
Autochthonous Bacilli and Fructooligosaccharide as Functional Feed Additives Improve Growth, Feed Utilisation, Haemato-Immunological Parameters and Disease Resistance in Rohu, Labeo rohita (Hamilton)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Probiotics and Prebiotic Sources
2.3. Diet Preparation
2.4. Experimental Design
2.5. Proximate Composition of Experimental Diets and Fish Carcass
2.6. Growth Indices
2.7. Estimation of Digestive Enzymes
2.8. Collection of Blood Samples
2.9. Collection of Skin Mucus and Head Kidney Leucocytes
2.10. Analysis of Haemato-Biochemical Profile
2.11. Estimation of Immunological Parameters
2.12. Estimation of Stress Enzymes
2.13. Statistical Analysis
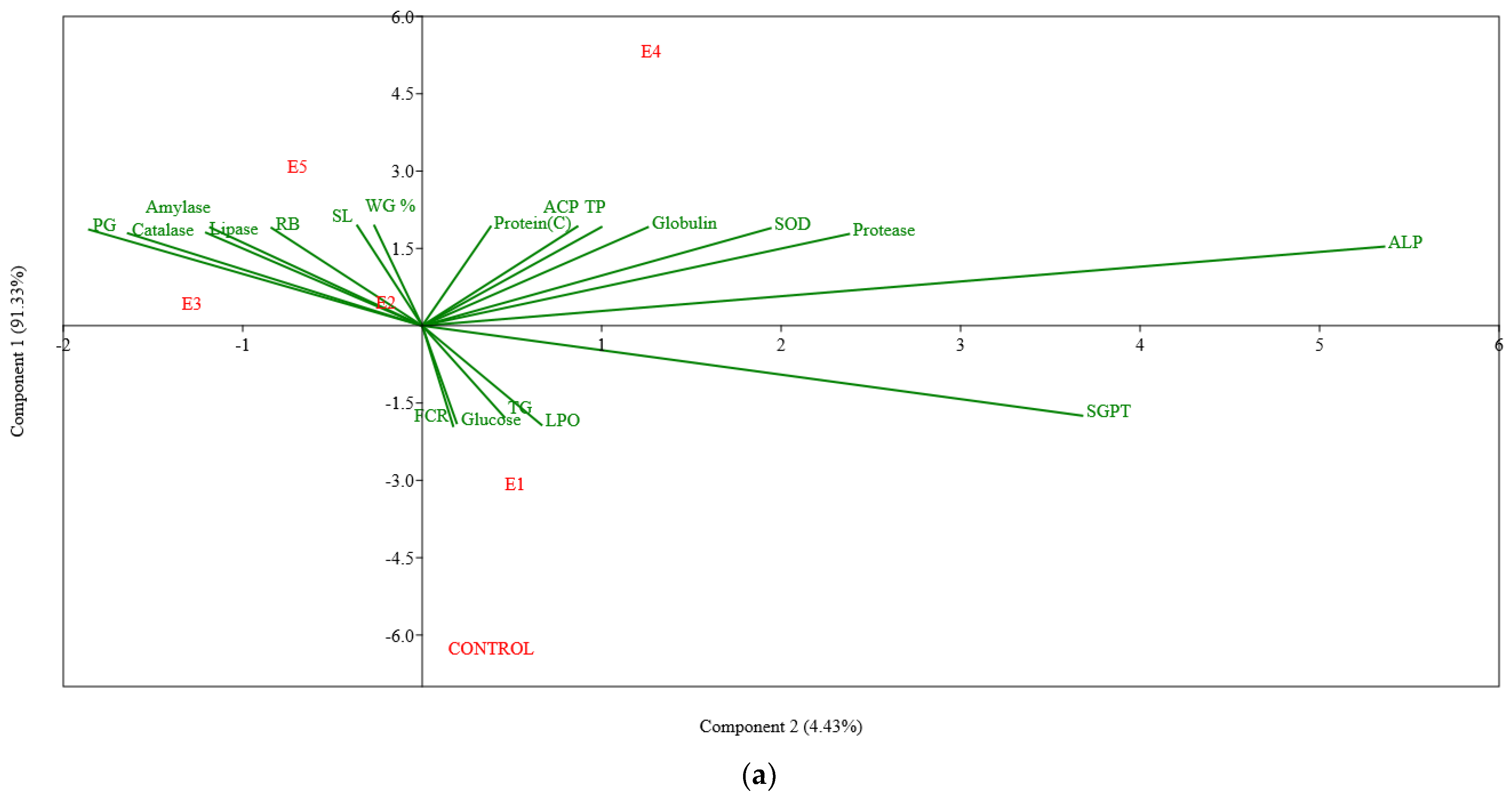
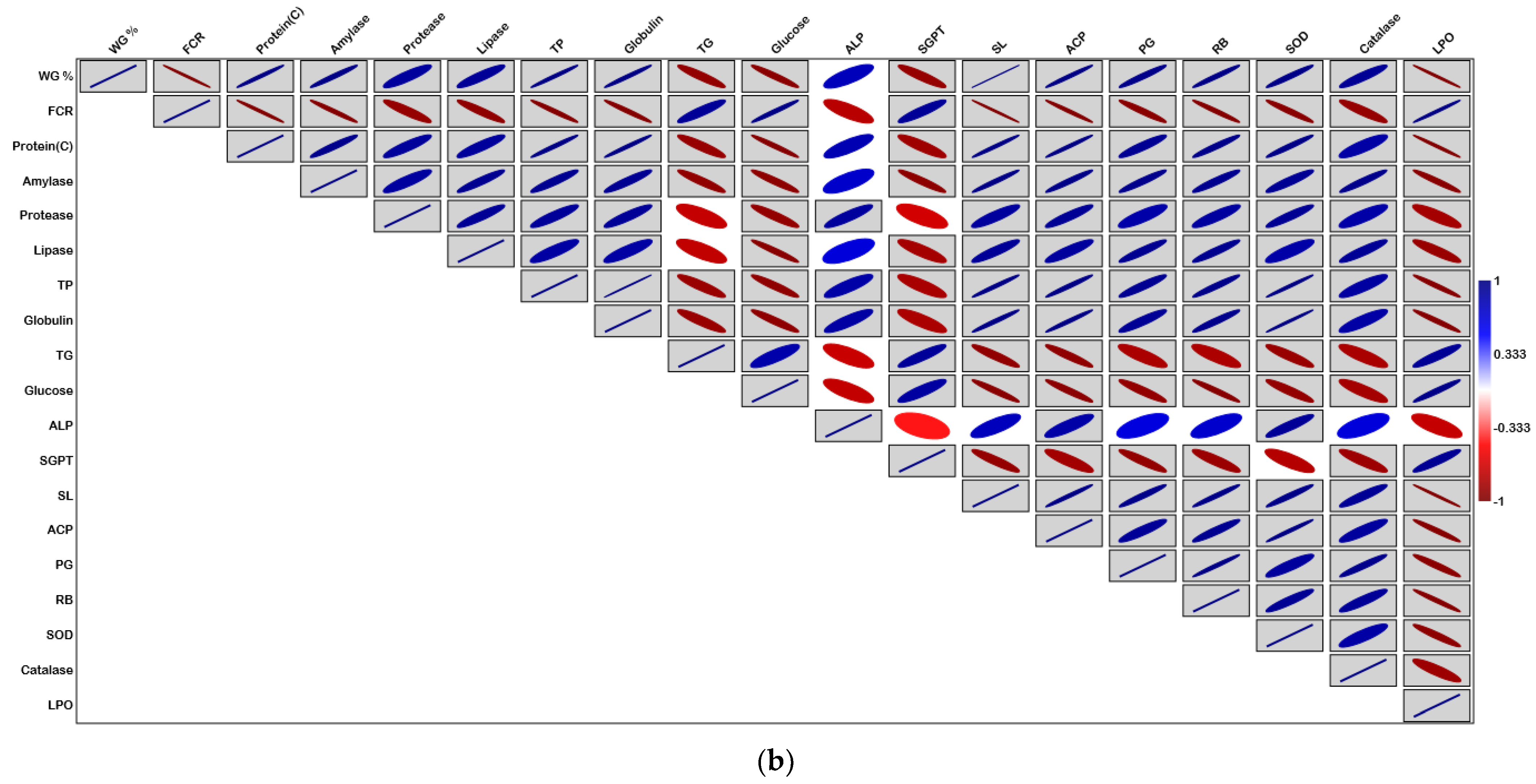
3. Results
3.1. Growth Parameters
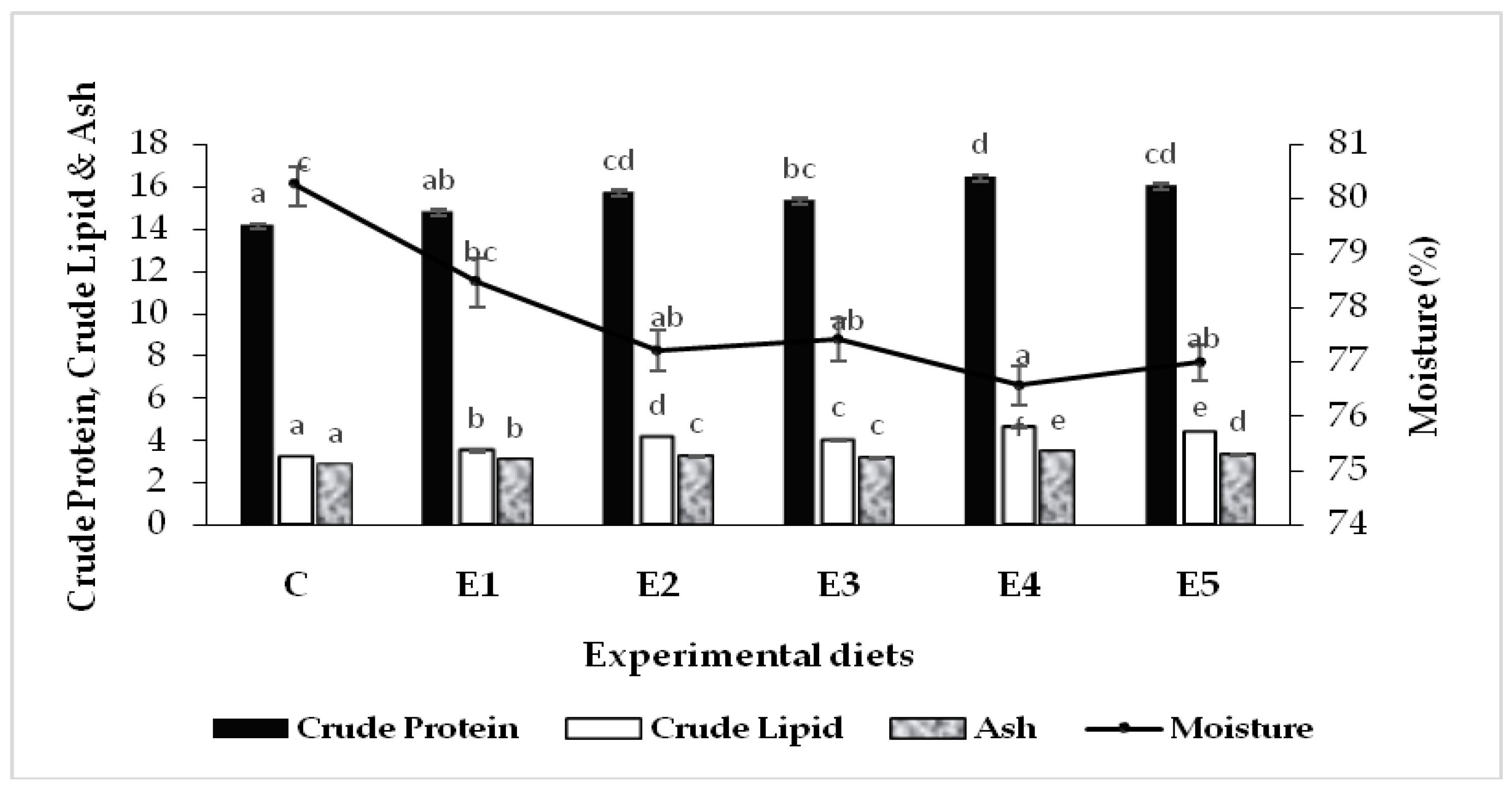
3.2. Carcass Composition
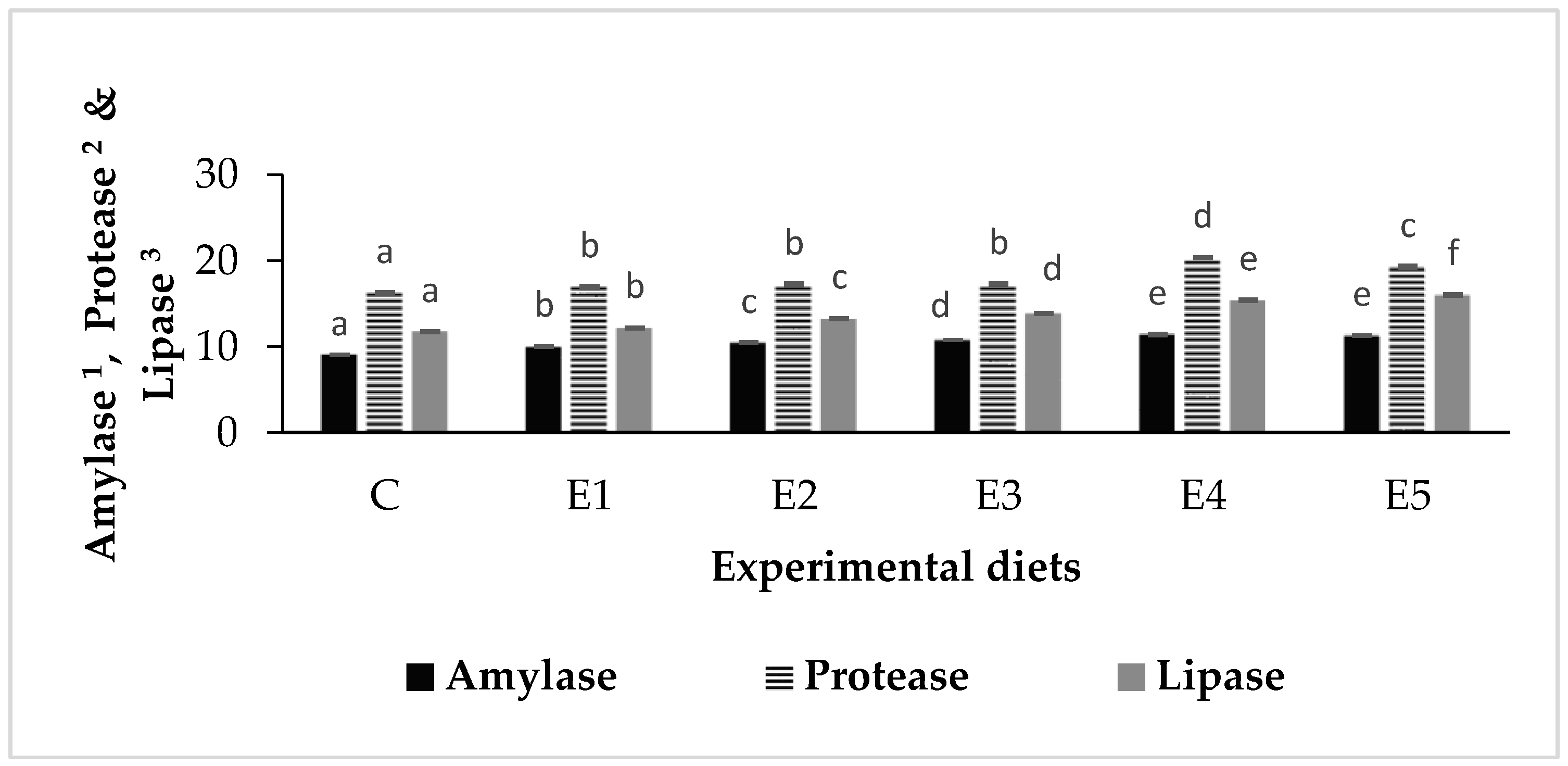
3.3. Digestive Enzymes
3.4. Haematological Parameters
3.5. Serum Biochemical Parameters
3.6. Immune Parameters
3.7. Stress Parameters
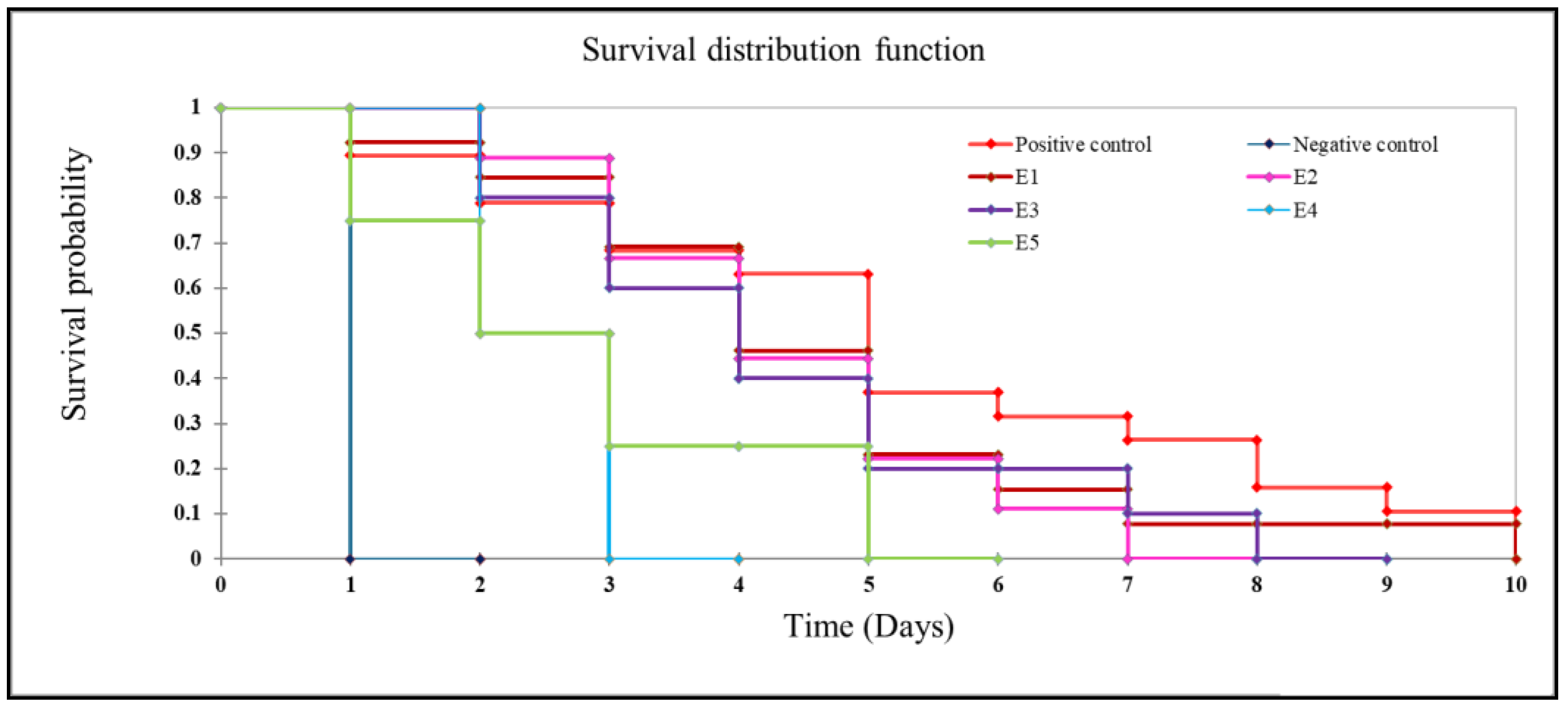
3.8. Post-Challenge Survivability
4. Discussion
4.1. Growth Performance and Feed Utilisation
4.2. Haematological Parameters
4.3. Serum Biochemical Parameters
4.4. Immune Parameters
4.5. Stress Parameters
4.6. Challenge Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giri, S.S.; Sukumaran, V.; Sen, S.S.; Jena, P.K. Effects of dietary supplementation of potential probiotic Bacillus subtilis VSG1 singularly or in combination with Lactobacillus plantarum VSG3 or/and Pseudomonas aeruginosa VSG2 on the growth, immunity and disease resistance of Labeo rohita. Aquac. Nutr. 2014, 20, 163–171. [Google Scholar] [CrossRef]
- Mukherjee, A.; Chandra, G.; Ghosh, K. Single or conjoint application of autochthonous Bacillus strains as potential probiotics: Effects on growth, feed utilization, immunity and disease resistance in Rohu, Labeo rohita (Hamilton). Aquaculture 2019, 512, 734302. [Google Scholar]
- Devi, G.; Harikrishnan, R.; Paray, B.A.; Al-Sadoon, M.K.; Hoseinifar, S.H.; Balasundaram, C. Effect of symbiotic supplemented diet on innate-adaptive immune response, cytokine gene regulation and antioxidant property in Labeo rohita against Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 89, 687–700. [Google Scholar]
- Kavitha, M.; Raja, M.; Perumal, P. Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822). Aquac. Rep. 2018, 11, 59–69. [Google Scholar] [CrossRef]
- Dutta, D.; Ghosh, K. Improvement of growth, nutrient utilization and haemato-immunological parameters in rohu, Labeo rohita (Hamilton) using Bacillus tequilensis (KF623287) through diets or as water additive. Aquac. Nutr. 2021, 27, 29–47. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Mirvaghefi, A.; Amoozegar, M.A.; Sharifian, M.; Esteban, M.A. Modulation of innate immune response, mucosal parameters and disease resistance in rainbow trout (Oncorhynchus mykiss) upon synbiotic feeding. Fish Shellfish Immunol. 2015, 45, 27–32. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front Microbiol. 2018, 9, 2429. [Google Scholar] [PubMed] [Green Version]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture 2008, 274, 1–14. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Nayak, S.K. Multifaceted applications of probiotic Bacillus species in aquaculture with special reference to Bacillus subtilis. Rev. Aquac. 2021, 13, 862–906. [Google Scholar]
