
Preliminary Investigation of Mixed Orchard Hays on the Meat Quality, Fatty Acid Profile, and Gastrointestinal Microbiota in Goat Kids

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Design
2.2. Experimental Animal Feeding and Management
2.3. Sample Collection
2.4. Meat Quality Determination
2.4.1. Meat pH Determination
2.4.2. Meat Color Determination
2.4.3. Drip Loss and Cooking Loss Determination
2.4.4. Shear Force Determination
2.5. Fatty Acid Profile Analysis
2.6. The Profiles of Total Amino Acids and Free Amino Acids
2.7. 16S rDNA Determination of Rumen and Intestinal Microorganisms
2.8. Statistical Analysis
3. Results
3.1. Final Body Weight
3.2. Meat Quality Characteristics
3.3. Relative Percentage of Fatty Acids in Longissimus Dorsi Muscle
3.4. Essential Amino Acid and Nonessential Amino Acid Contents in longissimus dorsi Muscles of Saanen Dairy Goats
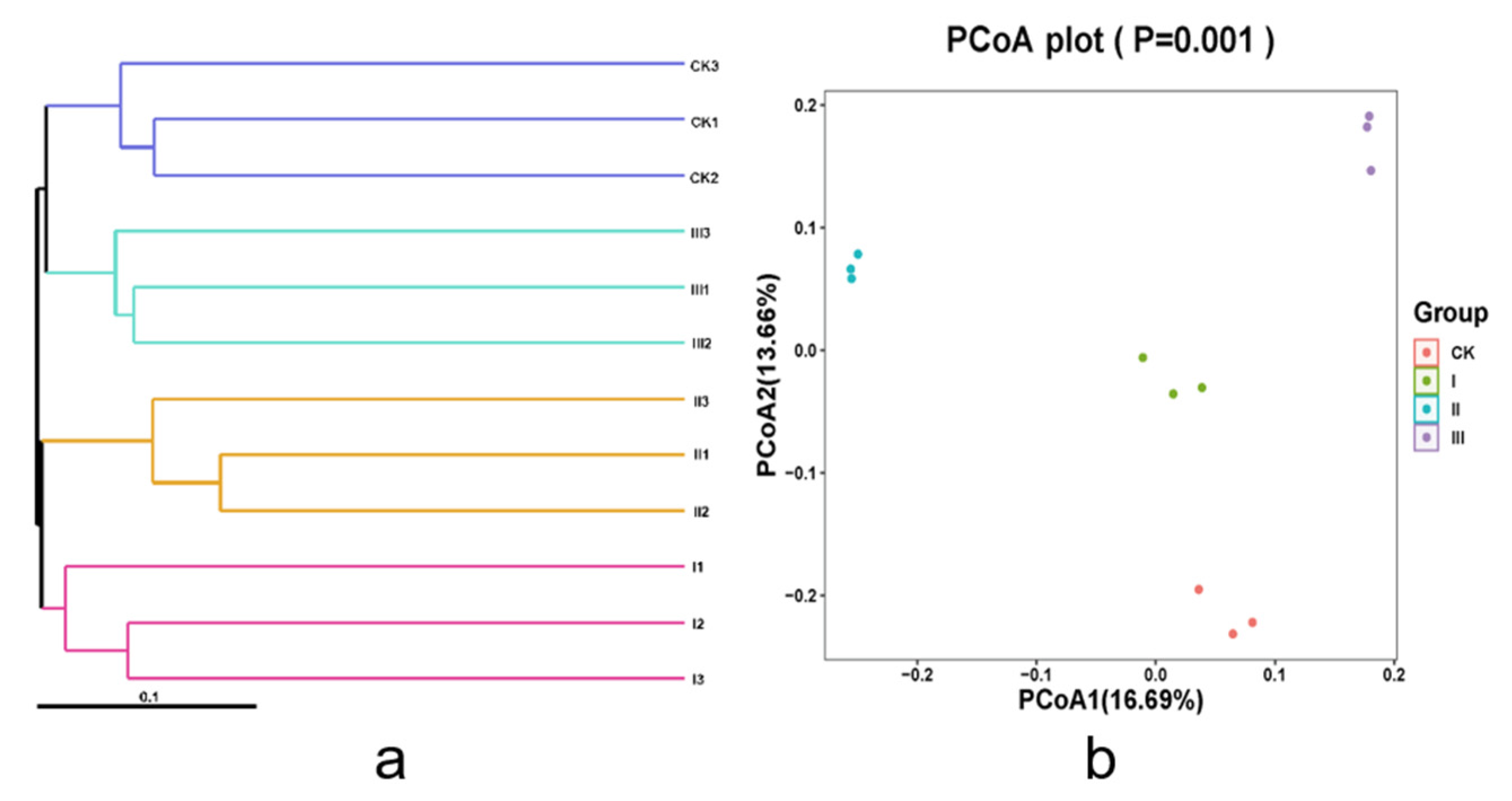
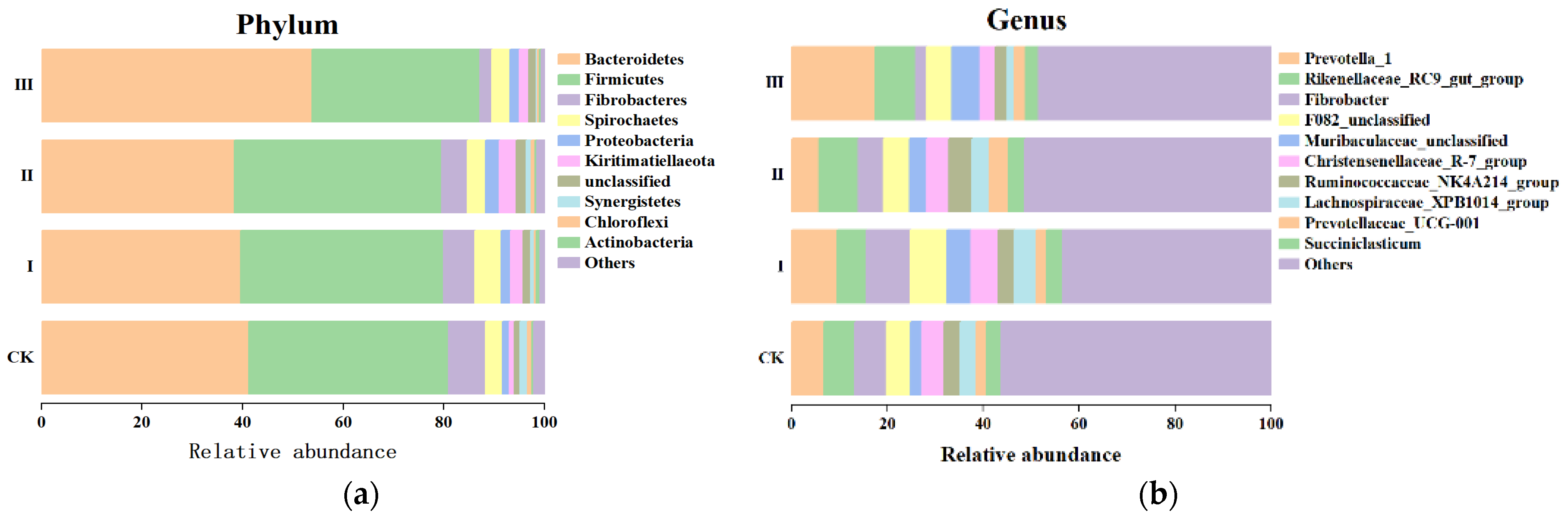
3.5. Rumen Bacteria Diversity of Goats Fed with Different Mixed Hays
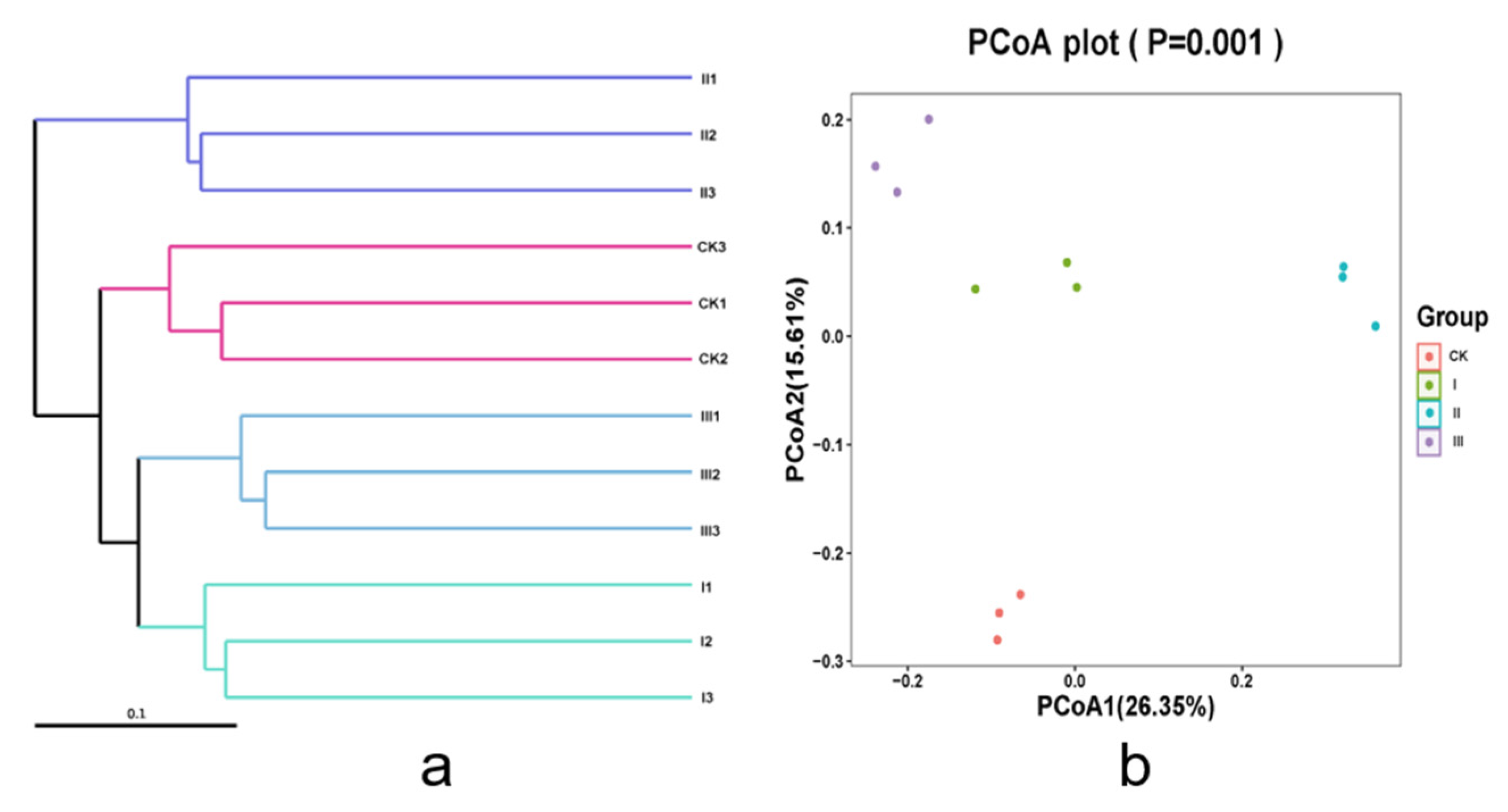
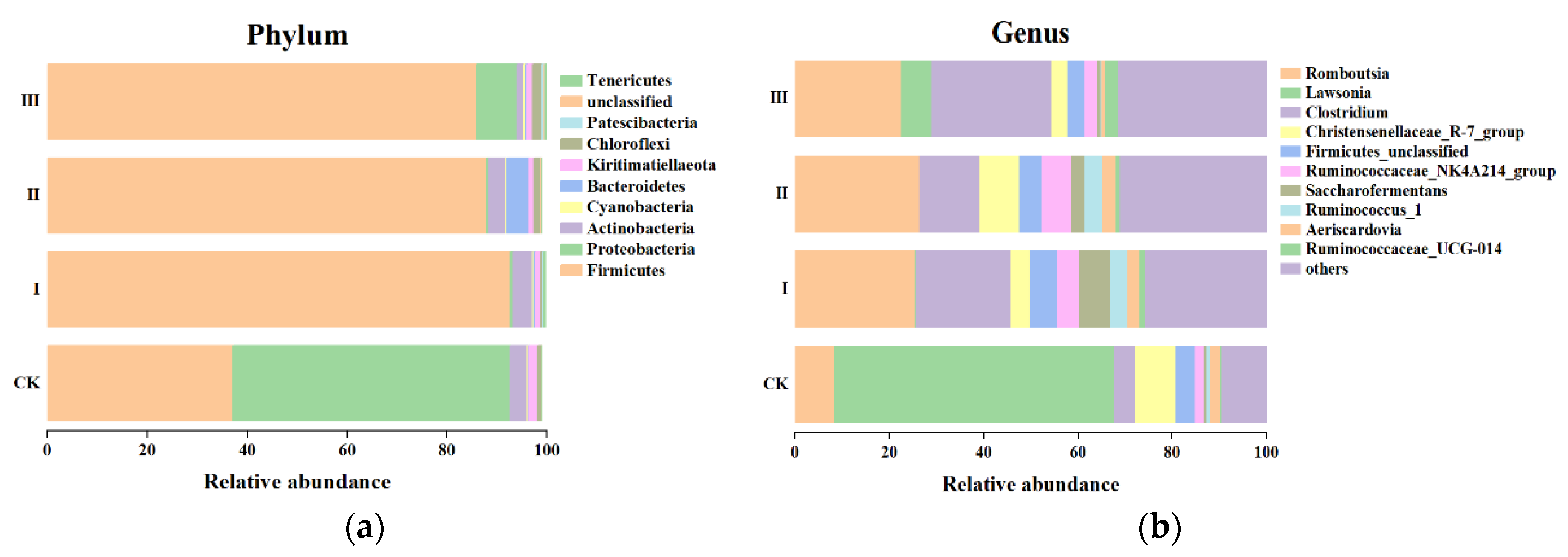
3.6. Intestinal Bacteria Diversity of Goats Fed with Different Mixed Hays
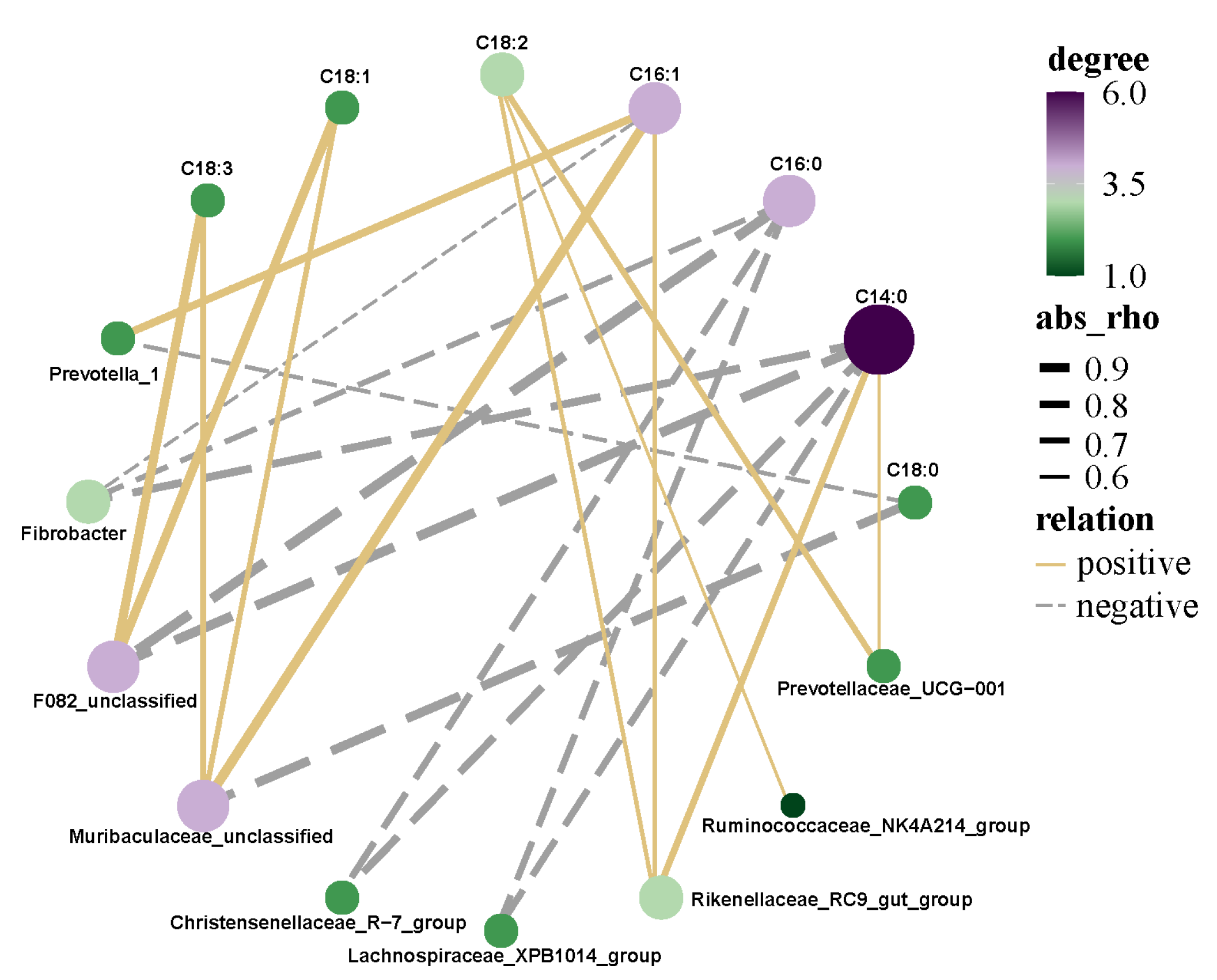
3.7. Correlation Analysis of Rumen Microflora and Fatty Acids of Saanen Dairy Goats
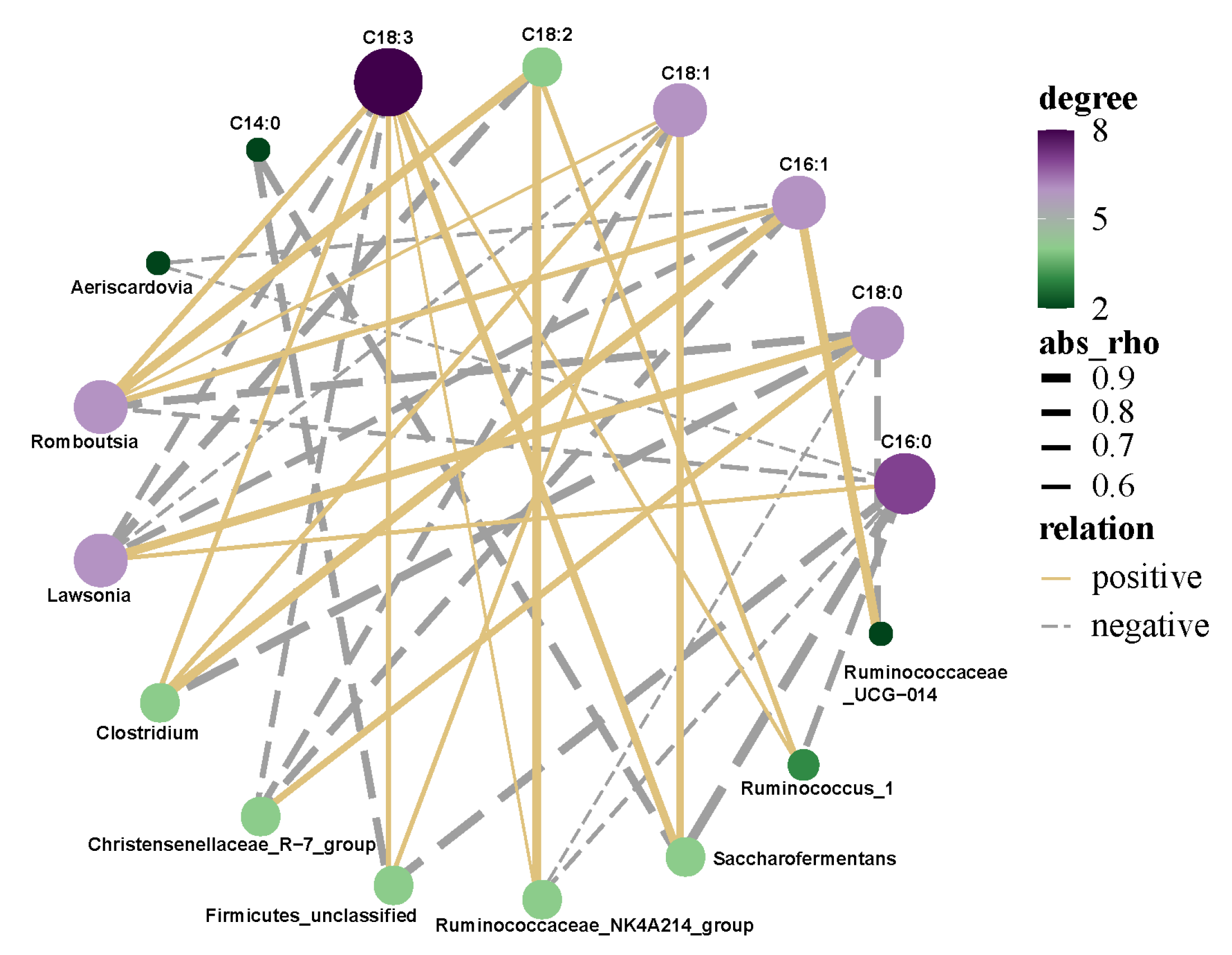
3.8. Correlation Analysis of Intestinal Microflora and Fatty Acids of Saanen Dairy Goats
4. Discussion
4.1. Meat Quality Characteristics
4.2. Relative Percentage of Fatty Acids in Longissimus Dorsi Muscles
4.3. Essential Amino Acid and Nonessential Amino Acid Contents of longissimus dorsi Muscles in Saanen Dairy Goats
4.4. The Rumen Microflora Composition of Goats Fed with Mixed Hay
4.5. The Intestinal Microflora Composition of Goats Fed with Mixed Hay
4.6. Correlation Analysis of Rumen Microflora and Fatty Acids of Saanen Dairy Goats
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Mottet, A.; Teillard, F.; Boettcher, P.; De’ Besi, G.; Besbes, B. Review: Domestic herbivores and food security: Current contribution, trends and challenges for a sustainable development. Animal 2018, 12, S188–S198. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhao, Y.; Yu, C.; Luo, L.; Pan, Y. Land management influences trade-offs and the total supply of ecosystem services in alpine grassland in Tibet, China. J. Environ. Manag. 2017, 193, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, F.W.; Blignaut, D.J.C.