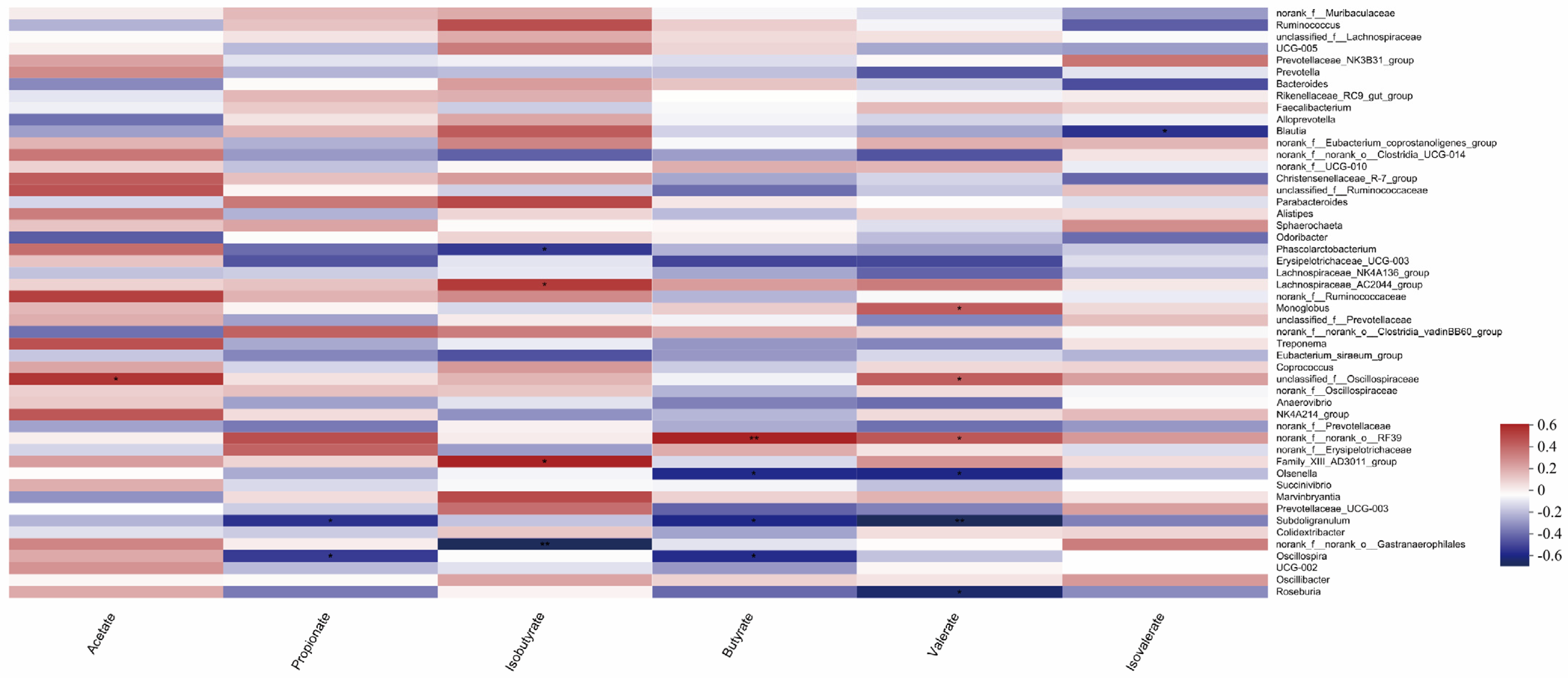
3.5. Correlation Analysis of Gut Microbiome and Production of Volatile Fatty Acids in Calves
4. Discussion
4.1. Effects of Oregano Essential Oil, Sodium Butyrate and Their Mixtures on Intestinal Microbial Diversity of Calves
4.2. Effects of Oregano Essential Oil, Sodium Butyrate, and Their Mixtures on Intestinal Microbial Species Composition of Calves
4.3. Effects of Oregano Essential Oil, Sodium Butyrate, and Their Mixtures in Species Differences of Intestinal Microbiome in Calves
4.4. Effects of Oregano Essential Oil, Sodium Butyrate, and Their Mixtures on Intestinal Microbes of Calves
4.5. Duration of Influence of Treatment Group on Microbial Structure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naeem, A.; Drackley, J.K.; Stamey, J.; Loor, J.J. Role of metabolic and cellular proliferation genes in ruminal development in response to enhanced plane of nutrition in neonatal Holstein calves. J. Dairy Sci. 2012, 95, 1807–1820. [Google Scholar] [CrossRef] [PubMed]
- Fonty, G.; Gouet, P.; Nebout, J.M. Development of the cellulolytic microflora in the rumen of lambs transferred into sterile isolators a few days after birth. Can. J. Microbiol. 1989, 35, 416–422. [Google Scholar] [CrossRef]
- Conroy, M.E.; Shi, H.N.; Walker, W.A. The long-term health effects of neonatal microbial flora. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; et al. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.H.; Nielsen, D.S.; Kverka, M.; Zakostelska, Z.; Klimesova, K.; Hudcovic, T.; Tlaskalova-Hogenova, H.; Hansen, A.K. Patterns of early gut colonization shape future immune responses of the host. PLoS ONE 2012, 7, e34043. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G. Microbiology of the Rumen. In Rumenology; Springer: Cham, Switzerland, 2016; Chapter 2; pp. 39–61. [Google Scholar] [CrossRef]
- Sun, W.; Shi, H.; Gong, C.; Liu, K.; Li, G. Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth. Microorganisms 2023, 11, 1444. [Google Scholar] [CrossRef]
- Uyeno, Y.; Chao, W.; Jayanegara, A.; Kondo, M.; Ban-Tokuda, T.; Matsui, H. Increase in Rumen Fibrolytic Bacteria and the Improvement of Fiber Degradability of Ensiled Total Mixed Ration Assessed by in vitro Rumen Culture. Adv. Anim. Vet. Sci. 2016, 4, 183–186. [Google Scholar] [CrossRef]
- Wu, H.M.; Zhang, J.; Wang, C.; Liu, Q.; Guo, G.; Huo, W.J.; Chen, L.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L. Effects of riboflavin supplementation on performance, nutrient digestion, rumen microbiota composition and activities of Holstein bulls. Br. J. Nutr. 2021, 126, 1288–1295. [Google Scholar] [CrossRef]
- Du, Y.; Gao, Y.; Hu, M.; Hou, J.; Yang, L.; Wang, X.; Du, W.; Liu, J.; Xu, Q. Colonization and development of the gut microbiome in calves. J. Anim. Sci. Biotechnol. 2023, 14, 46. [Google Scholar] [CrossRef]