- Ai, Q.; Xu, H.; Mai, K.; Xu, W.; Wang, J.; Zhang, W. Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, nonspecific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture 2011, 317, 155–161. [Google Scholar] [CrossRef]
- van Doan, H.; Hoseinifar, S.H.; Dawood, M.A.O.; Chitmanat, C.; Tayyamath, K. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Soltani, M.; Ghosh, K.; Hoseinifar, S.H.; Kumar, V.; Lymbery, A.J.; Roy, S.; Ringo, E. Genus Bacillus, promising probiotics in aquaculture: Aquatic animal origin, bioactive components, bioremediation and efficacy in fish and shellfish. Rev. Fish. Sci. Aquac. 2019, 27, 331–379. [Google Scholar] [CrossRef] [Green Version]
- Talpur, A.D.; Munir, M.B.; Mary, A.; Hashim, R. Dietary probiotics and prebiotics improved food acceptability, growth performance, haematology and immunological parameters and disease resistance against Aeromonas hydrophila in snakehead (Channa striata) fingerlings. Aquaculture 2014, 426–427, 14–20. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.E.; Gifstad, T.O.; Dalmo, R.A.; Amlund, H.; Hemre, G.-I.; Bakke, A.M. Prebiotics in aquaculture: A review. Aquac. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Rodrigues, M.V.; Zanuzzo, F.S.; Koch, J.F.A.; de Oliveira, C.A.F.; Sima, P.; Vetvicka, V. Development of Fish Immunity and the Role of β-Glucan in Immune Responses. Molecules 2020, 25, 5378. [Google Scholar] [CrossRef]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish Physiol. Biochem. 2013, 39, 431–457. [Google Scholar]
- Sang, H.M.; Kien, N.T.; Thuy, N.T.T. Effects of dietary mannan oligosaccharide on growth, survival, physiological, immunological and gut morphological conditions of black tiger prawn (Penaeus monodon Fabricius 1798). Aquac. Nutr. 2014, 20, 341–348. [Google Scholar] [CrossRef]
- Soleimani, N.; Hoseinifar, S.H.; Merrifield, D.L.; Barati, M.; Abadi, Z.H. Dietary supplementation of fructooligosaccharide (FOS) improves the innate immune response, stress resistance, digestive enzyme activities and growth performance of Caspian roach (Rutilus rutilus) fry. Fish Shellfish Immunol. 2012, 32, 316–321. [Google Scholar] [PubMed]
- Singh, S.K.; Tiwari, V.K.; Chadha, N.K.; Munilkumar, S.; Prakash, C.; Pawar, N.A. Effect of dietary synbiotic supplementation on growth, immune and physiological status of Labeo rohita juveniles exposed to low pH stress. Fish Shellfish Immunol. 2019, 91, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Mahious, A.S.; Gatesoupe, F.J.; Hervi, M.; Metailler, R.; Ollevier, F. Effect of dietary inulin and oligosaccharides as prebiotics for weaning turbot, Psetta maxima (Linnaeus, C. 1758). Aquac. Int. 2006, 14, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Grisdale-Helland, B.; Helland, S.J.; Gatlin, D.M. The effects of dietary supplementation with mannanoligosaccharide, fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon (Salmo salar). Aquaculture 2008, 283, 163–167. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Soleimani, N.; Ringø, E. Effects of dietary fructo-oligosaccharide supplementation on the growth performance, haemato-immunological parameters, gut microbiota and stress resistance of common carp (Cyprinus carpio) fry. Br.J. Nutr. 2014, 112, 1296–1302. [Google Scholar]