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Kinobe, R.T.; Malau-Aduli, A.E.O. Lipid Metabolism, Carcass Characteristics and Longissimus dorsi Muscle Fatty Acid Composition of Tropical Crossbred Beef Cattle in Response to Desmanthus spp. Forage Backgrounding. Metabolites 2021, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Goncalves da Silva, P.C.; Brandao Ferreira Itavo, C.C.; Vinhas Itavo, L.C.; Bonin Gomes, M.d.N.; Dias Feijo, G.L.; Silva Monteiro Ferelli, K.L.; Heimbach, N.d.S.; da Silva, J.A.; Ayardes de Melo, G.K.; Filgueira Pereira, M.W. Carcass traits and meat quality of Texel lambs raised in Brachiaria pasture and feedlot systems. Anim. Sci. J. 2020, 91, e13394. [Google Scholar] [CrossRef]
- Kagan, I.A. Soluble phenolic compounds of perennial ryegrass (Lolium perenne L.): Potential effects on animal performance, and challenges in determining profiles and concentrations. Anim. Feed Sci. Technol. 2021, 277, 114960. [Google Scholar] [CrossRef]
- Khiaosa-ard, R.; Kreuzer, M.; Leiber, F. Apparent recovery of C18 polyunsaturated fatty acids from feed in cow milk: A meta-analysis of the importance of dietary fatty acids and feeding regimens in diets without fat supplementation. J. Dairy Sci. 2015, 98, 6399–6414. [Google Scholar] [CrossRef]
- Rodriguez, R.; Alomar, D.; Morales, R. Milk and meat fatty acids from sheep fed a plantain-chicory mixture or a grass-based permanent sward. Animal 2020, 14, 1102–1109. [Google Scholar] [CrossRef]
- Siurana, A.; Calsamiglia, S. A metaanalysis of feeding strategies to increase the content of conjugated linoleic acid (CLA) in dairy cattle milk and the impact on daily human consumption. Anim. Feed Sci. Technol. 2016, 217, 13–26. [Google Scholar] [CrossRef]
- Magan, J.B.; O’Callaghan, T.F.; Kelly, A.L.; McCarthy, N.A. Compositional and functional properties of milk and dairy products derived from cows fed pasture or concentrate-based diets. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2769–2800. [Google Scholar] [CrossRef]
- Logan, B.G.; Bush, R.D.; Biffin, T.E.; Hopkins, D.L.; Smith, M.A. Measurement of drip loss in alpaca (Vicugna pacos) meat using different techniques and sample weights. Meat Sci. 2019, 151, 1–3. [Google Scholar] [CrossRef]
- Atsbha, K.; Gebremariam, T.; Aregawi, T. Slaughter performance and meat quality of Begait breed lambs fattened under different diets. Heliyon 2021, 7, e06935. [Google Scholar] [CrossRef] [PubMed]
- De Brito, G.F.; Holman, B.W.B.; McGrath, S.R.; Friend, M.A.; van de Ven, R.; Hopkins, D.L. The effect of forage-types on the fatty acid profile, lipid and protein oxidation, and retail colour stability of muscles from White Dorper lambs. Meat Sci. 2017, 130, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Z.; Cui, Y.; Lu, H.; Wang, G.; Ma, X. Effect of long-term dietary probiotic Lactobacillus reuteri 1 or antibiotics on meat quality, muscular amino acids and fatty acids in pigs. Meat Sci. 2021, 171, 108234. [Google Scholar] [CrossRef] [PubMed]