- Zhang, Y.; Choi, S.H.; Nogoy, K.M.; Liang, S. Review: The development of the gastrointestinal tract microbiota and intervention in neonatal ruminants. Animal 2021, 15, 100316. [Google Scholar] [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C.; et al. A Recent Insight Regarding the Phytochemistry and Bioactivity of Origanum vulgare L. Essential Oil. Int. J. Mol. Sci. 2020, 21, 9653. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Di Stefano, A.; Cacciatore, I. Carvacrol and its derivatives as antibacterial agents. Phytochem 2018, 17, 903–921. [Google Scholar] [CrossRef]
- Lin, B.; Lu, Y.; Wang, J.H.; Liang, Q.; Liu, J.X. The effects of combined essential oils along with fumarate on rumen fermentation and methane production in vitro. J. Anim. Feed Sci. 2012, 21, 198–210. [Google Scholar] [CrossRef]
- Khorrami, B.; Vakili, A.R.; Mesgaran, M.D.; Klevenhusen, F. Thyme and cinnamon essential oils: Potential alrnatives for monensin as a rumen modifier in beef production systems. Anim. Feed Sci. Technol. 2015, 200, 8–16. [Google Scholar] [CrossRef]
- Mullen, K.A.; Lee, A.R.; Lyman, R.L.; Mason, S.E.; Washburn, S.P.; Anderson, K.L. An in vitro assessment of the antibacterial activity of plant-derived oils. J. Dairy Sci. 2014, 97, 5587–5591. [Google Scholar] [CrossRef]
- Miguel, M.; Lee, S.S.; Mamuad, L.; Choi, Y.J.; Jeong, C.D.; Son, A.; Cho, K.K.; Kim, E.T.; Kim, S.B.; Lee, S.S. Enhancing Butyrate Production, Ruminal Fermentation and Microbial Population through Supplementation with Clostridium saccharobutylicum. J. Microbiol. Biotechnol. 2019, 29, 1083–1095. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Penner, G.B.; Stumpff, F.; Gäbel, G. Ruminant Nutrition Symposium: Role of fermentation acid absorption in the regulation of ruminal pH. J. Anim. Sci. 2011, 89, 1092–1107. [Google Scholar] [CrossRef]
- Ahsan, U.; Cengiz, Ö.; Raza, I.; Kuter, E.; Chacher, M.F.A.; Iqbal, Z.; Umar, S.; Çakir, S. Sodium butyrate in chicken nutrition: The dynamics of performance, gut microbiota, gut morphology, and immunity. World Poultry Sci. J. 2016, 72, 265–275. [Google Scholar] [CrossRef]
- Guilloteau, P. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef]
- Sun, Q.; Ji, Y.C.; Wang, Z.L.; She, X.; He, Y.; Ai, Q.; Li, L.Q. Sodium Butyrate Alleviates Intestinal Inflammation in Mice with Necrotizing Enterocolitis. Mediators Inflamm. 2021, 2021, 6259381. [Google Scholar] [CrossRef]
- Karaca, N.; Şener, G.; Demirci, B.; Demirci, F. Synergistic antibacterial combination of Lavandula latifolia Medik. essential oil with camphor. Z. Naturforsch C J. Biosci. 2020, 76, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhao, D.; Ma, W.; Guo, Y.; Wang, A.; Wang, Q.; Lee, D.J. Denitrifying sulfide removal process on high-salinity wastewaters in the presence of Halomonas sp. Appl. Microbiol. Biotechnol. 2016, 100, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Caporaso, J.G.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef]