- Li, P.; Wen, Q.; Gatlin, D.M. Dose-dependent influences of dietary β-1, 3-glucan on innate immunity and disease resistance of hybrid striped bass Morone chrysops x Morone saxatilis. Aquac. Res. 2009, 40, 1578–1584. [Google Scholar]
- Zhang, Q.; Ma, H.; Mai, K.; Zhang, W.; Liufu, Z.; Xu, W. Interaction of dietary Bacillus subtilis and fructooligosaccharide on the growth performance, non-specific immunity of sea cucumber, Apostichopus japonicus. Fish Shellfish Immunol. 2010, 29, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Food Agriculture Organization (FAO). The State of Food Insecurity in the World; FAO: Rome, Italy; United Nations: New York, NY, USA, 2012. [Google Scholar]
- Sahoo, M.; Ceballos-Olvera, I.; del Barrio, L.; Re, F. Role of the Inflammasome, IL-1β, and IL-18 in Bacterial Infections. Sci. World J. 2011, 11, 2037–2050. [Google Scholar]
- Giri, S.S.; Sukumaran, V.; Oviya, M. Potential probiotic Lactobacillus plantarum VSG3 improves the growth, immunity, and disease resistance of tropical freshwater fish Labeo rohita. Fish Shellfish Immunol. 2013, 34, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Kumari, U.; Mittal, S.; Mittal, A.K. Immunoprotective role of aloin and disease resistance in Labeo rohita, infected with bacterial fish pathogen, Aeromonas hydrophila. Environ. Sci. Pollut. Res. 2023, 30, 30062–30072. [Google Scholar]
- Mukherjee, A.; Dutta, D.; Banerjee, S.; Ringø, E.; Breines, E.M.; Hareide, E.; Chandra, G.; Ghosh, K. Culturable autochthonous gut bacteria in rohu, Labeo rohita. In vitro growth inhibition against pathogenic Aeromonas spp., stability in gut, bio-safety and identification by 16S rRNA gene sequencing. Symbiosis 2017, 73, 165–177. [Google Scholar] [CrossRef]
- Sugita, H.; Hirose, Y.; Matsuo, N.; Deguchi, Y. Production of antibacterial substances by Bacillus sp. strain NM 12, an intestinal bacterium of Japanese coastal fish. Aquaculture 1998, 165, 269–280. [Google Scholar] [CrossRef]
- Sun, Y.Z.; Yang, H.L.; Ma, R.L.; Lin, W.Y. Probiotic applications of two dominant gut Bacillus strains with antagonistic activity improved the growth performance and immune responses of grouper Epinephelus coioides. Fish Shellfish Immunol. 2010, 29, 803–809. [Google Scholar] [CrossRef]
- Standard Methods for the Examination of Water and Wastewater, 22nd ed.; Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. (Eds.) American Public Health Association (APHA), American Water Works Association, Water Environment Federation: Washington, DC, USA, 2012. [Google Scholar]
- Official Methods of Analysis of the Association of Official Analytical Chemists, 19th ed.; Association of Official Analytical Chemists (AOAC): Washington, DC, USA, 2012.
