

Dietary Curcumin Supplementation Could Improve Muscle Quality, Antioxidant Enzyme Activities and the Gut Microbiota Structure of Pelodiscus sinensis
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diet Preparation
2.2. Experimental Turtles and Feeding Management
2.3. Sample Collection
2.4. Muscle Composition Analysis and Texture Profile Analysis
2.5. Serum and Liver Biochemical Analyses
2.6. Gut Microbiota Analysis
2.7. RNA Sequencing and Bioinformatics Analysis
2.8. Quantitative Real-Time PCR Assay (qRT-PCR)
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Muscle-Proximate Composition and Texture Profile
3.3. Serum Biochemistry Indexes
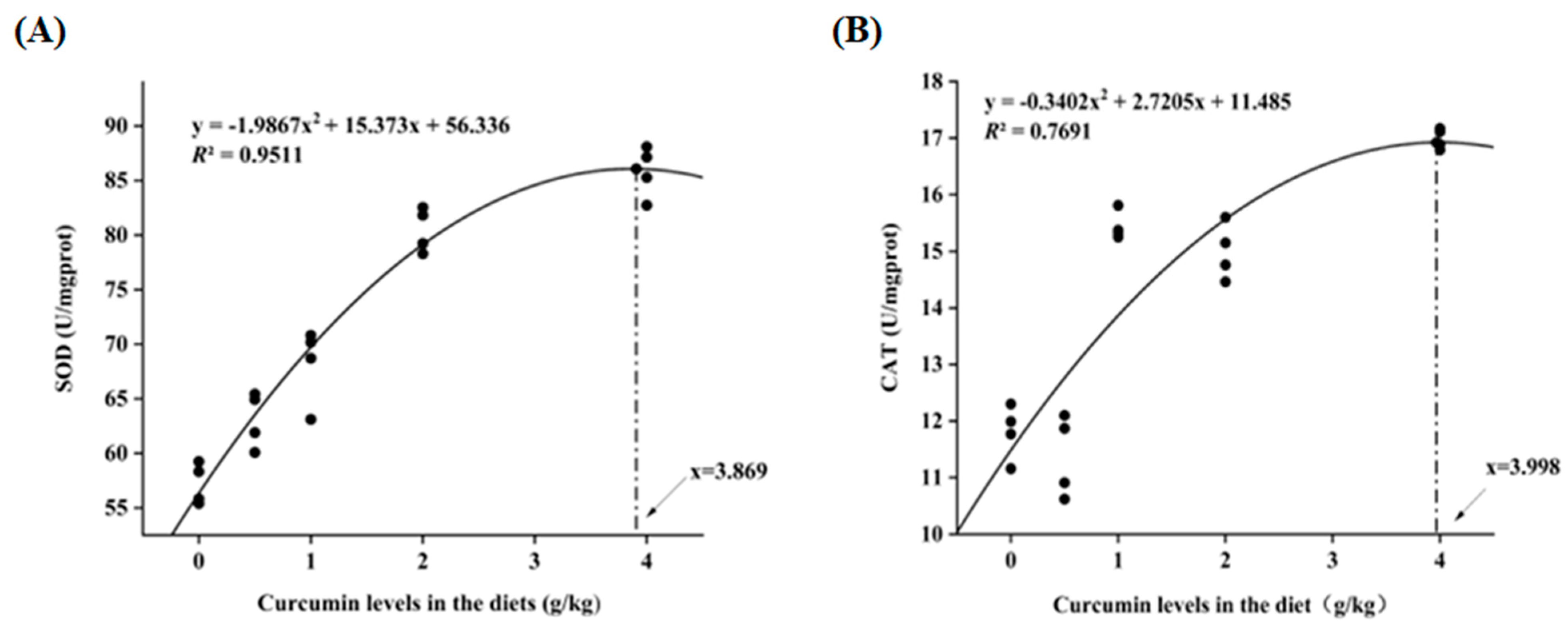
3.4. Antioxidant Enzyme Activities in Serum and Liver
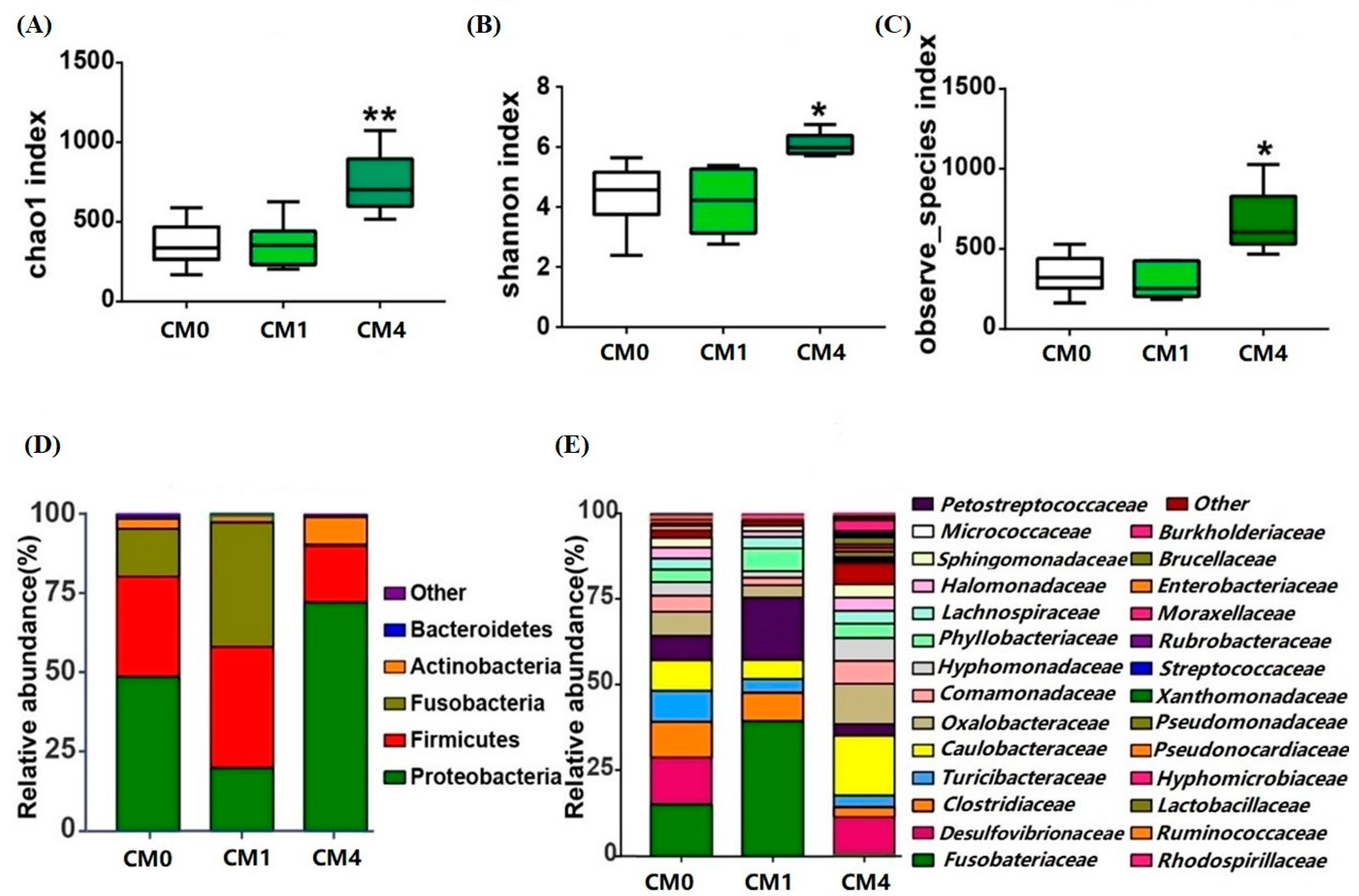
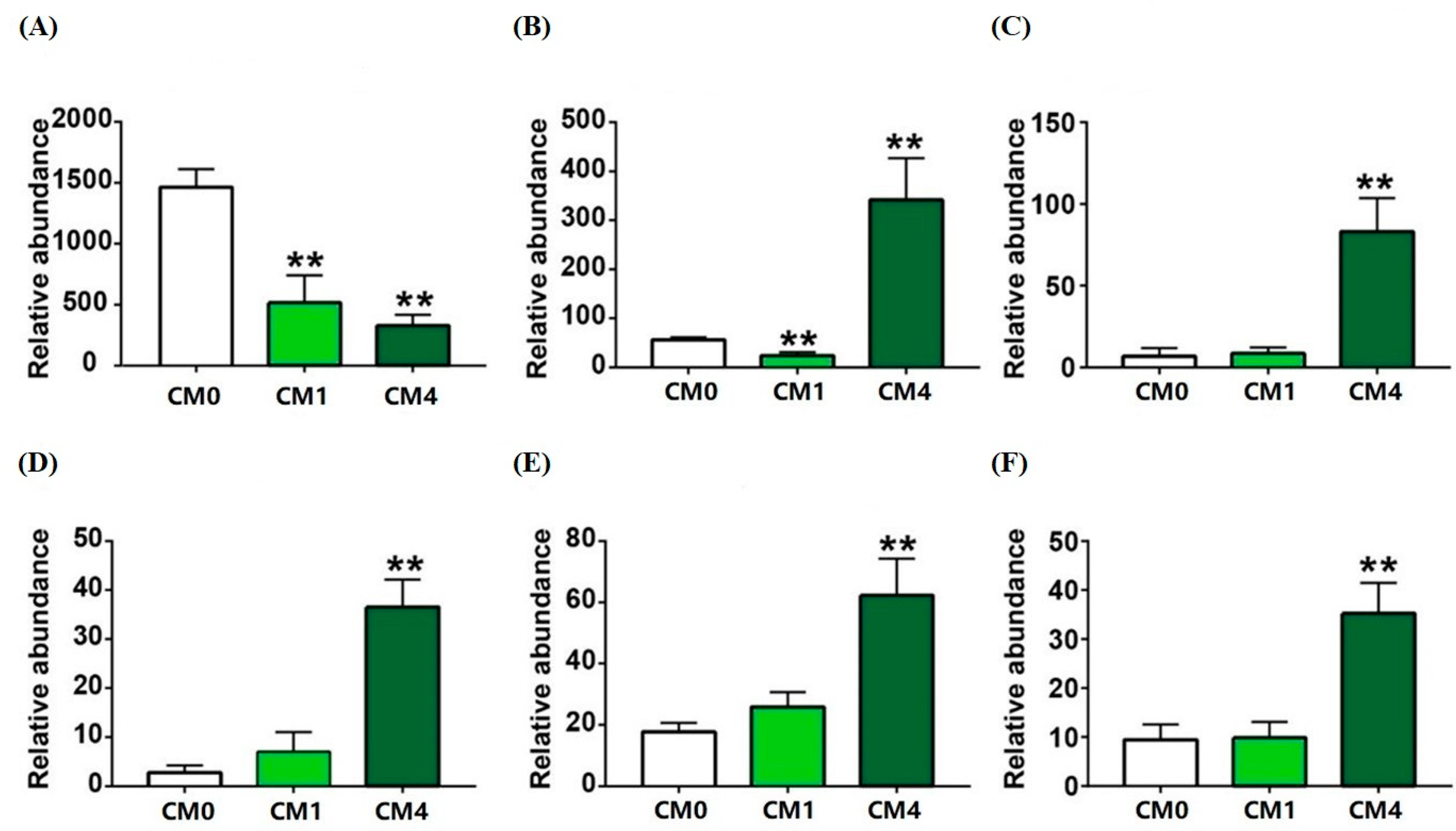
3.5. Gut Microbial Diversity Analysis
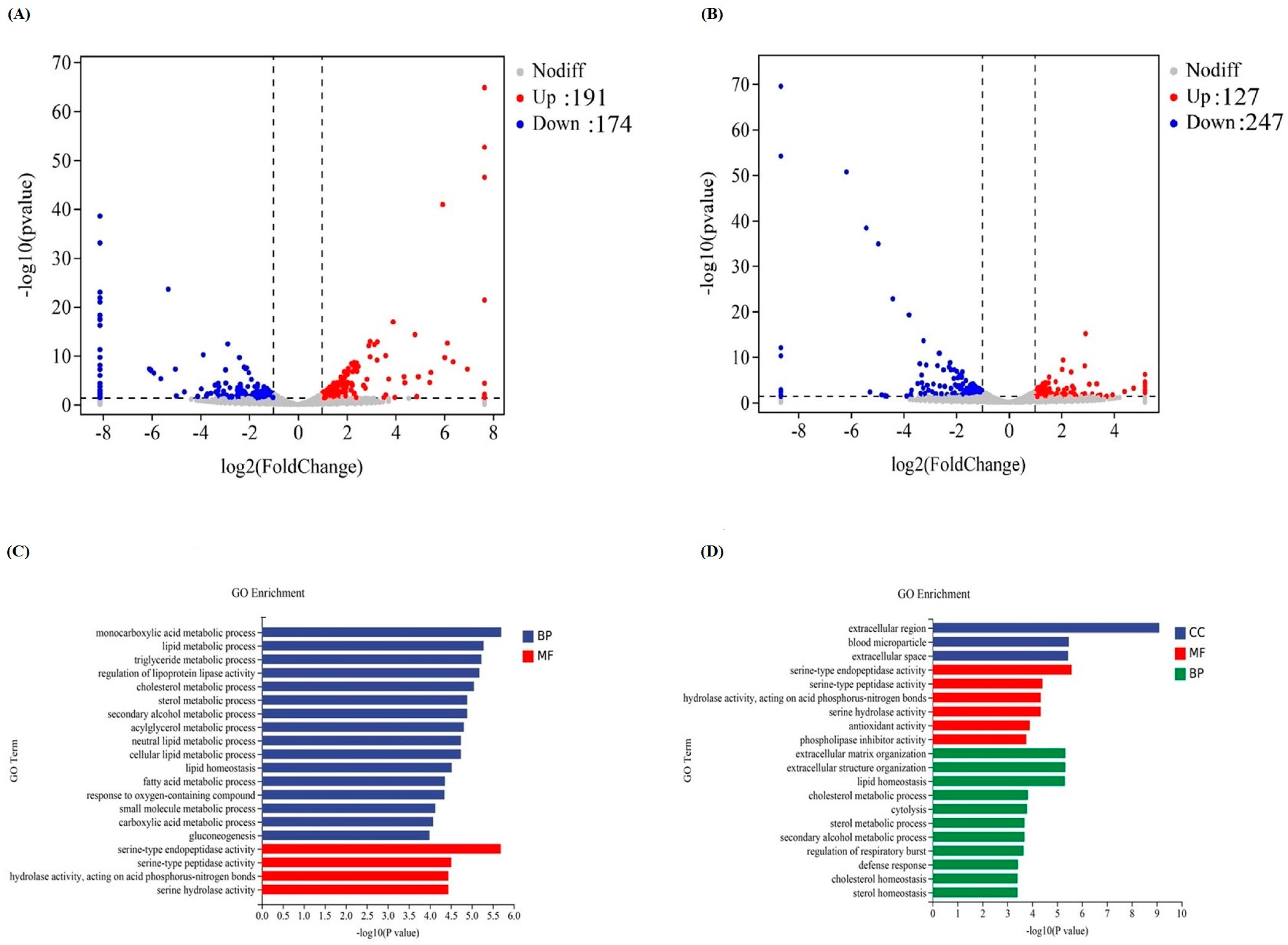
3.6. Liver Transcriptomic Changes
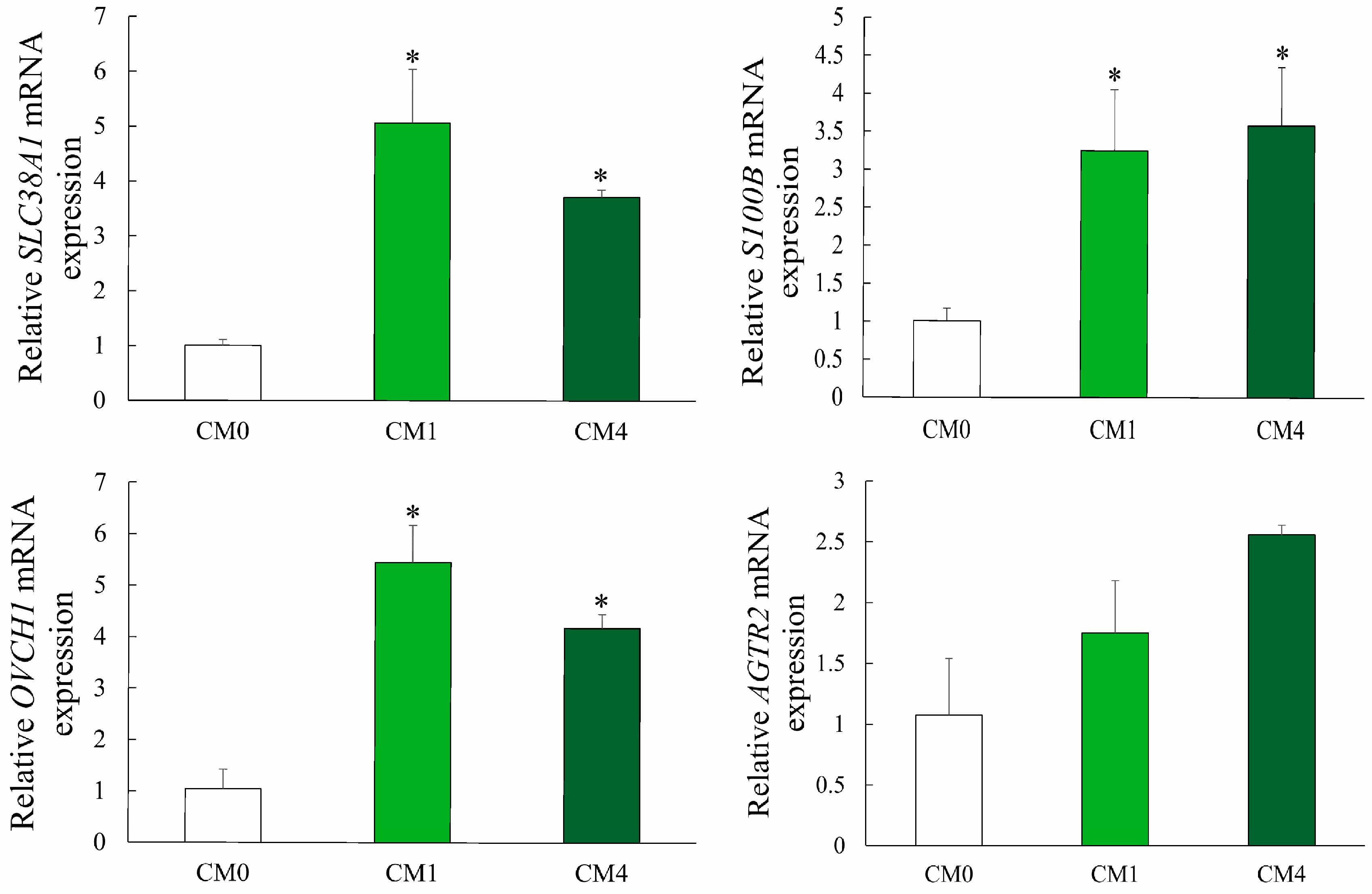
3.7. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kou, H.; Hu, J.; Vijayaraman, S.B.; Wang, A.-L.; Zheng, Y.; Chen, J.; He, G.; Miao, Y.; Lin, L. Evaluation of dietary zinc on antioxidant-related gene expression, antioxidant capability and immunity of soft-shelled turtles Pelodiscus sinensis. Fish Shellfish Immunol. 2021, 118, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tan, S.-L.; Lu, Q.; Xu, R.; Cao, J.; Wu, S.-Q.; Wang, Y.-H.; Zhao, X.-K.; Zhong, Z.-H. Curcumin Suppresses microRNA-7641-Mediated Regulation of p16 Expression in Bladder Cancer. Am. J. Chin. Med. 2018, 46, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- FBMA. China Fishery Statistical Yearbook; China Statistics Press: Beijing, China, 2022. [Google Scholar]