- Van der Waaij, D.; Berghuis-de Vries, J.M.; Lekkerkerk-Van der Wees, J.E.C. Colonization resistance of the digestive tract in conventional and antibiotic-treated mice. J. Hyg. 1971, 69, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD—What role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Yu, Z.; Wang, B. Perilla frutescens Leaf Alters the Rumen Microbial Community of Lactating Dairy Cows. Microorganisms 2019, 7, 562. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.H.; Ahmed, M.S.; Aboubakr, M.; Ghoneim, H.A.; Abdel-Daim, M.M.; Albadrani, G.M.; Arafat, N.; Fadl, S.E.; Abdo, W. Impact of Dietary or Drinking Water Ruminococcus sp. Supplementation and/or Heat Stress on Growth, Histopathology, and Bursal Gene Expression of Broilers. Front. Vet. Sci. 2021, 8, 663577. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Ye, W.; Zu, X.; Xie, H.; Li, H.; Li, Y.; Zhang, W. Integrative metabolic and microbial profiling on patients with Spleen-yang-deficiency syndrome. Sci. Rep. 2018, 8, 6619. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, F.; Zou, P.; Li, X. Bacillus amyloliquefaciens SC06 in the diet improves egg quality of hens by altering intestinal microbiota and the effect is diminished by antimicrobial peptide. Front. Nutr. 2022, 9, 999998. [Google Scholar] [CrossRef]
- Huang, P.; Jiang, A.; Wang, X.; Zhou, Y.; Tang, W.; Ren, C.; Qian, X.; Zhou, Z.; Gong, A. NMN Maintains Intestinal Homeostasis by Regulating the Gut Microbiota. Front. Nutr. 2021, 8, 714604. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, J.; Cai, Z.; Zhou, K.; Chang, L.; Bai, Y.; Ma, Y. Prevotella Induces the Production of Th17 Cells in the Colon of Mice. J. Immunol. Res. 2020, 2020, 9607328. [Google Scholar] [CrossRef]
- Wu, W.; Lv, L.; Shi, D.; Ye, J.; Fang, D.; Guo, F.; Li, Y.; He, X.; Li, L. Protective Effect of Akkermansia muciniphila against Immune-Mediated Liver Injury in a Mouse Model. Front. Microbiol. 2017, 8, 1804. [Google Scholar] [CrossRef]
- Ahir, V.B.; Koringa, P.G.; Bhatt, V.D.; Ramani, U.V.; Joshi, C.G. Metagenomic analysis of poultry gut microbes. J. Poult. Sci. 2010, 45, 111–114. [Google Scholar]
- Cheng, J.; Zhang, X.; Xu, D.; Zhang, D.; Zhang, Y.; Song, Q.; Li, X.; Zhao, Y.; Zhao, L.; Li, W.; et al. Relationship between rumen microbial differences and traits among Hu sheep, Tan sheep, and Dorper sheep. J. Anim. Sci. 2022, 100, skac261. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, Z.G.; Pei, C.F.; Allan, D.; Hao, L.Z.; Cao, X.L.; Liu, H.S.; Zhou, J.Y.; Long, R.J. A comparison between yaks and Qaidam cattle in in vitro rumen fermentation, methane emission, and bacterial community composition with poor quality substrate. Anim. Feed Sci. Technol. 2022, 291, 115395. [Google Scholar] [CrossRef]