- Maynard, L.A.; Loosli, J.K.; Hintz, H.F.; Warner, R.G. Animal Nutrition, 7th ed.; McGraw Hill: New York, NY, USA, 1979; pp. 13–14. [Google Scholar]
- Steffens, W. Principles of Fish Nutrition; Ellis Horwood Ltd.: Chichester, UK, 1989. [Google Scholar]
- Bernfeld, P. Amylase (alpha) and (beta). In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; pp. 149–150. [Google Scholar]
- Walter, H.E. Methods of Enzymatic Analysis; Verlag Chemie: Weinheim, Germany, 1984; pp. 270–277. [Google Scholar]
- Bier, M. Lipases. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; pp. 627–642. [Google Scholar]
- Lowry, O.H.; Rosenbrough, W.J.; Fair, H.L.; Randall, R.J. Protein measurement with Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Khan, K.U.; Zuberi, A.; Ullah, I.; Shami, S.A. Effects of graded level of dietary L-ascorbyl-2-polyphosphate on growth performance and some hematological indices of juvenile mahseer (Tor putitora). Int. J. Agric. Biol. 2015, 17, 821–827. [Google Scholar] [CrossRef]
- Ai, Q.; Mai, K.; Tan, B.; Xu, W.; Zhang, W.; Ma, H.; Liufu, Z. Effects of dietary vitamin C on survival, growth, and immunity of large yellow croaker, Pseudosciaena crocea. Aquaculture 2006, 261, 327–336. [Google Scholar] [CrossRef]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzymeactivities of naive atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Org. 2000, 41, 43–51. [Google Scholar] [CrossRef]
- Secombes, C.J. Isolation of salmonid macrophages and analysis of their killing activity. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., van-Muiswinkel, W.B., Eds.; SOS Publications: Fair Haven, CT, USA, 1990; pp. 137–155. [Google Scholar]
- Geng, X.; Dong, X.H.; Tan, B.P.; Yang, Q.H.; Chi, S.Y.; Liu, H.Y. Effects of dietary probiotic on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Aquac. Nutr. 2012, 18, 46–55. [Google Scholar] [CrossRef]
- Al-Dohail, A.M.; Hashim, R.; Aliyu-Paiko, M. Effects of the probiotic, Lactobacillus acidophilus, on the growth performance, haematology parameters and immunoglobulin concentration in African catfish (Clarias gariepinus, Burchell 1822) fingerling. Aquac. Res. 2009, 40, 1642–1652. [Google Scholar] [CrossRef]
- Johnson, C.W.; Timmons, D.L.; Hall, P.E.; Kuby, P. Essential Laboratory Mathematics: Concepts and Applications for the Chemical and Clinical Laboratory Technician, 2nd ed.; Delmar Cengage Learning: New York, NY, USA, 2002; p. 268. [Google Scholar]
- Blaxhall, P.C.; Daisley, K.W. Routine haematological methods for use with fish blood. J. Fish Biol. 1973, 5, 771–781. [Google Scholar]
- Wedemeyer, G.A.; Gould, R.W.; Yasutake, W.T. Some potentials and limits of the leucocrit test as fish health assessment method. J. Fish Biol. 1983, 23, 711–716. [Google Scholar] [CrossRef]
- Andani, H.R.R.; Tukmechi, A.; Meshkini, S.; Sheikhzadeh, N. Antagonistic activity of two potential probiotic bacteria from fish intestines and investigation of their effects on growth performance and immune response in rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2012, 28, 728–734. [Google Scholar] [CrossRef]
- Heo, W.-S.; Kim, Y.-R.; Kim, E.-Y.; Bai, S.C.; Kong, I.-S. Effects of dietary probiotic, Lactococcus lactis subsp. lactis I2, supplementation on the growth and immune response of olive flounder (Paralichthys olivaceus). Aquaculture 2013, 376–379, 20–24. [Google Scholar] [CrossRef]
- Zuo, X.; Woo, P.T.K. Natural antiproteases in rainbow trout. Oncorhynchus mykiss and brook charr. Salvelinus fontinalis and the in vitro neutralization of fish a2-macroglobulin by the metalloprotease from the pathogenic haemoflagellate, Cryptobia salmositica. Parasitology 1997, 114, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.P.; Chang, C.A.; Chang, C.Y.; Liu, C.H.; Cheng, W. Dietary sodium alginate administration affects the fingerling growth and resistance to Streptococcus sp. And iridovirus, and juvenile non-specific immune responses of the orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2008, 25, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Kim, M.C.; Kim, J.S.; Balasundaram, C.; Heo, M.S. Immunomodulatory effect of probiotics enriched diets on Uronema marinum infected olive flounder. Fish Shellfish Immunol. 2011, 30, 964–971. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulphate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxidation in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Education Singapore Pte. Ltd: New Delhi, India, 2010. [Google Scholar]
- Studio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 6 December 2022).
- Cerezuela, R.; Meseguer, J.; Esteban, M.A. Current Knowledge in Synbiotic Use for Fish Aquaculture: A Review. J. Aquac. Res. Dev. 2011, 1, 1–7. [Google Scholar]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subedi, B.; Shrestha, A. A review: Application of probiotics in aquaculture. Int. J. For. Anim. Fish. Res. 2020, 4, 52–60. [Google Scholar]
- Rohani, M.F.; Islam, S.M.; Hossain, M.K.; Ferdous, Z.; Siddik, M.A.; Nuruzzaman, M.; Padeniya, U.; Brown, C.; Shahjahan, M. Probiotics, prebiotics and synbiotics improved the functionality of aquafeed: Upgrading growth, reproduction, immunity and disease resistance in fish. Fish Shellfish Immunol. 2022, 120, 569–589. [Google Scholar] [CrossRef]
- Wee, W.; Hamid, N.K.A.; Mat, K.; Khalif, R.I.A.R.; Rusli, N.D.; Rahman, M.M.; Kabir, M.A.; Wei, L.S. The effects of mixed prebiotics in aquaculture: A review. Aquac. Fish. 2022, in press. [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Rodiles, A.; Merrifield, D.L.; Chandra, G.; Ghosh, K. Exploring intestinal microbiome composition in three Indian major carps under polyculture system: A high-throughput sequencing based approach. Aquaculture 2020, 524, 735206. [Google Scholar] [CrossRef]
- Li, Z.; Tran, N.T.; Ji, P.; Sun, Z.; Wen, X.; Li, S. Effects of prebiotic mixtures on growth performance, intestinal microbiota and immune response in juvenile chu’s croaker, Nibea coibor. Fish Shellfish Immunol. 2019, 89, 564–573. [Google Scholar] [CrossRef]
- Kapiki, A.; Costalos, C.; Oikonomidou, C.; Triantafyllidou, A.; Loukatou, E.; Pertrohilou, V. The effect of a fructo-oligosaccharide supplemented formula on gut flora of preterm infants. Early Hum. Dev. 2007, 83, 335–339. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, H.; Tong, T.; Tong, W.; Dong, L.; Xu, M.; Wang, Z. Dietary supplementation of Bacillus subtilis and fructooligosaccharide enhance the growth, non-specific immunity of juvenile ovate pompano, Trachinotus ovatus and its disease resistance against Vibrio vulnificus. Fish Shellfish Immunol. 2014, 38, 7–14. [Google Scholar] [CrossRef]
- Mehrabi, Z.; Firouzbakhsh, F.; Jafarpour, A. Effects of dietary supplementation of synbiotic on growth performance, serum biochemical parameters and carcass composition in rainbow trout (Oncorhynchus mykiss) fingerlings. J. Anim. Physiol. Anim. Nutr. 2012, 96, 474–481. [Google Scholar] [CrossRef]
