
Histological Characteristics of Conjunctiva-Associated Lymphoid Tissue in Young and Adult Holstein Cattle
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
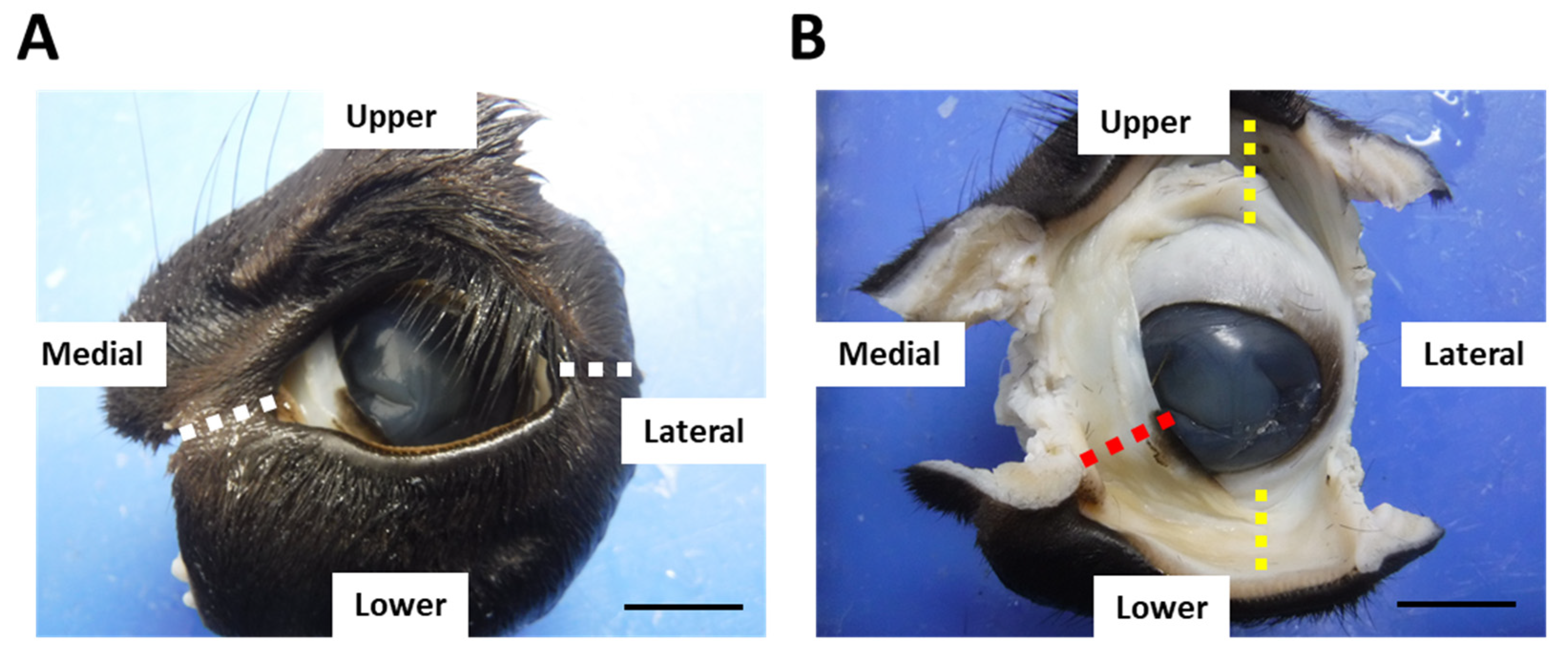
2.1. Animals and Sample Preparation
2.2. Histological Examination
2.3. Histoplanimetry
2.4. Statistical Analysis
3. Results
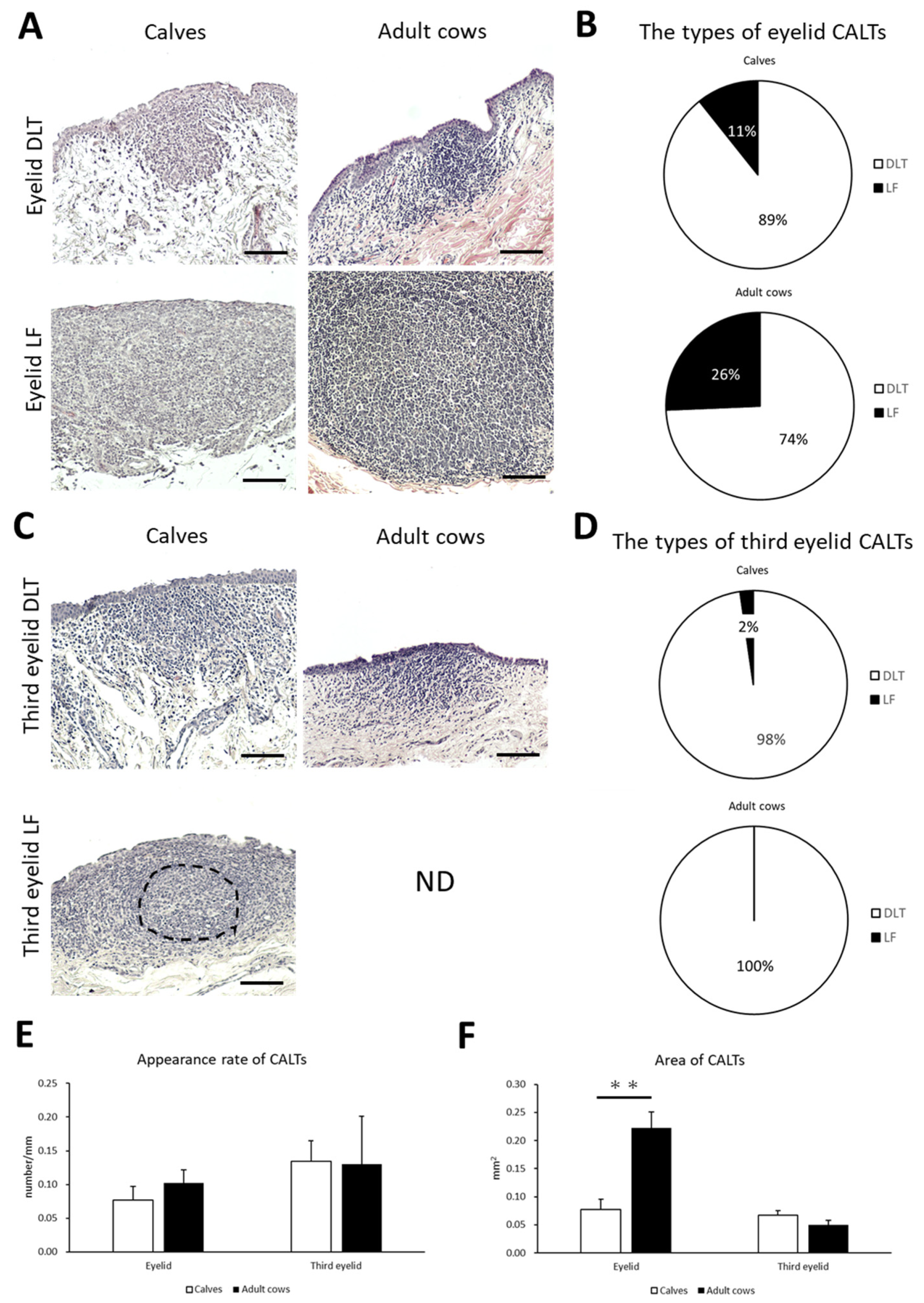
3.1. Histological Structure of CALT in Calves and Adult Cows
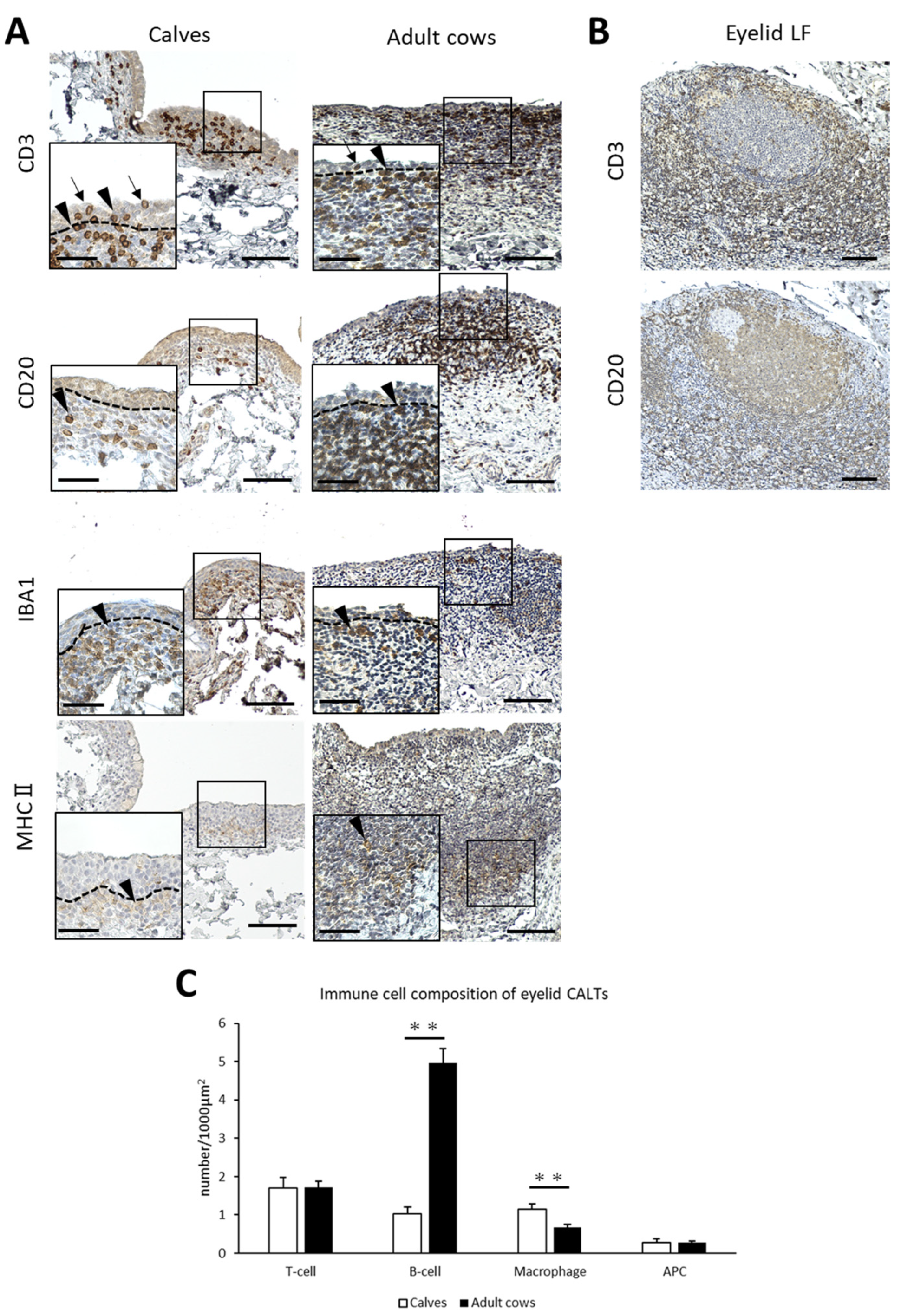
3.2. Immune Cell Composition of Eyelid CALT in Calves and Adult Cows
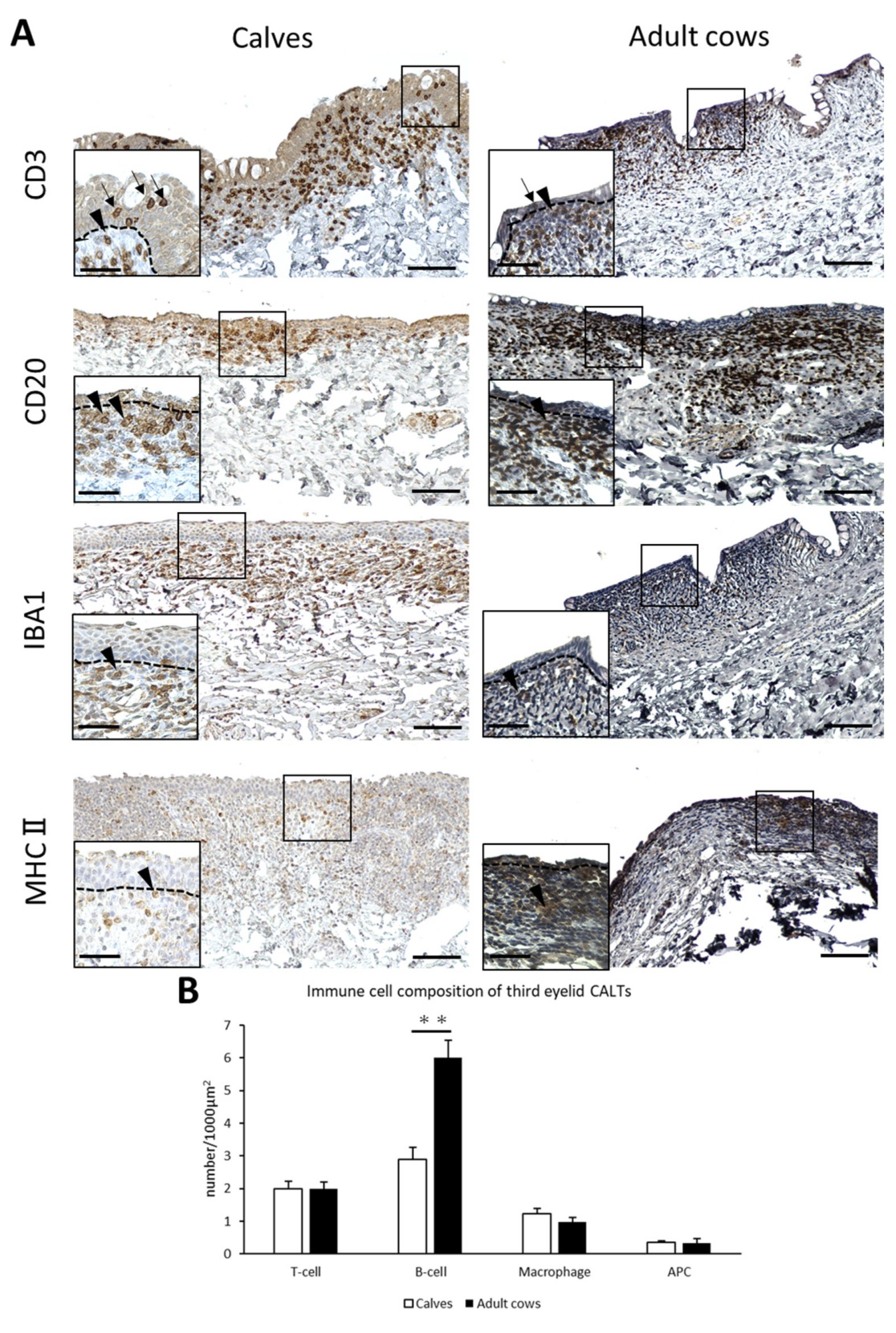
3.3. Immune Cell Composition of Third Eyelid CALT in Calves and Adult Cows
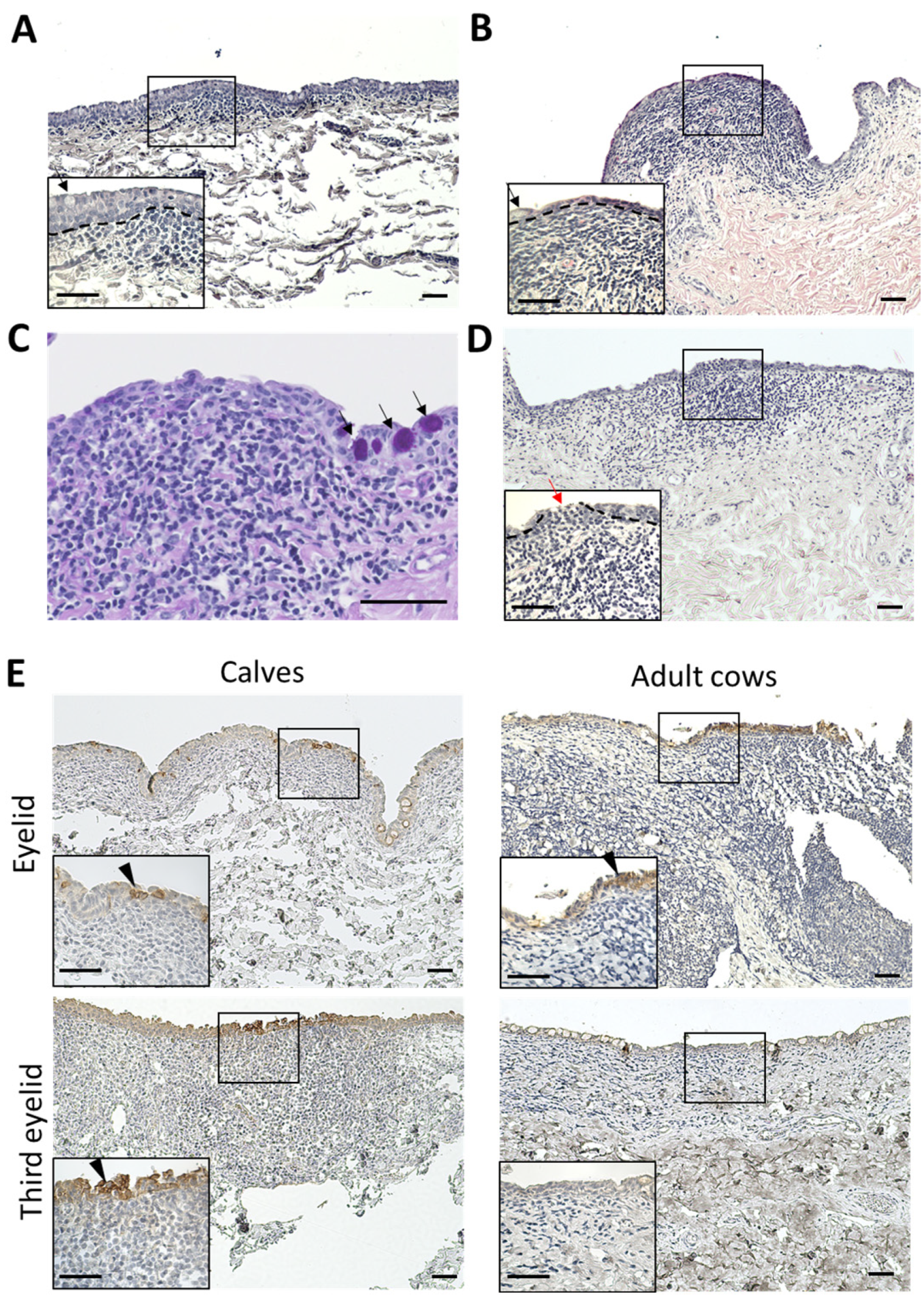
3.4. Histological Characteristics and CK18 Expression of Epithelial Layer of CALT
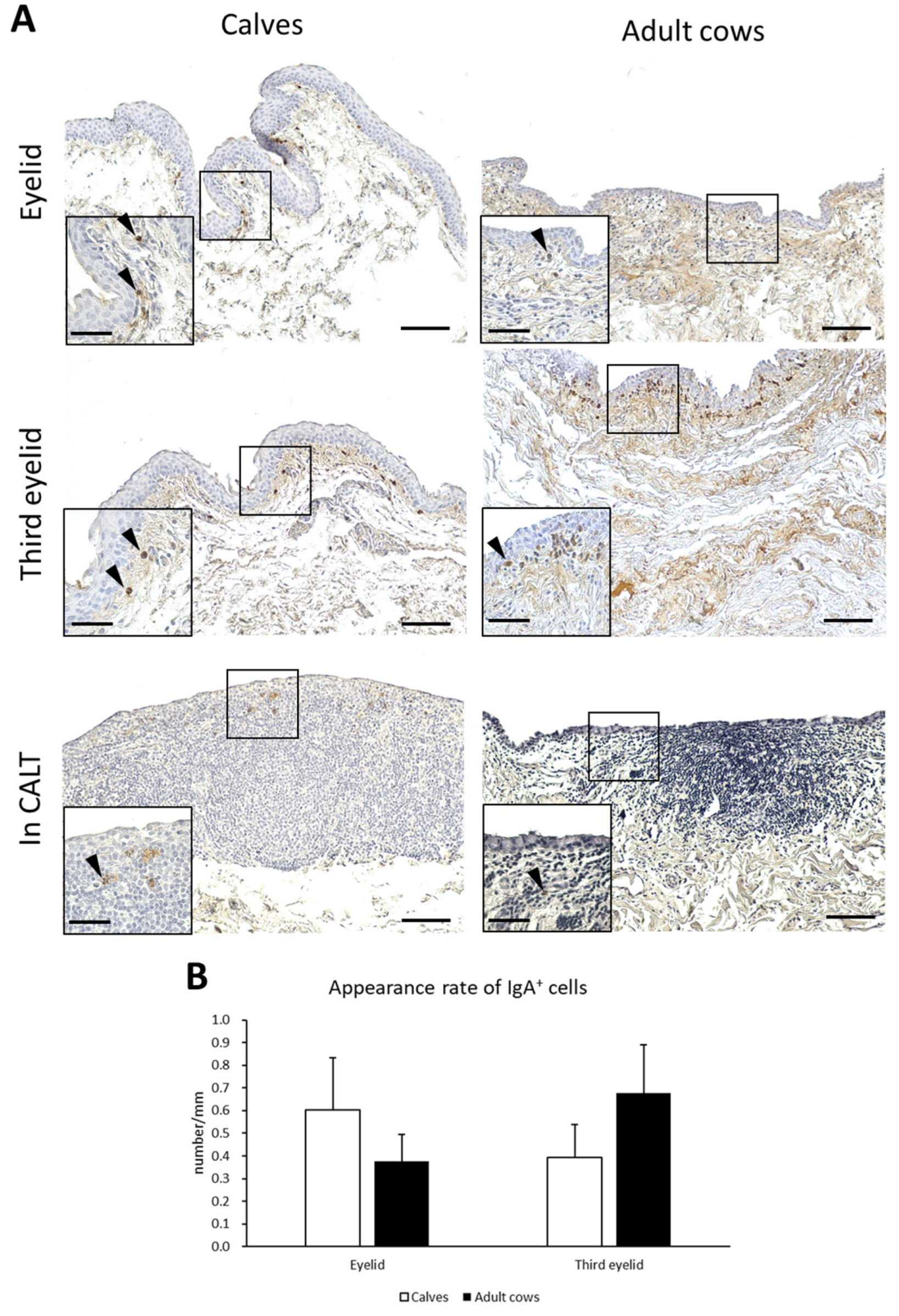
3.5. IgA+ Cells in Conjunctiva of Calves and Adult Cows
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liebler-Tenorio, E.M.; Pabst, R. MALT structure and function in farm animals. Vet. Res. 2006, 37, 257–280. [Google Scholar] [CrossRef] [PubMed]
- Kiyono, H.; Fukuyama, S. NALT-versus Peyer’s-patch-mediated mucosal immunity. Nat. Rev. Immunol. 2004, 4, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Forehielli, M.L.; Walker, W.A. The role of gut-associated lymphoid tissues and mucosal defence. Br. J. Nutr. 2009, 93, S41–S48. [Google Scholar] [CrossRef]