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.M.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 6, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bindels, L.B.; Segura Munoz, R.R.; Martínez, I.; Walter, J.; Ramer-Tait, A.E.; Rose, D.J. Disparate Metabolic Responses in Mice Fed a High-Fat Diet Supplemented with Maize-Derived Non-Digestible Feruloylated Oligo- and Polysaccharides Are Linked to Changes in the Gut Microbiota. PLoS ONE 2016, 11, e0146144. [Google Scholar] [CrossRef]
- Ahmad, A.A.; Zhang, J.; Liang, Z.; Du, M.; Yang, Y.; Zheng, J.; Yan, P.; Long, R.; Tong, B.; Han, J.; et al. Age-dependent variations in rumen bacterial community of Mongolian cattle from weaning to adulthood. BMC Microbiol. 2022, 22, 213. [Google Scholar] [CrossRef]
- Liao, R.; Xie, X.; Lv, Y.; Dai, J.; Lin, Y.; Zhu, L. Ages of weaning influence the gut microbiota diversity and function in Chongming white goats. Appl. Microbiol. Biotechnol. 2021, 105, 3649–3658. [Google Scholar] [CrossRef]
- Hao, Y.; Gong, Y.; Huang, S.; Ji, S.; Wang, W.; Wang, Y.; Yang, H.; Cao, Z.; Li, S. Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle. Microorganisms 2021, 9, 1788. [Google Scholar] [CrossRef]
- Zhong, Y.B.; Kang, Z.P.; Wang, M.X.; Long, J.W.; Hai, Y.; Huang, J.Q.; Wei, S.Y.; Zhou, W.; Zhao, H.M.; Liu, D.Y. Curcumin ameliorated dextran sulfate sodium-induced colitis via regulating the homeostasis of DCs and Treg and improving the composition of the gut microbiota. J. Funct. Foods 2021, 86, 104716. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, L.; Li, H.; Xia, F.; Bai, H.; Piao, X.; Sun, Z.; Cui, H.; Shi, L. Dietary Oregano Essential Oil Supplementation Influences Production Performance and Gut Microbiota in Late-Phase Laying Hens Fed Wheat-Based Diets. Animals 2022, 12, 3007. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, G.; Zhang, S.; Zhao, J. Fructooligosaccharide Reduces Weanling Pig Diarrhea in Conjunction with Improving Intestinal Antioxidase Activity and Tight Junction Protein Expression. Nutrients. 2022, 14, 512. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, Y.; Huang, K.; Yang, B.; Zhang, Y.; Yu, Z.; Wang, J. Fecal microbiota dynamics and its relationship to diarrhea and health in dairy calves. J. Anim. Sci. Biotechnol. 2022, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Sun, S.; Luo, Z.; Shi, B.; Shan, A.; Cheng, B. Maternal dietary resveratrol alleviates weaning-associated diarrhea and intestinal inflammation in pig offspring by changing intestinal gene expression and microbiota. Food Funct. 2019, 10, 5626–5643. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wu, J.; Lang, X.; Liu, L.; Casper, D.P.; Wang, C.; Zhang, L.; Wei, S. Effects of oregano essential oil on in vitro ruminal fermentation, methane production, and ruminal microbial community. J. Dairy Sci. 2020, 103, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.W.; Radovanovic, A.; Willson, N.L.; Bajagai, Y.S.; Hao Van, T.T.; Moore, R.J.; Stanley, D. Oregano: A potential prophylactic treatment for the intestinal microbiota. Heliyon 2019, 5, e02625. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, J.; Zhang, W.; Nie, C. Impacts of sodium butyrate on intestinal mucosal barrier and intestinal microbial community in a weaned piglet model. Front. Microbiol. 2023, 13, 1041885. [Google Scholar] [CrossRef]