- Wu, X.; Wu, H.; Ye, J. Purification effects of two eco-ditch systems on Chinese soft-shelled turtle greenhouse culture wastewater pollution. Environ. Sci. Pollut. Res. 2014, 21, 5610–5618. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, Q.; Dai, H. Identification of differentially expressed immune-relevant genes in Chinese soft-shelled turtle (Trionyx sinensis) infected with Aeromonas hydrophila. Vet. Immunol. Immunopathol. 2008, 125, 82–91. [Google Scholar] [CrossRef]
- Liu, T.; Han, Y.; Chen, S.; Zhao, H. Genome-wide identification of Toll-like receptors in the Chinese soft-shelled turtle Pelodiscus sinensis and expression analysis responding to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2019, 87, 478–489. [Google Scholar] [CrossRef]
- Xiaoli, C.; Meifeng, L.; Xiangjun, L.; Hua, W.; Fan, W.; Lijuan, Y.; Ming, J.; Xing, L.; Weihua, G.; Wenbing, Z.; et al. Creatine improves the flesh quality of Pacific white shrimp (Litopenaeus vannamei) reared in freshwater. Food Chem. 2021, 354, 129498. [Google Scholar]
- Fan, W.; Juan, T.; Lijuan, Y.; Hua, W.; Ming, J.; Xing, L. Effects of dietary rapeseed meal levels on growth performance, biochemical indices and flesh quality of juvenile genetically improved farmed tilapia. Aquac. Rep. 2021, 20, 100679. [Google Scholar]
- Mahjoob, M.; Stochaj, U. Curcumin nanoformulations to combat aging-related diseases. Ageing Res. Rev. 2021, 69, 101364. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.J.A.; Stojanovski, E.; MacDonald-Wicks, L.; Garg, M.L. Curcumin potentiates cholesterol-lowering effects of phytosterols in hypercholesterolaemic individuals. A randomised controlled trial. Metabolism 2018, 82, 22–35. [Google Scholar] [CrossRef]
- Xia, Z.-H.; Zhang, S.-Y.; Chen, Y.-S.; Li, K.; Chen, W.-B.; Liu, Y.-Q. Curcumin anti-diabetic effect mainly correlates with its anti-apoptotic actions and PI3K/Akt signal pathway regulation in the liver. Food Chem. Toxicol. 2020, 146, 111803. [Google Scholar] [CrossRef] [PubMed]