- Spit, B.J.; Hendriksen, E.G.; Bruijntjes, J.P.; Kuper, C.F. Nasal lymphoid tissue in the rat. Cell Tissue Res. 1989, 255, 193–198. [Google Scholar] [CrossRef]
- Pabst, R.; Gehrke, I. Is the bronchus-associated lymphoid tissue (BALT) an integral structure of the lung in normal mammals, including humans? Am. J. Respir. Cell Mol. Biol. 1990, 3, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Belz, G.T.; Heath, T.J. The epithelium of canine palatine tonsils. Anat. Embryol. 1995, 192, 189–194. [Google Scholar] [CrossRef]
- Bockman, D.E.; Cooper, M.D. Pinocytosis by epithelium associated with lymphoid follicles in the bursa of Fabricius, appendix and Peyer’s patches. An electron microscopic study. Am. J. Anat. 1973, 136, 455–478. [Google Scholar] [CrossRef]
- Owen, R.L.; Bye, W.A. The membranous (M) cell and the mucosal immune system. Ann. Rev. Med. 1984, 35, 95–112. [Google Scholar]
- Owen, R.L.; Jones, A.L. Epithelial cell specialization within human Peyer's patches: An ultrastructural study of intestinal lymphoid follicles. Gastroenterology 1974, 66, 189–203. [Google Scholar] [CrossRef]
- Bye, W.A.; Allan, C.H.; Trier, J.S. Structure, distribution, and origin of M cells in Peyer’s patches of mouse ileum. Gastroenterology 1984, 86, 789–801. [Google Scholar]
- Neutra, M.R. M cells in antigen sampling in mucosal tissues. Curr. Top. Microbiol. Immunol. 1999, 236, 17–32. [Google Scholar]