- Torrecillas, S.; Rivero-Ramírez, F.; Izquierdo, M.S.; Caballero, M.J.; Makol, A.; Suarez-Bregua, P.; Fernández-Montero, A.; Rotllant, J.; Montero, D. Feeding European sea bass (Dicentrarchus labrax) juveniles with a functional synbiotic additive (mannan oligosaccharides and Pediococcus acidilactici): An effective tool to reduce low fishmeal and fish oil gut health effects? Fish Shellfish Immunol. 2018, 81, 10–20. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Soltan, M.A.; Ghonemy, M.M.R. Effect of synbiotics between Bacillus licheniformis and yeast extract on growth, hematological and biochemical indices of the Nile tilapia (Oreochromis niloticus). Egypt. J. Aquat. Res. 2014, 40, 199–208. [Google Scholar]
- Dawood, M.A.O.; Eweedah, N.M.; Moustafa, E.M.; Shahin, M.G. Synbiotic Effects of Aspergillus oryzae and β-Glucan on Growth and Oxidative and Immune Responses of Nile Tilapia, Oreochromis niloticus. Probiotics Antimicrob. Proteins 2020, 12, 172–183. [Google Scholar] [CrossRef]
- Naseri, S.; Khara, H.; Shakoori, M. Effects of probiotics and Fe ion on the growth and survival and body composition of rainbow trout, Oncorhynchus mykiss (Walbaum, 1792) frys. J. Appl. Anim. Res. 2013, 41, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Azarin, H.; Aramli, M.S.; Imanpour, M.R.; Rajabpour, M. Effect of a Probiotic Containing Bacillus licheniformis and Bacillus subtilis and Ferroin Solution on Growth Performance, Body Composition and Haematological Parameters in Kutum (Rutilus frisii kutum) fry. Probiotics Antimicrob. Proteins 2015, 7, 31–37. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Dadar, M.; Ringø, E. Modulation of nutrient digestibility and digestive enzyme activities in aquatic animals: The functional feed additives scenario. Aquac. Res. 2017, 48, 3987–4000. [Google Scholar] [CrossRef]
- Ashouri, G.; Soofiani, N.M.; Hoseinifar, S.H.; Jalali, S.A.H.; Morshedi, V.; Valinassab, T.; Bagheri, D.; van Doan, H.; Mozanzadeh, M.T.; Carnevali, O. Influence of dietary sodium alginate and Pediococcus acidilactici on liver antioxidant status, intestinal lysozyme gene expression, histomorphology, microbiota, and digestive enzymes activity, in Asian sea bass (Lates calcarifer) juveniles. Aquaculture 2020, 518, 734638. [Google Scholar] [CrossRef]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. Probiotics in marine larviculture. FEMS Microbiol. Rev. 2006, 30, 404–427. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Chakraborty, T.; Prusty, A.K.; Das, P.; Paniprasad, K.; Mohanta, K.N. Use of different microbial probiotics in the diet of rohu, Labeo rohita fingerlings: Effects on growth, nutrient digestibility and retention, digestive enzyme activities and intestinal microflora. Aquac. Nutr. 2012, 18, 1–11. [Google Scholar] [CrossRef]
- Ye, J.-D.; Wang, K.; Li, F.-D.; Sun, Y.-Z. Single or combined effects of fructo- and mannan oligosaccharide supplements and Bacillus clausii on the growth, feed utilization, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquac. Nutr. 2011, 17, 902–911. [Google Scholar]
- Zhang, C.-N.; Li, X.-F.; Xu, W.-N.; Zhang, D.-D.; Lu, K.-L.; Wang, L.-N.; Tian, H.-Y.; Liu, W.-B. Combined effects of dietary fructooligosaccharide and Bacillus licheniformis on growth performance, body composition, intestinal enzymes activities and gut histology of triangular bream (Megalobrama terminalis). Aquac. Nutr. 2015, 21, 755–766. [Google Scholar] [CrossRef]
- Nayak, S.K.; Swain, P.; Mukherjee, S.C. Effect of dietary supplementation of probiotic and vitamin C on the immune response of Indian major carp, Labeo rohita (Ham.). Fish Shellfish Immunol. 2007, 23, 892–896. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sadegh, T.H.; Dawood, M.A.O.; Dadar, M.; Sheikhzadeh, N. The effects of dietary Pediococcus pentosaceus on growth performance, hemato-immunological parameters and digestive enzyme activities of common carp (Cyprinus carpio). Aquaculture 2020, 516, 734656. [Google Scholar] [CrossRef]
- John, P.J. Alteration of certain blood parameters of fresh water teleost Mystus vittatus after chronic exposure to Metasystox and Sevin. Fish Physiol. Biochem. 2007, 33, 15–20. [Google Scholar] [CrossRef]
- Ranzani-Paiva, M.J.T.; Ishikawa, C.M.; das Eiras, A.C.; da Silveira, V.R. Effects of an Experimental Challenge with Mycobacterium marinum on the Blood Parameters of Nile Tilapia, Oreochromis niloticus (Linnaeus, 1757). Braz. Arch. Biol. Technol. 2004, 47, 945–953. [Google Scholar] [CrossRef]
- Haniffa, M.A.; Abdul, K.M.K.P. Hematological Changes in Channa striatus Experimentally infected by Aeromonas hydrophila. Bioresearch Bulletin 2011, 4, 246–253. [Google Scholar]
- Firouzbakhsh, F.; Noori, F.; Khalesi, M.K.; Jani-Khalili, K. Effects of a probiotic, protexin, on the growth performance and hematological parameters in the Oscar (Astronotus ocellatus) fingerlings. Fish Physiol. Biochem. 2011, 37, 833–842. [Google Scholar] [CrossRef]
- Reda, R.M.; Selim, K. Evaluation of Bacillus amyloliquefaciens on the growth performance, intestinal morphology, hematology and body composition of Nile tilapia, Oreochromis niloticus. Aquac. Int. 2015, 23, 203–217. [Google Scholar] [CrossRef]
- Elsabagh, M.; Mohamed, R.; Moustafa, E.M.; Hamza, A.; Farrag, F.; Decamp, O.; Dawood, M.A.O.; Eltholth, M. Assessing the impact of Bacillus strains mixture probiotic on water quality, growth performance, blood profile and intestinal morphology of Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2018, 24, 1613–1622. [Google Scholar] [CrossRef]
- Aftabgard, M.; Salarzadeh, A.; Mohseni, M.; Shabanipour, A.H.B.; Zorriehzahra, M.E.J. The Combined Efficiency of Dietary Isomaltooligosaccharides and Bacillus spp. on the Growth, Hemato-Serological, and Intestinal Microbiota Indices of Caspian Brown Trout (Salmo trutta caspius Kessler, 1877). Probiotics Antimicrob. Proteins 2019, 11, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Mirvaghefi, A.; Merrifield, D.L.; Amiri, B.M.; Yelghi, S.; Bastami, K.D. The study of some haematological and serum biochemical parameters of juvenile beluga (Huso huso) fed oligofructose. Fish Physiol. Biochem. 2011, 37, 91–96. [Google Scholar] [CrossRef]
- Mansour, M.R.; Akrami, R.; Ghobadi, S.H.; Denji, K.A.; Ezatrahimi, N.; Gharaei, A. Effect of dietary mannan oligosaccharide (MOS) on growth performance, survival, body composition, and some hematological parameters in giant sturgeon juvenile (Huso huso Linnaeus, 1754). Fish Physiol. Biochem. 2012, 38, 829–835. [Google Scholar] [CrossRef]
- Rehulka, J.; Minarik, B.; Adamec, V.; Rehulkova, E. Investigations of physiological and pathological levels of total plasma protein in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Res. 2005, 36, 22–32. [Google Scholar] [CrossRef]
- Zeinab, A.K.; Aly, M.S.; Faiza, A.; Fatma, E.M. Effect of Spirulina platensis and Lactobacillus rhamnosus of Nile Tilapia (Oreochromis niloticus) fingerlings. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 747–763. [Google Scholar]