- Abu-Taweel, G.M.; Attia, M.F.; Hussein, J.; Mekawi, E.M.; Galal, H.M.; Ahmed, E.I.; Allam, A.A.; El-Naggar, M.E. Curcumin nanoparticles have potential antioxidant effect and restore tetrahydrobiopterin levels in experimental diabetes. Biomed. Pharmacother. 2020, 131, 110688. [Google Scholar] [CrossRef] [PubMed]
- Jahanbakhshi, F.; Maleki Dana, P.; Badehnoosh, B.; Yousefi, B.; Mansournia, M.A.; Jahanshahi, M.; Asemi, Z.; Halajzadeh, J. Curcumin anti-tumor effects on endometrial cancer with focus on its molecular targets. Cancer Cell Int. 2021, 21, 120. [Google Scholar] [CrossRef]
- Nm, J.; Joseph, A.; Maliakel, B.; Im, K. Dietary addition of a standardized extract of turmeric (TurmaFEEDTM) improves growth performance and carcass quality of broilers. J. Anim. Sci. Technol. 2018, 60, 8. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, A.; Senthil, N.; Min, T. Nanocurcumin: A Promising Candidate for Therapeutic Applications. Front. Pharmacol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Sanidad, K.Z.; Sukamtoh, E.; Xiao, H.; McClements, D.J.; Zhang, G. Curcumin: Recent advances in the development of strategies to improve oral bioavailability. Annu. Rev. Food Sci. Technol. 2019, 10, 597–617. [Google Scholar] [CrossRef] [Green Version]
- Xavier, M.J.; Navarro-Guillén, C.; Lopes, A.; Colen, R.; Teodosio, R.; Mendes, R.; Oliveira, B.; Valente, L.M.P.; Conceição, L.E.C.; Engrola, S. Effects of dietary curcumin in growth performance, oxidative status and gut morphometry and function of gilthead seabream postlarvae. Aquac. Rep. 2022, 24, 101128. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Alagawany, M.; Abdel-Moneim, A.-M.E.; Mohammed, N.G.; Khafaga, A.F.; Bin-Jumah, M.; Othman, S.I.; Allam, A.A.; Elnesr, S.S. Cinnamon (Cinnamomum zeylanicum) Oil as a Potential Alternative to Antibiotics in Poultry. Antibiotics 2020, 9, 210. [Google Scholar] [CrossRef]
- Yonar, M.E.; Mişe Yonar, S.; İspir, Ü.; Ural, M.Ş. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. achromogenes. Fish Shellfish Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 21st ed.; AOAC: Arlington, VA, USA, 2019. [Google Scholar]