- Owen, R.L. Sequential uptake of horseradish peroxidase by lymphoid follicle epithelium of Peyer’s patches in the normal unobstructed mouse intestine: An ultrastructural study. Gastroenterology 1977, 72, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Cesta, M.F. Normal structure, function, and histology of mucosa-associated lymphoid tissue. Toxicol. Pathol. 2006, 34, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Chuluunbaatar, T.; Ichii, O.; Masum, M.A.; Namba, T.; Islam, M.R.; Otani, Y.; Elewa, Y.H.A.; Kon, Y. Genital organ-associated lymphoid tissues arranged in a ring in the mucosa of cow vaginal vestibules. Res. Vet. Sci. 2022, 145, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Osman, R.; Malmuthuge, N.; Gonzalez-Cano, P.; Griebel, P. Development and function of the mucosal immune system in the upper respiratory tract of neonatal calves. Annu. Rev. Anim. Biosci. 2018, 6, 1–15. [Google Scholar] [CrossRef]
- Rossi, P.S.; Mattei, R.I.; Schllemer, N.R.; Thomaz, G.R.; Antunes, A.V.; Virmond, M.P.; Taube, M.J.; Bertagnon, H.G. The effect of bovine vaccines against respiratory viruses administered either intranasal or intramuscular on broncho-alveolar fluid cells of heifers. Vet. Q. 2021, 41, 97–106. [Google Scholar] [CrossRef]
- Hill, K.L.; Hunsaker, B.D.; Towsend, H.G.; Littel-van den Hurk, S.v.D.; Griebel, P.J. Mucosal immune response in newborn Holstein calves that had maternally derived antibodies and were vaccinated with an intranasal multivalent modified-live virus vaccine. J. Am. Vet. Med. Assoc. 2012, 240, 1231–1240. [Google Scholar] [CrossRef]