- Orzuna-Orzuna, J.F.; Dorantes-Iturbide, G.; Lara-Bueno, A.; Mendoza-Martinez, G.D.; Miranda-Romero, L.A.; Lee-Rangel, H.A. Growth Performance, Meat Quality and Antioxidant Status of Sheep Supplemented with Tannins: A Meta-Analysis. Animals 2021, 11, 3184. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, Z.; Chen, Y.; Liu, X.; Liu, K.; Zhang, Y.; Luo, H. Carcass Traits, Meat Quality, and Volatile Compounds of Lamb Meat from Different Restricted Grazing Time and Indoor Supplementary Feeding Systems. Foods 2021, 10, 2822. [Google Scholar] [CrossRef]
- Sgarro, M.F.; Maggiolino, A.; Pateiro, M.; Dominguez, R.; Iannaccone, F.; De Palo, P.; Lorenzo, J.M. Effects of Anthocyanin Supplementation and Ageing Time on the Volatile Organic Compounds and Sensory Attributes of Meat from Goat Kids. Animals 2022, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Sanudo, C.; Alfonso, M.; Sanchez, A.; Delfa, R.; Teixeira, A. Carcass and meat quality in light lambs from different fat classes in the EU carcass classification system. Meat Sci. 2000, 56, 89–94. [Google Scholar] [CrossRef]
- De Almeida Rego, F.C.; Francozo, M.C.; Ludovico, A.; Boscaro de Castro, F.A.; Zundt, M.; Lupo, C.R.; Belan, L.; Coelho Cunha Filho, L.F.; dos Santos, J.S.; Castilho, C. Fatty acid profile and lambs’ meat quality fed with different levels of crude glycerin replacing corn. Semin. Cienc. Agrar. 2017, 38, 2051–2064. [Google Scholar] [CrossRef] [Green Version]
- Jaakamo, M.J.; Luukkonen, T.J.; Kairenius, P.K.; Bayat, A.R.; Ahvenjarvi, S.A.; Tupasela, T.M.; Vilkki, J.H.; Shingfield, K.J.; Leskinen, H.M. The effect of dietary forage to concentrate ratio and forage type on milk fatty acid composition and milk fat globule size of lactating cows. J. Dairy Sci. 2019, 102, 8825–8838. [Google Scholar] [CrossRef]
- Hou, J.; Wang, F.; Wang, Y.; Liu, F. Effects of vitamin E on the concentration of conjugated linoleic acids and accumulation of intermediates of ruminal biohydrogenation in vitro. Small Rumin. Res. 2013, 111, 63–70. [Google Scholar] [CrossRef]
- Pertiwi, K.; Kupers, L.K.; de Goede, J.; Zock, P.L.; Kromhout, D.; Geleijnse, J.M. Dietary and Circulating Long-Chain Omega-3 Polyunsaturated Fatty Acids and Mortality Risk After Myocardial Infarction: A Long-Term Follow-Up of the Alpha Omega Cohort. J. Am. Heart Assoc. 2021, 10, e022617. [Google Scholar] [CrossRef] [PubMed]