- Yu, L.J.; Wu, F.; Jiang, M.; Yang, C.G.; Liu, W.; Tian, J.; Lu, X.; Wen, H. Ferulic acid: A natural compound as an efficient feed additive for GIFT (Oreochromis niloticus). Aquac. Nutr. 2018, 24, 27–35. [Google Scholar] [CrossRef]
- Yu, L.; Wen, H.; Jiang, M.; Wu, F.; Tian, J.; Lu, X.; Xiao, J.; Liu, W. Effects of ferulic acid on intestinal enzyme activities, morphology, microbiome composition of genetically improved farmed tilapia (Oreochromis niloticus) fed oxidized fish oil. Aquaculture 2020, 528, 735543. [Google Scholar] [CrossRef]
- Lu, X.; Wen, H.; Li, Q.; Wang, G.; Li, P.; Chen, J.; Sun, Y.; Yang, C.; Wu, F. Comparative analysis of growth performance and liver transcriptome response of juvenile Ancherythroculter nigrocauda fed diets with different protein levels. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100592. [Google Scholar] [CrossRef]
- Li, Y.; Lu, X.; Gao, W.; Yu, L.; Wen, H.; Jiang, M.; Tian, J.; Wu, F. The effect of dietary paeonol on growth performance, antioxidant enzyme activities and gene expressions of Genetic Improvement of Farmed Tilapia juveniles (Oreochromis niloticus). Aquac. Rep. 2022, 26, 101302. [Google Scholar] [CrossRef]
- Lu, X.; Wei, H.; Yang, C.; Li, Q.; Li, P.; Chen, J.; Sun, Y.; Wen, H.; Jiang, M.; Wang, G. Effects of dietary protein levels on growth performance and liver transcriptome changes in juvenile top-mouth culter Erythroculter ilishaeformis. Aquac. Rep. 2021, 21, 100964. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.-F.; Qian, H.-C.; Wang, J.-L.; Chen, Z.; Ordovas, J.M.; Lai, C.-Q.; Shen, L.-R. Supplementation with turmeric residue increased survival of the Chinese soft-shelled turtle (Pelodiscus sinensis) under high ambient temperatures. J. Zhejiang Univ.-SCIENCE B 2018, 19, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wu, X.