- Xu-Amano, J.; Kiyono, H.; Jackson, R.J.; Staats, H.F.; Fujihashi, K.; Burrows, P.D.; Pillai, S.; McGhee, J.R. Helper T cell subsets for immuno-globulin A responses: Oral immunization with tetanustoxoid and cholera toxin as adjuvant selectively induces Th2 cells in mucosa associated tissues. J. Exp. Med. 1993, 178, 1309–1320. [Google Scholar] [CrossRef]
- Shimizu, Y.; Iwasaki, T.; Tajima, T.; Yuba, E.; Kono, K.; Watari, S. Induction of antibody response in the oral cavity of dogs following intraocular (eye drop) immunization with Porphyromonas gingivalis cell lysate incorporated in pH-sensitive fusogenic polymer-modified liposomes. J. Vet. Med. Sci. 2017, 79, 290–298. [Google Scholar] [CrossRef]
- Chodosh, J.; Nordquist, R.E.; Kennedy, R.C. Comparative anatomy of mammalian conjunctival lymphoid tissue: A putative mucosal immune site. Dev. Comp. Immunol. 1998, 22, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, M.; Nakatsuka, K.; Yamaguchi, T.; Owen, R.L.; Shimada, T. Ocular defense mechanisms with special reference to the demonstration and functional morphology of the conjunctiva-associated lymphoid tissue in Japanese monkeys. Arch. Histol. Cytol. 2006, 69, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Knop, E.; Knop, N. A functional unit for ocular surface immune defense formed by the lacrimal gland, conjunctiva and lacrimal gland drainage system. Adv. Exp. Med. 2002, 506, 835–844. [Google Scholar]