- Kitessa, S.; Liu, S.; Briegel, J.; Pethick, D.; Gardner, G.; Ferguson, M.; Allingham, P.; Nattrass, G.; McDonagh, M.; Ponnampalam, E.; et al. Effects of intensive or pasture finishing in spring and linseed supplementation in autumn on the omega-3 content of lamb meat and its carcass distribution. Anim. Prod. Sci. 2010, 50, 130–137. [Google Scholar] [CrossRef]
- Pannier, L.; Ponnampalam, E.N.; Gardner, G.E.; Hopkins, D.L.; Ball, A.J.; Jacob, R.H.; Pearce, K.L.; Pethick, D.W. Prime Australian lamb supplies key nutrients for human health. Anim. Prod. Sci. 2010, 50, 1115–1122. [Google Scholar] [CrossRef] [Green Version]
- Piao, M.Y.; Jo, C.; Kim, H.J.; Lee, H.J.; Kim, H.J.; Ko, J.-Y.; Baik, M. Comparison of Carcass and Sensory Traits and Free Amino Acid Contents among Quality Grades in Loin and Rump of Korean Cattle Steer. Asian-Australas. J. Anim. Sci. 2015, 28, 1629–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoard, S.A.; Sales, F.A.; Sciascia, Q.L. Amino acids in sheep production. Front. Biosci. 2016, 8, 264–288. [Google Scholar] [CrossRef]
- Lu, C.; Zeng, S.H.; Zhang, T.; Qi, Z.Q.; Hu, Y.G. Research progress in the study of bio-active composition. Pratacult. Sci. 2005, 22, 28–32. (In Chinese) [Google Scholar]
- Kung, L.M.; Rode, L.M. Amino acid metabolism in ruminants. Anim. Feed Sci. Technol. 1996, 59, 167–172. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, X.F.; Zhan, J.S.; Huo, J.H.; Huang, Q.L.; Li, W.Y. Efeects of Rumen Lysine supplementation on growth performance, Nutrient Apparent Digestibility and Nitrogen Metabolism of Fuqing Goats. Chin. J. Anim. Nutr. 2022, 34, 432–441. (In Chinese) [Google Scholar]
- Raguindin, P.F.; Itodo, O.A.; Stoyanov, J.; Dejanovic, G.M.; Gamba, M.; Asllanaj, E.; Minder, B.; Bussler, W.; Metzger, B.; Muka, T.; et al. A systematic review of phytochemicals in oat and buckwheat. Food Chem. 2021, 338, 127982. [Google Scholar] [CrossRef]
- Feng, Z.H.; Gao, Y.X.; Li, J.G. Research progress of saponins regulating rumen fermentation and production performance in ruminants. Chin. J. Anim. Nutr. 2010, 22, 1515–1522. (In Chinese) [Google Scholar]
- Cremonesi, P.; Conte, G.; Severgnini, M.; Turri, F.; Monni, A.; Capra, E.; Rapetti, L.; Colombini, S.; Chessa, S.; Battelli, G.; et al. Evaluation of the effects of different diets on microbiome diversity and fatty acid composition of rumen liquor in dairy goat. Animal 2018, 12, 1856–1866. [Google Scholar] [CrossRef] [PubMed]
- Ellison, M.J.; Conant, G.C.; Cockrum, R.R.; Austin, K.J.; Huan, T.; Becchi, M.; Lamberson, W.R.; Cammack, K.M. Diet Alters Both the Structure and Taxonomy of the Ovine Gut Microbial Ecosystem. DNA Res. 2014, 21, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, X.; Zhang, L.; Luo, J.; Zhao, S.; Jiao, T. Effects of Oat Hay Content in Diets on Nutrient Metabolism and the Rumen Microflora in Sheep. Animals 2020, 10, 2341. [Google Scholar] [CrossRef] [PubMed]