-Y.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Mohamed, A.A.-R.; El-Houseiny, W.; El-Murr, A.E.; Ebraheim, L.L.M.; Ahmed, A.I.; El-Hakim, Y.M.A. Effect of hexavalent chromium exposure on the liver and kidney tissues related to the expression of CYP450 and GST genes of Oreochromis niloticus fish: Role of curcumin supplemented diet. Ecotoxicol. Environ. Saf. 2020, 188, 109890. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.O.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2021, 532, 736030. [Google Scholar] [CrossRef]
- Ejaz, A.; Wu, D.; Kwan, P.; Meydani, M. Curcumin inhibits adipogenesis in 3T3-L1 adipocytes and angiogenesis and obesity in C57/BL mice. J. Nutr. 2009, 139, 919–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/β-catenin signaling. Am. J. Physiol. Cell Physiol. 2010, 298, C1510–C1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Yang, H.; Liu, F.; Pang, Q.; Shan, A.; Feng, X. Effect of dietary curcumin supplementation on duck growth performance, antioxidant capacity and breast meat quality. Foods 2021, 10, 2981. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Gerbet, R.R.; Griss, L.G.; Fortuoso, B.F.; Petrolli, T.G.; Boiago, M.M.; Souza, C.F.; Baldissera, M.D.; Mesadri, J.; Wagner, R. Combination of herbal components (curcumin, carvacrol, thymol, cinnamaldehyde) in broiler chicken feed: Impacts on response parameters, performance, fatty acid profiles, meat quality and control of coccidia and bacteria. Microb. Pathog. 2020, 139, 103916. [Google Scholar] [CrossRef]
- Abbas, W.T.; Ibrahim, T.B.E.-D.; Elgendy, M.Y.; Zaher, M.F.A. Effect of Curcumin on Iron Toxicity and Bacterial Infection in Catfish (Clarias gariepinus). Pak. J. Biol. Sci. 2019, 22, 510–517. [Google Scholar] [CrossRef]