- Siebelmann, S.; Gehlsen, U.; Hüttmann, G.; Koop, N.; Bölke, T.; Gebert, A.; Stern, M.E.; Niederkorn, J.Y.; Steven, P. Development, alteration and real time dynamics of conjunctiva-associated lymphoid tissue. PLoS ONE 2013, 8, e82355. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.Y.; Han, S.J.; Cha, H.R.; Seo, S.U.; Song, J.H.; Chung, S.H.; Kweon, M.-N. Eye mucosa: An efficient vaccine delivery route for inducing protective immunity. J. Immunol. 2010, 185, 3610–3619. [Google Scholar] [CrossRef]
- Liebich, H.G.; Sótonyi, P.; König, H.E. Eye (organum visus). In Veterinary Anatomy of Domestic Mammals Textbook and Colour Atlas, 6th ed.; König, H.E., Liebich, H.G., Eds.; Midori shobo: Tokyo, Japan, 2016; p. 683. [Google Scholar]
- Lohrberg, M.; Pabst, R.; Wilting, J. Co-localization of lymphoid aggregates and lymphatic networks in nose- (NALT) and lacrimal duct-associated lymphoid tissue (LDALT) of mice. BMC Immunol. 2018, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.D.; Han, S.J.; Byun, Y.H.; Yoon, S.C.; Choi, K.S.; Seong, B.L.; Seo, K.Y. Inactivated eyedrop influenza vaccine adjuvanted with poly(I:C) is safe and effective for inducing protective systemic and mucosal immunity. PLoS ONE 2015, 10, e0137608. [Google Scholar] [CrossRef]
- Hassanzadeh, M.; Abdoshah, M.; Yousefi, A.R.; Masoudi, S. Comparison of the impact of different administration routes on the efficacy of a Thermoresistant Newcastle disease vaccine in chickens. Viral Immunol. 2020, 33, 361–366. [Google Scholar] [CrossRef]