- Dehority, B.A.; Tirabasso, P.A. Effect of ruminal cellulolytic bacterial concentrations on in situ digestion of forage cellulose. J. Anim. Sci. 1998, 76, 2905–2911. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.D.; Mamuad, L.L.; Kim, S.H.; Kang, S.H.; Kim, E.T.; Cho, Y.I.; Lee, S.S.; Lee, S.S. Diet Transition from High-Forage to High-Concentrate Alters Rumen Bacterial Community Composition, Epithelial Transcriptomes and Ruminal Fermentation Parameters in Dairy Cows. Animals 2021, 11, 838. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, M.; Xue, C.; Zhu, W.; Mao, S. Characterization and comparison of the temporal dynamics of ruminal bacterial microbiota colonizing rice straw and alfalfa hay within ruminants. J. Dairy Sci. 2016, 99, 9668–9681. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Teather, R.; Forster, R. Composition, spatial distribution, and diversity of the bacterial communities in the rumen of cows fed different forages. FEMS Microbiol. Ecol. 2010, 74, 612–622. [Google Scholar] [CrossRef]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- Myer, P.R.; Wells, J.E.; Smith, T.P.L.; Kuehn, L.A.; Freetly, H.C. Cecum microbial communities from steers differing in feed efficiency. J. Anim. Sci. 2015, 93, 5327–5340. [Google Scholar] [CrossRef] [Green Version]
- Reti, K.L.; Thomas, M.C.; Yanke, L.J.; Selinger, L.B.; Inglis, G.D. Effect of antimicrobial growth promoter administration on the intestinal microbiota of beef cattle. Gut Pathog. 2013, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Valerio de Oliveira, M.N.; Jewell, K.A.; Freitas, F.S.; Benjamin, L.A.; Totola, M.R.; Borges, A.C.; Moraes, C.A.; Suen, G. Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore steer. Vet. Microbiol. 2013, 164, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wang, X.; Bernstein, S.; Huffman, M.A.; Xia, D.-P.; Gu, Z.; Chen, R.; Sheeran, L.K.; Wagner, R.S.; Li, J. Marked variation between winter and spring gut microbiota in free-ranging Tibetan Macaques (Macaca thibetana). Sci. Rep. 2016, 6, 26035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Yang, Y.; Zhang, Y.; Lv, S.; Jin, T.; Li, K.; Han, Z.; Li, Y. Microbiome analysis reveals the alterations in gut microbiota in different intestinal segments of Yimeng black goats. Microb. Pathog. 2021, 155, 104900. [Google Scholar] [CrossRef] [PubMed]