- Toutou, M.M.; Soliman, A.A.A.; Farrag, M.M.S. Effect of Probiotic and Synbiotic Food Supplementation on Growth Performance and Healthy Status of Grass Carp, Ctenopharyngodon idella (Valenciennes,1844). Int. J. Ecotoxicol. Ecobiol. 2016, 1, 111–117. [Google Scholar]
- Dhanalakshmi, P.; Ramasubramanian, V. Effect of Probiotics and Prebiotics Supplemented Diets on Immune Resistance against Aeromonas hydrophila in Mrigal Carp. Ann. Aquac. Res. 2017, 4, 1042. [Google Scholar]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and Non-alcoholic Fatty Liver Disease: Biochemical, Metabolic, and Clinical Implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panigrahi, A.; Kiron, V.; Satoh, S.; Watanabe, T. Probiotic bacteria Lactobacillus rhamnosus influences the blood profile in rainbow trout Oncorhynchus mykiss (Walbaum). Fish Physiol. Biochem. 2010, 36, 969–977. [Google Scholar] [CrossRef]
- Lye, H.S.; Rusul, G.; Liong, M.T. Removal of cholesterol by lactobacilli via incorporation and conversion to coprostanol. J. Dairy Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Talpur, A.D.; Ikhwanuddin, M. Dietary effects of garlic (Allium sativum) on haemato-immunological parameters, survival, growth, and disease resistance against Vibrio harveyi infection in Asian seabass, Lates calcarifer (Bloch). Aquaculture 2012, 364–365, 6–12. [Google Scholar] [CrossRef]
- Park, S.; Ji, Y.; Park, H.; Lee, K.; Park, H.; Beck, B.R.; Shin, H.; Holzapfel, W.H. Evaluation of functional properties of lactobacilli isolated from Korean white kimchi. Food Control 2016, 69, 5–12. [Google Scholar] [CrossRef]
- Kopel, E.; Kivity, S.; Morag-Koren, N.; Segev, S.; Sidi, Y. Relation of serum lactate dehydrogenase to coronary artery disease. Am. J. Cardiol. 2012, 110, 1717–1722. [Google Scholar] [CrossRef]
- Lara-Flores, M.; Olvera-Novoa, M.A.; Guzman-Mendez, B.E.; Lopez-Madrid, W. Use of the bacteria Streptococcus faecium and Lactobacillus acidophilus, and the yeast Saccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromis niloticus). Aquaculture 2003, 216, 193–201. [Google Scholar] [CrossRef]
- Alexander, J.B.; Ingram, G.A. Noncellular nonspecific defence mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 249–279. [Google Scholar] [CrossRef]
- Magnado’ttir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Staykov, Y.; Spring, P.; Denev, S.; Sweetman, J. Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2007, 15, 153–161. [Google Scholar] [CrossRef]
- Buentello, J.A.; Neill, W.H.; Gatlin, D.M. Effects of dietary prebiotics on the growth, feed efficiency and non-specific immunity of juvenile red drum Sciaenops ocellatus fed soybean-based diets. Aquac. Res. 2010, 41, 411–418. [Google Scholar] [CrossRef]
- Akrami, R.; Nasri-Tajan, M.; Jahedi, A.; Jahedi, M.; Mansour, M.R.; Jafarpour, S.A. Effects of dietary synbiotic on growth, survival, lactobacillus bacterial count, blood indices and immunity of beluga (Huso huso Linnaeus, 1754) juvenile. Aquac. Nutr. 2015, 21, 952–959. [Google Scholar] [CrossRef]
- Esteban, M.A.; Cerezuela, R. Fish mucosal immunity. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 67–92. [Google Scholar]
- Modanloo, M.; Soltanian, S.; Akhlaghi, M.; Hoseinifar, S.H. The effects of single or combined administration of galactooligosaccharide and Pediococcus acidilactici on cutaneous mucus immune parameters, humoral immune responses and immune related genes expression in common carp (Cyprinus carpio) fingerlings. Fish Shellfish Immunol. 2017, 70, 391–397. [Google Scholar] [CrossRef]
- Ostrycharz, E.; Hukowska-Szematowicz, B. New Insights into the Role of the Complement System in Human Viral Diseases. Biomolecules 2022, 12, 226. [Google Scholar] [CrossRef]
- Chitsaz, H.; Akrami, R.; Arab, A.M. Effect of dietary synbiotics on growth, immune response and body composition of Caspian roach (Rutilus rutilus). Iran. J. Fish. Sci. 2016, 15, 170–182. [Google Scholar]
- Sheikhvaisy, F.; Safari, O.; Vakili, R. Modulation of growth performance, haemato-immunological parameters, gut microbiota and stress resistance upon feeding juvenile Schizothorax zarudnyi (Nikolskii, 1897) by fructo-oligosaccharid. Iran. J. Vet. Sci. Technol. 2017, 9, 46–56. [Google Scholar]
- Wang, Y.B.; Tian, Z.Q.; Yao, J.T.; Li, W. Effect of probiotics, Enterococcus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture 2008, 277, 203–207. [Google Scholar] [CrossRef]
- van Doan, H.; Hoseinifar, S.H.; Tapingkae, W.; Seel-audom, M.; Jaturasitha, S.; Dawood, M.A.O.; Wongmaneeprateep, S.; Thu, T.T.N.; Esteban, M.Á. Boosted Growth Performance, Mucosal and Serum Immunity, and Disease Resistance Nile Tilapia (Oreochromis niloticus) Fingerlings Using Corncob-Derived Xylooligosaccharide and Lactobacillus plantarum CR1T5. Probiotics Antimicrob. Proteins 2020, 12, 400–411. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; El-Sabagh, M.; Billah, M.M.; Zaineldin, A.I.; Zayed, M.M.; Omar, A.A.E.D. Changes in the growth, humoral and mucosal immune responses following β-glucan and vitamin C administration in red sea bream, Pagrus major. Aquaculture 2017, 470, 214–222. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Austin, B. Influence of probiotic feeding duration on disease resistance and immune parameters in rainbow trout. Fish Shellfish Immunol. 2009, 27, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Chiu, C.-H.; Wang, S.-W.; Cheng, W. Dietary administration of the probiotic, Bacillus subtilis E20, enhances the growth, innate immune responses, and disease resistance of the grouper, Epinephelus coioides. Fish Shellfish Immunol. 2012, 33, 699–706. [Google Scholar] [CrossRef]
- Ramesh, D.; Souissi, S.; Ahamed, T.S. Effects of the potential probiotics Bacillus aerophilus KADR3 in inducing immunity and disease resistance in Labeo rohita. Fish Shellfish Immunol. 2017, 70, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.; Mohapatra, P.K.D. Effect of a probiotic bacterium Bacillus circulans PB7 in the formulated diets: On growth, nutritional quality and immunity of Catla catla (Ham.). Fish Physiol. Biochem. 2009, 35, 467–478. [Google Scholar] [CrossRef]
- Forman, H.J.; Torres, M. Reactive oxygen species and cell signaling: Respiratory burst in macrophage signalling. Am. J. Respir.Crit. Care Med. 2002, 166, 4–8. [Google Scholar] [CrossRef]
- Kumar, P.; Jain, K.K.; Sardar, P. Effects of dietary synbiotic on innate immunity, antioxidant activity and disease resistance of Cirrhinus mrigala juveniles. Fish Shellfish Immunol. 2018, 80, 124–132. [Google Scholar] [CrossRef]
- Chiu, C.-H.; Cheng, C.-H.; Gua, W.-R.; Guu, Y.-K.; Cheng, W. Dietary administration of the probiotic, Saccharomyces cerevisiae P13, enhanced the growth, innate immune responses and disease resistance of the grouper, Epinephelus coioides. Fish Shellfish Immunol. 2010, 29, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Chanagun, C.; Thongsri, S.; Phimpimon, T. The Influences of Dietary Supplementation with Fructooligosaccharide on Growth and Immune Responses of Climbing Perch (Anabas testudineus). Int. J.Agric. Biol. 2017, 19, 787–791. [Google Scholar]