- Parsons, K.R.; Howard, C.J.; Jones, B.V.; Sopp, P. Investigation of bovine gut associated lymphoid tissue (GALT) using monoclonal antibodies against bovine lymphocytes. Vet. Pathol. 1989, 26, 396–408. [Google Scholar] [CrossRef]
- Meek, H.C.; Stenfeldt, C.; Arzt, J. Morphological and phenotypic characteristics of the bovine nasopharyngeal mucosa and associated lymphoid tissue. J. Comp. Pathol. 2022, 198, 62–79. [Google Scholar] [CrossRef]
- Anderson, M.L.; Moore, P.F.; Hyde, D.M.; Dungworth, D.L. Bronchus associated lymphoid tissue in the lungs of cattle: Relationship to age. Res. Vet. Sci. 1986, 41, 211–220. [Google Scholar] [CrossRef]
- Casteleyn, C.; Simoens, P.; Van den Broeck, W. Larynx-associated lymphoid tissue (LALT) in young cattle. Vet. Immunol. Immunopathol. 2008, 124, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Hondo, T.; Kanaya, T.; Takakura, I.; Watanabe, H.; Takahashi, Y.; Nagasawa, Y.; Terada, S.; Ohwada, S.; Watanabe, K.; Kitazawa, H.; et al. Cytokeratin 18 is a specific marker of bovine intestinal M cell. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G442–G453. [Google Scholar] [CrossRef] [PubMed]
- Huang, C. Germinal center reaction. Adv. Exp. Med. Biol. 2020, 1254, 47–53. [Google Scholar] [PubMed]
- Yan, Z.; Wang, J.B.; Gong, S.S.; Huang, X. Cell proliferation in the endolymphatic sac in situ after the rat Waldeyer ring equivalent immunostimulation. Laryngoscope 2003, 113, 1609–1614. [Google Scholar] [CrossRef]