- Cao, L.; Ding, W.; Du, J.; Jia, R.; Liu, Y.; Zhao, C.; Shen, Y.; Yin, G. Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinus carpio var. Jian) with CCl4-induced liver damage. Fish Shellfish Immunol. 2015, 43, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, X.; Li, W.; Deng, X.; Deng, Y.; Niu, X. Curcumin protects mice from Staphylococcus aureus pneumonia by interfering with the self-assembly process of α-hemolysin. Sci. Rep. 2016, 6, 28254. [Google Scholar] [CrossRef] [Green Version]
- Yap, R.W.K.; Shidoji, Y.; Yap, W.S.; Masaki, M. Association and interaction effect of AGTR1 and AGTR2 gene polymorphisms with dietary pattern on metabolic risk factors of cardiovascular disease in Malaysian adults. Nutrients 2017, 9, 853. [Google Scholar] [CrossRef] [Green Version]
- Baum, L.; Cheung, S.K.; Mok, V.C.; Lam, L.C.; Leung, V.P.; Hui, E.; Ng, C.C.; Chow, M.; Ho, P.C.; Lam, S. Curcumin effects on blood lipid profile in a 6-month human study. Pharmacol. Res. 2007, 56, 509–514. [Google Scholar] [CrossRef]
- Peschel, D.; Koerting, R.; Nass, N. Curcumin induces changes in expression of genes involved in cholesterol homeostasis. J. Nutr. Biochem. 2007, 18, 113–119. [Google Scholar] [CrossRef] [PubMed]
- DiSilvestro, R.A.; Joseph, E.; Zhao, S.; Bomser, J. Diverse effects of a low dose supplement of lipidated curcumin in healthy middle aged people. Nutr. J. 2012, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Priyadarsini, K.I.; Maity, D.K.; Naik, G.; Kumar, M.S.; Unnikrishnan, M.; Satav, J.; Mohan, H. Role of phenolic OH and methylene hydrogen on the free radical reactions and antioxidant activity of curcumin. Free Radic. Biol. Med. 2003, 35, 475–484. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Bhatt, I.D.; Ichikawa, H.; Ahn, K.S.; Sethi, G.; Sandur, S.K.; Natarajan, C.; Seeram, N.; Shishodia, S. 10 Curcumin—Biological and Medicinal Properties. 2006. Available online: https://www.researchgate.net/publication/235993633_Curcumin_-_Biologival_and_medicinal_properties#fullTextFileContent (accessed on 15 March 2023).