- Yu, D.Y.; Oh, S.H.; Kim, I.S.; Kim, G.I.; Kim, J.A.; Moon, Y.S.; Jang, J.C.; Lee, S.S.; Jung, J.H.; Park, H.C.; et al. Effects of lactic acid bacteria fermented feed and three types of lactic acid bacteria (L. plantarum, L. acidophilus, B. animalis) on intestinal microbiota and T cell polarization (Th1, Th2, Th17, Treg) in the intestinal lymph nodes and spleens of rats. Anim. Biosci. 2023, 36, 156–166. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Acidified Milk |
|---|---|
| Milk fat (%) | 2.81 |
| Milk protein (%) | 3.29 |
| SNF (%) | 9.58 |
| Lactose (%) | 4.88 |
| Ash (%) | 0.74 |
| TDM (%) | 11.86 |
| Proportion | 31.63 |
| pH | 4.13 |
| Conductivity (mS/cm) | 7.72 |
| Freezing Point (°C) | −0.558 |
| Ingredients | Content | Nutrient Level | Content |
|---|---|---|---|
| Corn | 40.54 | DM | 87.95 |
| Soybean meal | 32.00 | CP | 22.17 |
| Wheat bran | 5.80 | EE | 3.79 |
| Cottonseed meal | 5.30 | Ash | 5.91 |
| Puffed soybeans | 5.00 | ADF | 6.18 |
| Whey powder | 4.00 | NDF | 12.23 |
| Molasses | 4.00 | Ca | 0.91 |
| CaCO3 | 1.60 | P | 0.59 |
| Soybean oil | 0.80 | ||
| NaCl | 0.60 | ||
| CaHPO4 | 0.10 | ||
| MgO | 0.10 | ||
| Selenium yeast | 0.02 | ||
| Premix | 0.14 | ||
| Total | 100.00 |
| Index | CON | OEO | CSB | EPS |
|---|---|---|---|---|
| Before weaning | ||||
| Firmicutes | 57.29 | 56.97 | 54.17 | 57.19 |
| Bacteroidota | 38.77 | 37.49 | 43.06 | 39.24 |
| Spriochatetota | 1.93 | 3.56 | 0.86 | 1.38 |
| Actinobacteriota | 1.30 | 1.74 | 1.26 | 0.50 |
| Firmicutes/Bacteroidota (F/B) | 1.48 | 1.52 | 1.26 | 1.46 |
| After weaning | ||||
| Firmicutes | 55.64 | 57.99 | 47.63 | 57.19 |
| Bacteroidota | 39.93 | 36.43 | 46.32 | 38.39 |
| Actinobacteriota | 1.10 | 1.84 | 4.17 | 1.33 |
| Spriochatetota | 1.49 | 1.03 | 0.37 | 2.30 |
| Proteobacteria | 0.88 | 2.10 | 1.14 | 0.34 |
| Firmicutes/Bacteroidota (F/B) | 1.39 | 1.59 | 1.03 | 1.49 |
| Index | CON | OEO | CSB | EPS |
|---|---|---|---|---|
| Before weaning | ||||
| norank_f__Muribaculaceae | 12.05 | 7.27 | 12.49 | 10.82 |
| Ruminococcus | 4.59 | 5.57 | 13.16 | 4.25 |
| unclassified_f__Lachnospiraceae | 6.06 | 7.66 | 2.50 | 5.62 |
| UCG-005 | 6.92 | 3.61 | 4.31 | 6.41 |
| Prevotellaceae_NK3B31_group | 1.70 | 4.63 | 5.63 | 8.98 |
| Prevotella | 5.69 | 7.87 | 3.97 | 2.66 |
| Bacteroides | 5.25 | 5.74 | 5.08 | 1.81 |
| Rikenellaceae_RC9_gut_group | 5.02 | 2.12 | 5.71 | 3.55 |
| Faecalibacterium | 5.09 | 7.93 | 1.16 | 2.25 |
| Alloprevotella | 4.40 | 2.15 | 4.05 | 5.79 |
| After weaning | ||||