- Wyatt, C.R.; Barrett, W.J.; Brackett, E.J.; Davis, W.C.; Besser, T.E. Phenotypic comparison of ileal intraepithelial lymphocyte populations of suckling and weaned calves. Vet. Immunol. Immunopathol. 1999, 67, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, C.R.; Brackett, E.J.; Perryman, L.E.; Rice-Ficht, A.C.; Brown, W.C.; O’Rourke, K.I. Activation of intestinal intraepithelial T lymphocytes in calves infected with Cryptospridium parvum. Infect. Immun. 1997, 65, 185–190. [Google Scholar] [CrossRef]
- Verdugo, P. Goblet cells secretion and mucogenesis. Annu. Rev. Physiol. 1990, 52, 157–176. [Google Scholar] [CrossRef]
- Crespo-Moral, M.; García-Posadas, L.; Lópes-García, A.; Diebold, Y. Histological and immunohistochemical characterization of the porcine ocular surface. PLoS ONE 2020, 15, e0227732. [Google Scholar] [CrossRef]
- Knop, E.; Knop, N.; Claus, P. Local production of secretory IgA in the eye-associated lymphoid tissue (EALT) of the normal human ocular surface. Invest. Ophthalmol. Vis. Sci. 2008, 49, 2322–2329. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Primary Antibody | Dilution | Blocking | Antigen Retrieval | Secondary Antibody |
|---|---|---|---|---|---|
| CD3 | Rabbit monoclonal antibodies, ab16669 (Abcam) | 1:800 | 10% NGS | TB 115 °C, 15 min | Goat anti-rabbit (SABPO kit, Nichirei) |
| CD20 | Rabbit polyclonal antibodies, ab27093 (Abcam) | 1:800 | 10% NGS | TB 115 °C, 15 min | Goat anti-rabbit (SABPO kit, Nichirei) |