- Rastmanesh, R. High polyphenol, low probiotic diet for weight loss because of intestinal microbiota interaction. Chem.-Biol. Interact. 2011, 189, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Wells, W.G.; Smith, C.J. Characterization of the primary starch utilization operon in the obligate anaerobe Bacteroides fragilis: Regulation by carbon source and oxygen. J. Bacteriol. 2006, 188, 4663–4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xu, T.; Zhu, W.; Mao, S. High-Grain feeding alters caecal bacterial microbiota composition and fermentation and results in caecal mucosal injury in goats. Br. J. Nutr. 2014, 112, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buccioni, A.; Decandia, M.; Minieri, S.; Molle, G.; Cabiddu, A. Lipid metabolism in the rumen: New insights on lipolysis and biohydrogenation with an emphasis on the role of endogenous plant factors. Anim. Feed Sci. Technol. 2012, 174, 1–25. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef]
- Toral, P.G.; Monahan, F.J.; Hervas, G.; Frutos, P.; Moloney, A.P. Review: Modulating ruminal lipid metabolism to improve the fatty acid composition of meat and milk. Challenges and opportunities. Animal 2018, 12, S272–S281. [Google Scholar] [CrossRef] [Green Version]
- Gillis, M.H.; Duckett, S.K.; Sackmann, J.R. Effects of supplemental rumen-protected conjugated linoleic acid or corn oil on fatty acid composition of adipose tissues in beef cattle. J. Anim. Sci. 2004, 82, 1419–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Zeng, S.; Zhang, R.; Diao, Q.; Tu, Y. Effects of dietary energy levels on rumen bacterial community composition in Holstein heifers under the same forage to concentrate ratio condition. BMC Microbiol. 2018, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, T.; Wang, X.; Geng, Y.; Zhao, N.; Hu, L.; Liu, H.; Kang, S.; Xu, S. Effect of Dietary Protein Levels on Dynamic Changes and Interactions of Ruminal Microbiota and Metabolites in Yaks on the Qinghai-Tibetan Plateau. Front. Microbiol. 2021, 12, 684340. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Group | |||
|---|---|---|---|---|
| CK 1 | I 2 | II 3 | III 4 | |
| Dry matter (%) | 81.28 | 84.90 | 85.98 | 83.72 |
| Crude protein (%) | 13.32 | 15.5 | 16.80 | 16.00 |
| Neutral detergent fibre (%) | 41.57 | 40.90 | 41.20 | 35.70 |
| Acid detergent fibre (%) | 26.50 | 28.5 | 27.70 | 29.30 |
| Crude ash (%) | 11.90 | 10.80 | 11.53 | 12.20 |
| Group | Final Body Weight/kg |
|---|---|
| CK 1 | 16.99 ± 0.72 c |
| I 2 | 21.92 ± 0.84 a |
| II 3 | 19.9 ± 0.74 b |
| III 4 | 19.73 ± 0.21 b |
| Items | Group | p-Value | SEM 5 | |||
|---|---|---|---|---|---|---|
| CK 1 | I 2 | II 3 | III 4 | |||
| Shear force/N | 37.45 ± 0.32 a | 35.76 ± 1.15 a,b | 33.22 ± 1.50 b | 37.87 ± 0.17 a | 0.03 | 1.36 |