| norank_f__Muribaculaceae | 15.85 | 8.02 | 16.65 | 11.66 |
| UCG-005 | 10.93 | 9.70 | 4.97 | 11.90 |
| Prevotellaceae_NK3B31_group | 2.74 | 11.50 | 6.37 | 7.08 |
| unclassified_f__Lachnospiraceae | 5.34 | 7.00 | 8.63 | 4.48 |
| Prevotella | 8.81 | 1.91 | 7.12 | 6.39 |
| Christensenellaceae_R7_group | 5.55 | 10.01 | 0.63 | 0.60 |
| Ruminococcus | 2.29 | 2.61 | 4.24 | 6.13 |
| Rikenellaceae_RC9_gut_group | 3.04 | 2.15 | 5.19 | 2.99 |
| Alloprevotella | 3.53 | 5.42 | 1.78 | 2.54 |
| Blautia | 4.59 | 2.58 | 3.07 | 2.05 |
| Index | Before Weaning | After Weaning | ||||||
|---|---|---|---|---|---|---|---|---|
| CON | OEO | CSB | EPS | CON | OEO | CSB | EPS | |
| Global and overview maps | 40.83 | 40.71 | 40.99 | 40.60 | 40.76 | 40.64 | 40.83 | 40.69 |
| Carbohydrate metabolism | 9.84 | 9.87 | 9.99 | 9.67 | 9.81 | 9.66 | 9.92 | 9.80 |
| Amino acid metabolism | 7.16 | 7.05 | 7.15 | 7.09 | 7.22 | 7.15 | 7.12 | 7.13 |
| Energy metabolism | 4.25 | 4.20 | 4.29 | 4.27 | 4.22 | 4.28 | 4.25 | 4.26 |
| Metabolism of cofactors and vitamin | 4.16 | 4.22 | 4.25 | 4.24 | 4.18 | 4.31 | 4.22 | 4.19 |
| Translation | 3.77 | 3.74 | 3.83 | 3.90 | 3.85 | 3.90 | 3.76 | 3.81 |
| Replication and repair | 3.23 | 3.24 | 3.29 | 3.35 | 3.21 | 3.32 | 3.23 | 3.27 |
| Membrane transport | 3.07 | 3.09 | 2.26 | 2.91 | 3.03 | 2.78 | 3.02 | 2.99 |
| Nucleotide metabolism | 2.87 | 2.85 | 2.89 | 2.96 | 2.88 | 2.93 | 2.88 | 2.87 |
| Signal transduction | 2.17 | 2.19 | 2.13 | 2.16 | 2.17 | 2.17 | 2.07 | 2.20 |
| Index | CON | OEO | CSB | EPS | SEM | p-Value |
|---|---|---|---|---|---|---|
| Acetic acid | 355.40 ± 60.46 b | 462.95 ± 72.29 a | 418.36 ± 86.84 a | 453.05 ± 83.73 a | 11.248 | 0.002 |
| Propanoic acid | 147.60 ± 51.01 ab | 172.43 ± 52.47 ab | 189.46 ± 57.49 a | 135.82 ± 62.02 b | 7.662 | 0.054 |
| Isobutyric acid | 2.49 ± 0.78 | 3.13 ± 1.56 | 2.43 ± 0.95 | 2.74 ± 0.82 | 0.143 | 0.286 |
| Butyric acid | 153.93 ± 70.56 | 241.90 ± 287.57 | 139.35 ± 83.89 | 122.87 ± 83.48 | 21.560 | 0.201 |
| Valeric acid | 3.14 ± 1.02 b | 4.99 ± 2.60 a | 4.86 ± 1.64 a | 4.60 ± 1.92 a | 0.261 | 0.044 |
| Isovaleric acid | 2.95 ± 1.35 | 3.97 ± 1.14 | 3.80 ± 2.42 | 3.94 ± 1.71 | 0.228 | 0.363 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Z.; Liu, T.; Li, P.; Cheng, S.; Casper, D.P. Effects of Essential Oil and/or Encapsulated Butyrate on Fecal Microflora in Neonatal Holstein Calves. Animals 2023, 13, 3523. https://doi.org/10.3390/ani13223523
Luo Z, Liu T, Li P, Cheng S, Casper DP. Effects of Essential Oil and/or Encapsulated Butyrate on Fecal Microflora in Neonatal Holstein Calves. Animals. 2023; 13(22):3523. https://doi.org/10.3390/ani13223523
Chicago/Turabian StyleLuo, Zhihao, Ting Liu, Peng Li, Shuru Cheng, and David P. Casper. 2023. "Effects of Essential Oil and/or Encapsulated Butyrate on Fecal Microflora in Neonatal Holstein Calves" Animals 13, no. 22: 3523. https://doi.org/10.3390/ani13223523