| IBA1 | Rabbit polyclonal antibodies, 013-27691 (Wako) | 1:2400 | 10% NGS | CB 115 °C, 15 min | Goat anti-rabbit (SABPO kit, Nichirei) |
| MHCⅡ | Mouse polyclonal antibodies, BOV-CT2012 (Washington State University) | 1:200 | 10% NGS | TB 115 °C, 15 min | Goat anti-mouse IgG (Abcam, ab6788) 1: 500(Biotinylated) |
| CK18 | Mouse monoclonal antibodies, 65028 (Progen Biotechnik) | No need | 10% NGS | TB 115 °C, 15 min | Goat anti-mouse IgG (Abcam, ab6788) 1: 500(Biotinylated) |
| IgA | Rabbit polyclonal antibodies, A10-108A (Bethyl Laboratories) | 1:40,000 | 10% NGS | No need | Goat anti-rabbit (SABPO kit, Nichirei) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosenda, K.; Ichii, O.; Yamashita, Y.; Ohtsuka, H.; Fukuda, S.; Kon, Y. Histological Characteristics of Conjunctiva-Associated Lymphoid Tissue in Young and Adult Holstein Cattle. Animals 2023, 13, 3481. https://doi.org/10.3390/ani13223481
Kosenda K, Ichii O, Yamashita Y, Ohtsuka H, Fukuda S, Kon Y. Histological Characteristics of Conjunctiva-Associated Lymphoid Tissue in Young and Adult Holstein Cattle. Animals. 2023; 13(22):3481. https://doi.org/10.3390/ani13223481
Chicago/Turabian StyleKosenda, Keigo, Osamu Ichii, Yusuke Yamashita, Hiromichi Ohtsuka, Shigeo Fukuda, and Yasuhiro Kon. 2023. "Histological Characteristics of Conjunctiva-Associated Lymphoid Tissue in Young and Adult Holstein Cattle" Animals 13, no. 22: 3481. https://doi.org/10.3390/ani13223481