- Gu, H.; Yuan, Y.; Cai, M.; Wang, D.; Lv, W. Toxicity of isoprocarb to earthworms (Eisenia fetida): Oxidative stress, neurotoxicity, biochemical responses and detoxification mechanisms. Environ. Pollut. 2021, 290, 118038. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shah, S.W.A.; Zhou, Q.; Yin, X.; Teng, X. The contributions of miR-25-3p, oxidative stress, and heat shock protein in a complex mechanism of autophagy caused by pollutant cadmium in common carp (Cyprinus carpio L.) hepatopancreas. Environ. Pollut. 2021, 287, 117554. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Miao, Z.; Wang, S.; Shi, X.; Xu, S. Quercetin antagonizes imidacloprid-induced mitochondrial apoptosis through PTEN/PI3K/AKT in grass carp hepatocytes. Environ. Pollut. 2021, 290, 118036. [Google Scholar] [CrossRef]
- Xiong, J.-B.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70. [Google Scholar]
- Nie, L.; Zhou, Q.-J.; Qiao, Y.; Chen, J. Interplay between the gut microbiota and immune responses of ayu (Plecoglossus altivelis) during Vibrio anguillarum infection. Fish Shellfish Immunol. 2017, 68, 479–487. [Google Scholar] [CrossRef]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wu, W.; Hu, G.; Qiu, L.; Meng, S.; Song, C.; Fan, L.; Zhao, Z.; Bing, X.; Chen, J. Gut microbiota analysis of juvenile genetically improved farmed tilapia (Oreochromis niloticus) by dietary supplementation of different resveratrol concentrations. Fish Shellfish Immunol. 2018, 77, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Jiang, M.; Lu, X.; Wen, H. Effects of dietary protein level on the gut microbiome and nutrient metabolism in tilapia (Oreochromis niloticus). Animals 2021, 11, 1024. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-Y.; Chen, X.-L.; Zhao, H.-L.; Dang, H.-Y.; Luan, X.-W.; Zhang, X.-Y.; He, H.-L.; Zhou, B.-C.; Zhang, Y.-Z. Diversity of both the cultivable protease-producing bacteria and their extracellular proteases in the sediments of the South China Sea. Microb. Ecol. 2009, 58, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Hossain, T.J.; Das, M.; Ali, F.; Chowdhury, S.I.; Zedny, S.A. Substrate preferences, phylogenetic and biochemical properties of proteolytic bacteria present in the digestive tract of Nile tilapia (Oreochromis niloticus). AIMS Microbiol. 2021, 7, 528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-N.; Zhang, J.-L.; Guan, W.-C.; Zhang, X.-F.; Guan, S.-H.; Zeng, Q.-H.; Cheng, G.-F.; Cui, W. Effects of Lactobacillus delbrueckii on immune response, disease resistance against Aeromonas hydrophila, antioxidant capability and growth performance of Cyprinus carpio Huanghe var. Fish Shellfish Immunol. 2017, 68, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Xu, J.; Lian, F.; Yu, X.; Zhao, Y.; Xu, L.; Zhang, M.; Zhao, X.; Shen, J.; Wu, S. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional Chinese herbal formula: A multicenter, randomized, open label clinical trial. MBio 2018, 9, e02392-17. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gao, J.; Huang, S. Effects of different dietary phospholipid levels on growth performance, fatty acid composition, PPAR gene expressions and antioxidant responses of blunt snout bream Megalobrama amblycephala fingerlings. Fish Physiol. Biochem. 2015, 41, 423–436. [Google Scholar] [CrossRef]