- Sahu, S.; Das, B.K.; Mishra, B.K.; Pradhan, J.; Sarangi, N. Effects of Allium sativum on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. J. Appl. Ichthyol. 2007, 23, 80–86. [Google Scholar] [CrossRef]
- Piner, P.; Uner, N. Oxidative Stress and apoptosis was induced by bio-insecticide spinosad in the liver of Orechromis niloticus. Environ.Toxicol. Pharmacol. 2013, 36, 956–963. [Google Scholar] [CrossRef]
- Ferreira, M.; Moradas-Ferreira, P.; Reis-Henriques, M.A. The effect of long-term depuration on levels of oxidative stress biomarkers in mullets (Mugil cephalus) chronically exposed to contaminants. Mar. Environ. Res. 2007, 64, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Kohen, R.; Nyska, A. Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, Y.-J.; Tian, L.-X.; Yang, H.-J.; Liang, G.-Y.; Yue, Y.-R.; Xu, D.-H. Effects of dietary astaxanthin on growth, antioxidant capacity and gene expression in Pacific white shrimp Litopenaeus vannamei. Aquac Nutr. 2013, 19, 917–927. [Google Scholar] [CrossRef]
- Nandi, A.; Banerjee, G.; Dan, S.K.; Ghosh, K.; Ray, A.K. Probiotic efficiency of Bacillus sp. in Labeo rohita challenged by Aeromonas hydrophila: Assessment of stress profile, haemato-biochemical parameters and immune responses. Aquac. Res. 2017, 48, 4334–4345. [Google Scholar] [CrossRef]
- Wongsasak, U.; Chaijamrus, S.; Kumkhong, S.; Boonanuntanasarn, S. Effects of dietary supplementation with β-glucanand synbiotics on immune gene expression and immune parameters under ammonia stress in Pacific white shrimp. Aquaculture 2015, 436, 179–187. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K.; Pandey, S.; Kumar, R. An alternative approach of toxic heavy metal removal by Arthrobacter phenanthrenivorans: Assessment of surfactant production and oxidative stress. Curr. Sci. 2016, 110, 2124–2128. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A. Increases in immune parameters by inulin and Bacillus subtilis dietary administration to gilthead seabream (Sparus aurata L.) did not correlate with disease resistance to Photobacterium damselae. Fish Shellfish Immunol. 2012, 32, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.M.; Ahmed, Y.A.G.; Ghareeb, A.A.A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient Profile (on % Dry Weight) | Proximate Composition | ||
|---|---|---|---|
| Fish meal | 37 | Dry matter | 92.32 ± 0.49 |
| Mustard oil cake | 25 | Crude protein | 35.27 ± 0.20 |
| Rice bran | 15 | Crude lipid | 6.71 ± 0.03 |
| Wheat bran | 20 | Crude fibre | 13.66 ± 0.14 |
| Cod liver oil | 1 | Nitrogen-free extract | 21.49 ± 0.12 |
| Vitamin + mineral premix a | 1 | Ash | 14.69 ± 0.07 |
| Carboxymethylcellulose | 1 | Gross energy (MJkg−1) | 17.06 ± 0.09 |
| Experimental Groups | Diets Used | FOS | B. licheniformis (KU556167) | B. methylotrophicus (KU556164) |
|---|---|---|---|---|
| C | Basal | - | - | - |
| E1 | Basal + Prebiotic | 1% | - | - |
| E2 | Basal + Probiotic | - | 7 Log CFU/g | - |
| E3 | Basal + Probiotic | - | - | 7 Log CFU/g |
| E4 | Basal + Synbiotic | 1% | 7 Log CFU/g | - |
| E5 | Basal + Synbiotic | 1% | - | 7 Log CFU/g |
| Parameters | C | E1 | E2 | E3 | E4 | E5 |
|---|---|---|---|---|---|---|
| Initial weight (g) | 10.21 ± 0.17 | 10.15 ± 0.15 | 10.1 ± 0.17 | 10.17 ± 0.16 | 10.19 ± 0.17 | 10.25 ± 0.16 |
| Final weight (g) | 29.97 ± 0.31 a | 31.64 ± 0.29 b | 33.87 ± 0.32 c | 34.07 ± 0.32 c | 36.66 ± 0.40 d | 35.39 ± 0.25 c |
| Weight gain (%) | 193.53 ± 2.03 a | 211.72 ± 1.92 b | 235.34 ± 2.26 c | 234.7 ± 2.21 c | 259.72 ± 2.83 d | 245.27 ± 1.77 c |
| SGR (% day−1) | 1.19 ± 0.02 a | 1.26 ± 0.01 a | 1.34 ± 0.02 b | 1.34 ± 0.01 b | 1.42 ± 0.02 c | 1.37 ± 0.01 bc |
| FCR 1 | 2.4 ± 0.01 e | 2.22 ± 0.01 d | 2.0 ± 0.01 c | 2.02 ± 0.02 c | 1.74 ± 0.01 a | 1.84 ± 0.01 b |
| PER 2 | 1.19 ± 0.02 a | 1.29 ± 0.02 b | 1.43 ± 0.01 c | 1.41 ± 0.01 c | 1.64 ± 0.02 e | 1.55 ± 0.02 d |
| ANPU 3 (%) | 17.98 ± 0.27 a | 20.57 ± 0.33 b | 24.53 ± 0.34 c | 23.47 ± 0.25 c | 29.58 ± 0.31 e | 27.19 ± 0.40 d |
| Survivability (%) | 93.33 ± 1.92 | 96.66 ± 0.00 | 96.66 ± 1.92 | 93.33 ± 3.9 | 96.66 ± 1.92 | 96.66 ± 0.00 |
| Condition factor (K) | 0.92 ± 0.01 a | 1.1 ± 0.01 b | 1.2 ± 0.01 c | 1.16 ± 0.01 c | 1.52 ± 0.01 e | 1.43 ± 0.01 d |
| After 90 Days | ||||||
|---|---|---|---|---|---|---|
| Parameters | C | E1 | E2 | E3 | E4 | E5 |
| RBC (×106/mm3) | 2.77 ± 0.01 a | 2.77 ± 0.02 a | 2.85 ± 0.02 ab | 2.78 ± 0.01 ab | 3.05 ± 0.02 c | 2.86 ± 0.02 b |
| WBC (×102/mm3) | 180 ± 0.88 a | 198 ± 0.93 b | 204 ± 0.97 c | 212 ± 1.02 d | 221 ± 0.97 e | 221 ± 1.24 e |
| Hb (g/dL) | 10.7 ± 0.11 ab | 10.4 ± 0.13 a | 11.3 ± 0.12 bc | 11.0 ± 0.14 ab | 11.8 ± 0.16 c | 11.0 ± 0.11 ab |
| PCV (%) | 40.0 ± 0.21 a | 40.1 ± 0.21 a | 43.3 ± 0.19 c | 43.3 ± 0.22 c | 45.6 ± 0.22 d | 41.9 ± 0.18 b |
| ESR (mm/h) | 3.8 ± 0.02 c | 3.3 ± 0.02 b | 2.6 ± 0.02 a | 2.5 ± 0.01 a | 2.6 ± 0.02 a | 2.5 ± 0.01 a |
| Platelet (×103/mm3) | 39 ± 0.42 b | 36 ± 0.75 a | 60 ± 0.73 f | 45 ± 0.67 d | 56 ± 0.19 e | 42 ± 0.16 c |
| MCV(μm3) | 144.4 ± 0.76 ab | 141.15 ± 0.71 a | 151.9 ± 0.83 d | 155.8 ± 0.93 e | 149.5 ± 0.79 cd | 146.5 ± 0.72 bc |
| MCH (pg/cell) | 38.62 ± 0.21 bc | 37.54 ± 0.18 a | 39.6 ± 0.20 c | 39.6 ± 0.18 c | 38.7 ± 0.24 bc | 38.5 ± 0.30 ab |
| MCHC (g%) | 26.8 ± 0.11 c | 25.9 ± 0.13 ab | 26.1 ± 0.13 b | 25.4 ± 0.15 a | 25.9 ± 0.15 ab | 26.3 ± 0.13 bc |
| After challenge with Aeromonas hydrophila | ||||||
| RBC (×106/mm3) | 1.97 ± 0.01 bc | 1.86 ± 0.01 a | 1.92 ± 0.01 b | 1.98 ± 0.01 c | 2.12 ± 0.01 d | 2.02 ± 0.01 c |
| WBC (×102/mm3) | 252 ± 1.43 a | 254 ± 1.23 ab | 262 ± 1.33 cd | 258 ± 1.28 bc | 264 ± 1.20 d | 257 ± 0.97 ab |
| Hb (g/dL) | 7.2 ± 0.07 a | 7.9 ± 0.09 b | 8.1 ± 0.09 b | 7.9 ± 0.08 b | 8.9 ± 0.11 c | 8.0 ± 0.10 b |
| PCV (%) | 33.3 ± 0.14 a | 33.1 ± 0.20 a | 34.5 ± 0.15 bc | 33.5 ± 0.18 ab | 36.5 ± 0.20 d | 34.8 ± 0.21 c |
| ESR (mm/h) | 4.7 ± 0.02 d | 3.7 ± 0.02 b | 3.8 ± 0.02 c | 3.8 ± 0.02 c | 3.3 ± 0.02 a | 3.7 ± 0.02 b |
| Platelet (×103/mm3) | 35 ± 0.43 a | 37 ± 0.20 b | 52 ± 0.40 f | 43 ± 0.31 d | 50 ± 0.35 e | 41 ± 0.23 c |
| MCV (μm3) | 169.13 ± 1.09 a | 178.06 ± 0.95 b | 179.89 ± 0.82 b | 169.39 ± 0.96 a | 171.93 ± 0.82 a | 172.07 ± 0.87 a |
| MCH (pg/cell) | 36.59 ± 0.30 a | 42.47 ± 0.16 c | 42.29 ± 0.40 c | 39.74 ± 0.36 b | 41.83 ± 0.37 c | 39.7 ± 0.29 b |
| MCHC (g%) | 21.63 ± 0.11 a | 23.85 ± 0.10 cd | 23.5 ± 0.13 bc | 23.46 ± 0.13 bc | 24.33 ± 0.14 d | 23.07 ± 0.12 b |
| After 90 Days | ||||||
|---|---|---|---|---|---|---|
| Parameters | C | E1 | E2 | E3 | E4 | E5 |
| Total protein (g dL−1) | 2.71 ± 0.02 a | 2.86 ± 0.02 b | 3.12 ± 0.01 c | 3.07 ± 0.03 c | 3.41 ± 0.02 d | 3.16 ± 0.02 c |
| Albumin (g dL−1) | 1.30 ± 0.01 a | 1.37 ± 0.01 b | 1.51 ± 0.01 c | 1.50 ± 0.004 c | 1.66 ± 0.01 d | 1.53 ± 0.01 c |
| Globulin (g dL−1) | 1.41 ± 0.02 a | 1.49 ± 0.01 b | 1.61 ± 0.02 c | 1.57 ± 0.02 c | 1.75 ± 0.01 d | 1.63 ± 0.01 c |