| Cooking loss/% | 62.53 ± 1.33 c | 65.71 ± 0.66 a,b | 66.17 ± 0.39 a | 63.25 ± 0.40 b,c | 0.03 | 1.12 |
| Drip loss/% | 3.43 ± 0.01 | 3.30 ± 0.003 | 2.99 ± 0.35 | 3.26 ± 0.01 | 0.41 | 0.25 |
| L* 45 min | 36.81 ± 0.45 a,b | 35.35 ± 0.63 b | 37.97 ± 0.85 a | 36.35 ± 0.35 a,b | 0.08 | 0.85 |
| a* 45 min | 9.46 ± 0.68 a,b | 10.48 ± 0.46 a | 8.56 ± 0.28 b | 10.51 ± 0.09 a | 0.04 | 0.62 |
| b* 45 min | 11.29 ± 0.56 | 11.03 ± 0.57 | 11.17 ± 1.19 | 11.77 ± 0.46 | 0.91 | 1.07 |
| pH 45 min | 6.69 ± 0.01 a,b | 6.66 ± 0.01 b | 6.69 ± 0.01 a | 6.66 ± 0.01 b | 0.07 | 0.01 |
| pH 24 h | 5.62 ± 0.01 | 5.61 ± 0.003 | 5.62 ± 0.01 | 5.62 ± 0.01 | 0.80 | 0.01 |
| Fatty Acid (%) | Group | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| CK 1 | I 2 | II 3 | III 4 | |||
| C16:0 | 19.66 ± 0.06 a | 15.63 ± 0.15 d | 18.13 ± 0.09 c | 19.16 ± 0.09 b | <0.001 | 0.14 |
| C18:0 | 28.90 ± 0.23 a | 23.73 ± 0.12 c | 24.83 ± 0.39 b | 23.17 ± 0.19 c | <0.001 | 0.36 |
| C14:0 | 1.26 ± 0.02 b | 0.81 ± 0.00 c | 1.38 ± 0.04 a | 1.38 ± 0.01 a | <0.001 | 0.32 |
| C16:1 | 0.56 ± 0.05 d | 1.18 ± 0.01 b | 1.05 ± 0.02 c | 1.58 ± 0.01 a | <0.001 | 0.39 |
| C18:1 | 44.70 ± 0.26 c | 50.14 ± 0.03 a | 45.20 ± 0.45 c | 46.97 ± 0.23 b | <0.001 | 0.41 |
| C18:2 | 3.59 ± 0.30 c | 6.14 ± 0.05 b | 7.72 ± 0.08 a | 5.92 ± 0.03 b | <0.001 | 0.22 |
| C18:3 | 1.24 ± 0.02 d | 2.36 ± 0.02 a | 1.65 ± 0.01 c | 1.75 ± 0.01 b | <0.001 | 0.02 |
| n-6: n-3 | 2.89 ± 0.21 c | 2.60 ± 0.03 c | 4.68 ± 0.05 a | 3.39 ± 0.03 b | <0.001 | 0.15 |
| Amino Acid (g/100 g) | Group | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| CK | I | II | III | |||
| Total amino acids | 19.55 ± 0.46 d | 24.43±0.41a | 22.13±0.11b | 20.72±0.22 c | <0.001 | 0.47 |
| Essential amino acids | ||||||
| Threonine | 0.94 ± 0.02 c | 1.18 ± 0.01 a | 1.06 ± 0.01 b | 0.98 ± 0.01 c | <0.001 | 0.02 |
| Valine | 0.91 ± 0.02 c | 1.17 ± 0.01 a | 1.03 ± 0.01 b | 0.95 ± 0.02 c | <0.001 | 0.02 |
| Methionine | 0.51 ± 0.01 b | 0.63 ± 0.01 a | 0.55 ± 0.02 b | 0.52 ± 0.02 b | 0.002 | 0.02 |
| Isoleucine | 0.89 ± 0.02 c | 1.12 ± 0.02 a | 0.99 ± 0.01 b | 0.92 ± 0.01 c | <0.001 | 0.02 |
| Leucine | 1.58 ± 0.03 d | 1.98 ± 0.03 a | 1.77 ± 0.01 b | 1.69 ± 0.01 c | <0.001 | 0.03 |
| Tyrosine | 0.67 ± 0.02 c | 0.85 ± 0.01 a | 0.76 ± 0.01 b | 0.71 ± 0.01 c | <0.001 | 0.02 |
| Phenylalanine | 0.85 ± 0.02 c | 1.10 ± 0.01 a | 0.96 ± 0.02 b | 0.93 ± 0.02 b | <0.001 | 0.02 |
| Lysine | 1.86 ± 0.04 c | 2.32 ± 0.03 a | 2.06 ± 0.04 b | 1.89 ± 0.06 c | <0.001 | 0.06 |
| Histidine | 0.70 ± 0.02 b | 0.84 ± 0.01 a | 0.83 ± 0.00 a | 0.81 ± 0.03 a | 0.002 | 0.03 |