- Jackson, B.C.; Carpenter, C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse forkhead box (FOX) gene families. Hum. Genom. 2010, 4, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Hong, P.-P.; Yang, M.-C.; Zhao, X.-F.; Wang, J.-X. FOXO regulates the expression of antimicrobial peptides and promotes phagocytosis of hemocytes in shrimp antibacterial immunity. PLoS Pathog. 2021, 17, e1009479. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CM0 | CM0.5 | CM1 | CM2 | CM4 | |
|---|---|---|---|---|---|
| 1 I BW (g) | 162.56 ± 4.50 | 159.61 ± 1.33 | 160.00 ± 3.15 | 158.89 ± 1.90 | 166.56 ± 0.97 |
| 2 FBW (g) | 221.75 ± 6.35 | 223.31 ± 4.31 | 226.29 ± 1.66 | 227.51 ± 8.94 | 237.50 ± 4.45 |
| 3 WG (%) | 37.53 ± 5.78 | 39.93 ± 3.05 | 42.00 ± 5.86 | 43.21 ± 5.67 | 44.59 ± 4.97 |
| 4 FCR | 2.36 ± 0.11 | 2.26 ± 0.15 | 2.21 ± 0.06 | 2.30 ± 0.17 | 2.29 ± 0.08 |
| 5 SR | 79.00 ± 2.31 a | 84.35 ± 1.61 a | 81.33 ± 2.83 a | 82.16 ± 2.66 a | 93.03 ± 2.67 b |
| CM0 | CM0.5 | CM1 | CM2 | CM4 | |
|---|---|---|---|---|---|
| Moisture (%) | 75.48 ± 5.12 | 73.77 ± 0.93 | 74.95 ± 0.97 | 74.23 ± 2.62 | 73.26 ± 2.00 |
| Crude protein (%) | 13.14 ± 0.79 a | 16.22 ± 0.20 b | 16.49 ± 0.12 b | 16.33 ± 1.26 b | 17.60 ± 0.30 c |
| Crude lipid (%) | 5.62 ± 0.78 b | 5.20 ± 1.04 b | 4.17 ± 0.65 a | 3.76 ± 0.36 a | 3.92 ± 0.32 a |
| Ash (%) | 5.40 ± 0.38 | 5.62 ± 1.32 | 5.43 ± 0.20 | 5.56 ± 0.19 | 5.44 ± 0.22 |
| CM0 | CM0.5 | CM1 | CM2 | CM4 | |
|---|---|---|---|---|---|
| Hardness (g) | 957.75 ± 83.90 a | 997.25 ± 77.47 a | 1287.00 ± 120.53 b | 1251.25 ± 106.52 b | 1308.00 ± 115.00 b |
| Springiness | 0.41 ± 0.04 | 0.40 ± 0.03 | 0.38 ± 0.02 | 0.45 ± 0.04 | 0.45 ± 0.06 |
| Cohesiveness | 0.51 ± 0.08 | 0.55 ± 0.06 | 0.48 ± 0.06 | 0.60 ± 0.03 | 0.52 ± 0.05 |
| Gumminess (g) | 570.90 ± 67.75 a | 577.29 ± 43.22 a | 606.26 ± 64.67 ab | 661.98 ± 43.59 ab | 734.31 ± 78.18 b |
| Chewiness (g) | 178.12 ± 12.71 a | 181.09 ± 15.32 a | 219.43 ± 16.79 b | 220.47 ± 21.87 b | 184.49 ± 14.52 ab |
| Resilience | 0.26 ± 0.05 | 0.26 ± 0.05 | 0.26 ± 0.03 | 0.25 ± 0.04 | 0.25 ± 0.04 |
| CM0 | CM0.5 | CM1 | CM2 | CM4 | |
|---|---|---|---|---|---|
| Serum | |||||
| SOD (U/mgprot) | 57.20 ± 1.88 a | 63.08 ± 2.54 b | 68.20 ± 3.51 b | 80.47 ± 2.03 c | 85.81 ± 2.36 c |
| Liver | |||||
| CAT (U/mgprot) | 11.80 ± 0.48 a | 11.37 ± 0.72 a | 15.43 ± 0.25 b | 14.99 ± 0.49 b | 16.99 ± 0.18 c |
| T-AOC (U/mgprot) | 2.87 ± 0.09 a | 2.95 ± 0.08 ab | 3.06 ± 0.04 b | 3.01 ± 0.08 ab | 3.06 ± 0.07 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.-Y.; Wen, H.; Jiang, M.; Tian, J.; Dong, L.-X.; Shi, Z.-C.; Zhou, T.; Lu, X.; Liang, H.-W. Dietary Curcumin Supplementation Could Improve Muscle Quality, Antioxidant Enzyme Activities and the Gut Microbiota Structure of Pelodiscus sinensis. Animals 2023, 13, 2626. https://doi.org/10.3390/ani13162626
Jiang J-Y, Wen H, Jiang M, Tian J, Dong L-X, Shi Z-C, Zhou T, Lu X, Liang H-W. Dietary Curcumin Supplementation Could Improve Muscle Quality, Antioxidant Enzyme Activities and the Gut Microbiota Structure of Pelodiscus sinensis. Animals. 2023; 13(16):2626. https://doi.org/10.3390/ani13162626
Chicago/Turabian StyleJiang, Jia-Yuan, Hua Wen, Ming Jiang, Juan Tian, Li-Xue Dong, Ze-Chao Shi, Tong Zhou, Xing Lu, and Hong-Wei Liang. 2023. "Dietary Curcumin Supplementation Could Improve Muscle Quality, Antioxidant Enzyme Activities and the Gut Microbiota Structure of Pelodiscus sinensis" Animals 13, no. 16: 2626. https://doi.org/10.3390/ani13162626