| Cholesterol (mg dL−1) | 128 ± 2.12 a | 133 ± 2.01 a | 145 ± 1.36 b | 142 ± 0.46 b | 150 ± 2.23 bc | 155 ± 1.49 c |
| HDL (mg dL−1) | 31 ± 0.33 a | 50 ± 0.18 b | 55 ± 0.26 c | 55 ± 0.18 c | 60 ± 0.20 d | 61 ± 0.23 d |
| LDL (mg dL−1) | 78 ± 0.84 b | 72 ± 0.26 a | 81 ± 0.38 c | 78 ± 0.25 b | 84 ± 0.28 d | 85 ± 0.32 d |
| Triglyceride (mg dL−1) | 95 ± 0.47 d | 55 ± 0.35 c | 45 ± 0.25 b | 45 ± 0.22 b | 30 ± 0.23 a | 45 ± 0.29 b |
| Glucose (mg dL−1) | 127.4 ± 0.63 d | 125.5 ± 0.60 cd | 121.5 ± 0.68 ab | 123 ± 0.78 bc | 118 ± 0.89 a | 118 ± 1.19 a |
| ALP (µ cat L−1) | 0.55 ± 0.002 a | 0.73 ± 0.002 c | 0.72 ± 0.002 c | 0.62 ± 0.002 b | 1.15 ± 0.004 e | 0.77 ± 0.003 d |
| SGPT (µ cat L−1) | 1.85 ± 0.02 c | 1.72 ± 0.02 b | 1.59 ± 0.02 a | 1.55 ± 0.02 a | 1.57 ± 0.02 a | 1.52 ± 0.02 a |
| SGOT (µ cat L−1) | 1.46 ± 0.03 b | 1.45 ± 0.02 b | 1.23 ± 0.02 a | 1.41 ± 0.02 b | 1.22 ± 0.01 a | 1.39 ± 0.01 b |
| LDH (µ cat L−1) | 6.26 ± 0.07 b | 6.24 ± 0.02 b | 6.04 ± 0.03 a | 6.1 ± 0.02 ab | 6.01 ± 0.02 a | 6.12 ± 0.02 ab |
| After challenge with Aeromonas hydrophila | ||||||
| Total protein (g dL−1) | 2.61 ± 0.01 a | 2.78 ± 0.03 b | 3.27 ± 0.03 c | 3.35 ± 0.02 c | 3.58 ± 0.02 d | 3.6 ± 0.02 d |
| Albumin (g dL−1) | 1.18 ± 0.01 a | 1.24 ± 0.01 b | 1.43 ± 0.01 c | 1.42 ± 0.01 c | 1.58 ± 0.01 e | 1.52 ± 0.01 d |
| Globulin (g dL−1) | 1.43 ± 0.02 a | 1.54 ± 0.01 b | 1.84 ± 0.01 c | 1.93 ± 0.01 d | 2.00 ± 0.01 e | 2.08 ± 0.01 f |
| Cholesterol (mg dL−1) | 175 ± 0.76 c | 178 ± 0.84 c | 160 ± 1.71 a | 161 ± 1.69 ab | 165 ± 0.84 ab | 167 ± 1.08 b |
| HDL (mg dL−1) | 34 ± 0.19 a | 48 ± 0.26 b | 50 ± 0.25 c | 53 ± 0.31 e | 51 ± 0.26 cd | 52 ± 0.34 de |
| LDL (mg dL−1) | 115 ± 0.63 d | 106 ± 0.58 c | 95 ± 0.47 a | 93 ± 0.55 a | 98 ± 0.50 b | 98 ± 0.63 b |
| Triglyceride (mg dL−1) | 130 ± 0.74 e | 120 ± 0.93 d | 75 ± 0.38 a | 75 ± 0.37 a | 80 ± 0.50 b | 85 ± 0.54 c |
| Glucose (mg dL−1) | 142.7 ± 0.81 c | 141.4 ± 1.09 c | 134.86 ± 0.67 ab | 138.85 ± 0.69 bc | 134.1 ± 0.84 a | 138.76 ± 0.89 bc |
| ALP(µ cat L−1) | 0.87 ± 0.003 c | 0.85 ± 0.004 b | 0.87 ± 0.004 c | 0.79 ± 0.005 a | 1.23 ± 0.005 d | 0.86 ± 0.004 bc |
| SGPT (µ cat L−1) | 2.11 ± 0.02 c | 2.05 ± 0.02 c | 1.89 ± 0.02 b | 1.87 ± 0.01 b | 1.76 ± 0.02 a | 1.81 ± 0.02 ab |
| SGOT (µ cat L−1) | 1.87 ± 0.02 c | 1.84 ± 0.01 c | 1.36 ± 0.01 a | 1.49 ± 0.01 b | 1.34 ± 0.01 a | 1.44 ± 0.01 b |
| LDH (µ cat L−1) | 8.78 ± 0.05 b | 8.72 ± 0.05 b | 8.45 ± 0.05 a | 8.44 ± 0.04 a | 8.41 ± 0.05 a | 8.32 ± 0.05 a |
| After 90 Days | ||||||
|---|---|---|---|---|---|---|
| Parameters | C | E1 | E2 | E3 | E4 | E5 |
| Serum Lysozyme 1 | 14.52 ± 0.09 a | 16.42 ± 0.14 b | 18.78 ± 0.10 c | 18.75 ± 0.14 c | 21.23 ± 0.15 e | 19.87 ± 0.13 d |
| Mucus Lysozyme 2 | 3.03 ± 0.05 a | 3.12 ± 0.02 a | 3.93 ± 0.07 b | 3.97 ± 0.07 b | 5.05 ± 0.09 c | 4.1 ± 0.07 b |
| ACP 3 | 127.09 ± 0.82 a | 152.45 ± 1.26 b | 166.69 ± 0.90 c | 162 ± 1.20 c | 194.03 ± 1.41 e | 178.23 ± 1.19 d |
| Serum Peroxidase 4 | 0.52 ± 0.012 a | 0.68 ± 0.016 b | 0.94 ± 0.022 c | 1.05 ± 0.023 d | 1.13 ± 0.020 de | 1.17 ± 0.032 e |
| Mucus Peroxidase 5 | 0.13 ± 0.004 a | 0.19 ± 0.005 b | 0.26 ± 0.007 c | 0.29 ± 0.008 c | 0.35 ± 0.009 d | 0.37 ± 0.010 d |
| Antiprotease 6 | 72.71 ± 0.53 a | 72.96 ± 0.39 a | 78.36 ± 0.31 c | 76.17 ± 0.87 b | 80.98 ± 0.45 d | 84.27 ± 0.42 e |
| Phagocytosis 7 | 23.43 ± 0.25 a | 24.76 ± 0.52 a | 31.84 ± 0.39 b | 35.23 ± 0.52 c | 38.54 ± 0.13 d | 36.76 ± 0.14 c |
| Super oxide anion 8 | 38.12 ± 0.69 a | 39.65 ± 0.10 a | 43.23 ± 0.78 bc | 41.2 ± 0.74 ab | 46.54 ± 0.84 c | 45.65 ± 0.82 c |
| RB 9 | 0.22 ± 0.005 a | 0.23 ± 0.008 a | 0.34 ± 0.009 b | 0.34 ± 0.010 b | 0.42 ± 0.012 d | 0.39 ± 0.010 c |
| After challenge with Aeromonas hydrophila | ||||||
| Serum Lysozyme 1 | 17.45 ± 0.09 a | 19.85 ± 0.11 b | 22.63 ± 0.09 c | 23.57 ± 0.31 d | 25.77 ± 0.12 f | 24.61 ± 0.30 e |
| Mucus Lysozyme 2 | 3.84 ± 0.07 a | 3.95 ± 0.07 a | 4.16 ± 0.02 b | 4.29 ± 0.02 b | 5.41 ± 0.02 c | 4.34 ± 0.02 b |
| ACP 3 | 166 ± 0.81 a | 191.94 ± 1.07 b | 196.11 ± 3.12 b | 224.22 ± 2.99 c | 268.66 ± 1.20 e | 236.25 ± 2.37 d |
| Serum Peroxidase 4 | 0.63 ± 0.014 a | 0.82 ± 0.022 b | 1.32 ± 0.036 c | 1.24 ± 0.029 c | 1.46 ± 0.030 d | 1.33 ± 0.026 c |
| Mucus Peroxidase 5 | 0.20 ± 0.005 a | 0.29 ± 0.008 b | 0.35 ± 0.010 c | 0.34 ± 0.008 c | 0.43 ± 0.012 d | 0.44 ± 0.013 d |
| Antiprotease 6 | 77.12 ± 0.21 a | 79.17 ± 0. 56 ab | 82.31 ± 0.74 cd | 80.29 ± 0.44 bc | 87.37 ± 0.52 e | 83.28 ± 0.42 d |
| Phagocytosis 7 | 28.87 ± 0.36 a | 32.07 ± 0.17 b | 42.43 ± 0.17 d | 40.13 ± 0.19 c | 45.87 ± 0.20 e | 41.87 ± 0.16 d |
| Super oxide anion 8 | 40.95 ± 0.74 a | 41.78 ± 0.75 a | 46.78 ± 0.48 b | 46.21 ± 0.28 b | 49.52 ± 0.61 c | 48.63 ± 0.22 bc |
| RB 9 | 0.29 ± 0.007 a | 0.30 ± 0.007 a | 0.41 ± 0.010 b | 0.39 ± 0.010 b | 0.49 ± 0.012 c | 0.48 ± 0.013 c |
| After 90 Days | ||||||
|---|---|---|---|---|---|---|
| Parameters | C | E1 | E2 | E3 | E4 | E5 |
| SOD 1 | 10.69 ± 0.13 a | 12.75 ± 0.19 b | 14.38 ± 0.26 cd | 13.76 ± 0.28 bc | 17.75 ± 0.35 e | 15.03 ± 0.24 d |
| Catalase 2 | 2.97 ± 0.02 a | 3.24 ± 0.01 b | 3.32 ± 0.02 c | 3.8 ± 0.02 d | 3.95 ± 0.02 e | 3.82 ± 0.02 d |
| Lipid peroxidation 3 | 19.42 ± 0.23 d | 17.68 ± 0.25 c | 14.56 ± 0.26 b | 15.23 ± 0.31 b | 12.76 ± 0.25 a | 13.79 ± 0.21 ab |
| After challenge with Aeromonas hydrophila | ||||||
| SOD 1 | 14.39 ± 0.20 a | 17.48 ± 0.20 b | 19.68 ± 0.38 c | 18.75 ± 0.31 bc | 22.97 ± 0.43 d | 19.57 ± 0.28 c |
| Catalase 2 | 3.23 ± 0.02 a | 3.86 ± 0.02 c | 3.47 ± 0.02 b | 3.95 ± 0.02 c | 4.28 ± 0.03 e | 4.06 ± 0.02 d |
| Lipid peroxidation 3 | 23.89 ± 0.33 d | 22.04 ± 0.25 c | 18.32 ± 0.35 b | 19.3 ± 0.32 b | 15.76 ± 0.29 a | 16.6 ± 0.23 a |
| Negative Control | Positive Control | E1 | E2 | E3 | E4 | E5 |
|---|---|---|---|---|---|---|
| 96.66 ± 3.33 d | 36.66 ± 8.81 a | 56.67 ± 3.33 ab | 70 ± 5.77 bc | 66.67 ± 3.33 bc | 93.33 ± 3.33 d | 86.66 ± 3.33 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukul, T.; Kari, Z.A.; Téllez-Isaías, G.; Ghosh, K. Autochthonous Bacilli and Fructooligosaccharide as Functional Feed Additives Improve Growth, Feed Utilisation, Haemato-Immunological Parameters and Disease Resistance in Rohu, Labeo rohita (Hamilton). Animals 2023, 13, 2631. https://doi.org/10.3390/ani13162631
Sukul T, Kari ZA, Téllez-Isaías G, Ghosh K. Autochthonous Bacilli and Fructooligosaccharide as Functional Feed Additives Improve Growth, Feed Utilisation, Haemato-Immunological Parameters and Disease Resistance in Rohu, Labeo rohita (Hamilton). Animals. 2023; 13(16):2631. https://doi.org/10.3390/ani13162631
Chicago/Turabian StyleSukul, Tanaya, Zulhisyam Abdul Kari, Guillermo Téllez-Isaías, and Koushik Ghosh. 2023. "Autochthonous Bacilli and Fructooligosaccharide as Functional Feed Additives Improve Growth, Feed Utilisation, Haemato-Immunological Parameters and Disease Resistance in Rohu, Labeo rohita (Hamilton)" Animals 13, no. 16: 2631. https://doi.org/10.3390/ani13162631