| Nonessential amino acids | ||||||
| Asparagine | 1.87 ± 0.04 c | 2.35 ± 0.02 a | 2.09 ± 0.02 b | 1.96 ± 0.03 c | <0.001 | 0.04 |
| Serine | 0.76 ± 0.01 d | 0.95 ± 0.01 a | 0.87 ± 0.01 b | 0.81 ± 0.01 c | <0.001 | 0.01 |
| Glutamic acid | 2.93 ± 0.07 d | 3.64 ± 0.06 a | 3.29 ± 0.03 b | 3.10 ± 0.02 c | <0.001 | 0.07 |
| Proline | 1.59 ± 0.11 c | 2.12 ± 0.09 a | 2.02 ± 0.02 a,b | 1.86 ± 0.04 b | 0.004 | 0.1 |
| Glycine | 0.93 ± 0.01 b | 1.05 ± 0.05 a | 1.00 ± 0.02a,b | 0.94 ± 0.01 b | 0.078 | 0.04 |
| Alanine | 1.22 ± 0.02 c | 1.48 ± 0.05 a | 1.34 ± 0.02 b | 1.28 ± 0.02 b,c | 0.001 | 0.04 |
| Cystine | 0.13 ± 0.00 b | 0.16 ± 0.01 a | 0.14 ± 0.01 a,b | 0.13 ± 0.01 a,b | 0.082 | 0.01 |
| Arginine | 1.22 ± 0.03 c | 1.47 ± 0.04 a | 1.38 ± 0.01 b | 1.24 ± 0.02 c | 0.001 | 0.04 |
| Items | Group | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| CK | I | II | III | |||
| OTUs | 1571 ± 55 | 1750 ± 134 | 1328 ± 48 | 1596 ± 148 | 0.12 | 151 |
| Shannon | 9.02 ± 0.09 | 9.10 ± 0.14 | 9.11 ± 0.08 | 9.09 ± 0.11 | 0.94 | 0.15 |
| Simpson | 0.99 ± 0.003 | 0.99 ± 0.003 | 1.00 | 0.99 ± 0.003 | 0.33 | 0.004 |
| Chao1 | 1592.05 ± 59.22 | 1782.02 ± 148.10 | 1331.70 ± 47.11 | 1618.25 ± 159.94 | 0.12 | 163.16 |
| Goods_coverage | 0.997 | 0.996 | 0.999 | 0.997 | 0.19 | 0.001 |
| Items | Group | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| CK | I | II | III | |||
| OTU | 581 ± 22 b,c | 728 ± 116 b | 1025 ± 75 a | 449 ± 53 c | <0.01 | 106 |
| Shannon | 3.79 ± 0.27 d | 6.31 ± 0.19 b | 7.04 ± 0.14 a | 5.28 ± 0.23 c | <0.01 | 0.30 |
| Simpson | 0.65 ± 0.08 b | 0.95 ± 0.01 a | 0.95 ± 0.01 a | 0.92 ± 0.01 a | <0.01 | 0.03 |
| Chao1 | 597.57 ± 24.92 b,c | 735.61 ± 119.72 b | 1040.46 ± 88.90 a | 451.93 ± 54.46 c | <0.01 | 113.63 |
| Goods_coverage | 0.998 | 0.999 | 0.997 | 0.999 | 0.11 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, T.; Chen, X.; Liu, C.; Jin, X.; Tan, H.; Long, M. Preliminary Investigation of Mixed Orchard Hays on the Meat Quality, Fatty Acid Profile, and Gastrointestinal Microbiota in Goat Kids. Animals 2022, 12, 780. https://doi.org/10.3390/ani12060780
Wang Y, Li T, Chen X, Liu C, Jin X, Tan H, Long M. Preliminary Investigation of Mixed Orchard Hays on the Meat Quality, Fatty Acid Profile, and Gastrointestinal Microbiota in Goat Kids. Animals. 2022; 12(6):780. https://doi.org/10.3390/ani12060780
Chicago/Turabian StyleWang, Yingying, Tengfei Li, Xinyi Chen, Chongyi Liu, Xumei Jin, Hua Tan, and Mingxiu Long. 2022. "Preliminary Investigation of Mixed Orchard Hays on the Meat Quality, Fatty Acid Profile, and Gastrointestinal Microbiota in Goat Kids" Animals 12, no. 6: 780. https://doi.org/10.3390/ani12060780