
Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farms, Animals and Diet
2.2. Sampling and Analysis
2.3. Statistical Analysis
3. Results and Discussion
3.1. Pasture Composition
3.2. Milk and Cheese Composition
3.3. Multivariate Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciotola, F.; Albarella, S.; Liotta, L.; Contessa, A.; Di Meo, G.P.; Barbieri, V.; Peretti, V. Native cattle breeds of Italy: Karyological profile. Ital. J. Anim. Sci. 2009, 8, 54–56. [Google Scholar] [CrossRef]
- Bonanno, A.; Tornambè, G.; Bellina, V.; De Pasquale, C.; Mazza, F.; Maniaci, G.; Di Grigoli, A. Effect of farming system and cheese making technology on the physicochemical characteristics, fatty acid profile, and sensory properties of Caciocavallo Palermitano cheese. J. Dairy Sci. 2013, 96, 710–724. [Google Scholar] [CrossRef]
- Di Gregorio, P.; Di Grigoli, A.; Di Trana, A.; Alabiso, M.; Maniaci, G.; Rando, A.; Vallucci, C.; Finizio, D.; Bonanno, A. Effects of different genotypes at the CSN3 and LGB loci on milk and cheese-making characteristics of the bovine Cinisara breed. Int. Dairy J. 2017, 71, 1–5. [Google Scholar] [CrossRef]
- Maniaci, G.; Alabiso, M.; Francesca, N.; Giosuè, C.; Di Grigoli, A.; Corona, O.; Cardamone, C.; Graci, G.; Portolano, B.; Bonanno, A. Bresaola made from Cinisara cattle: Effect of muscle type and animal category on physicochemical and sensory traits. CyTA J. Food 2020, 18, 383–391. [Google Scholar] [CrossRef]
- Alabiso, M.; Maniaci, G.; Giosuè, C.; Di Grigoli, A.; Bonanno, A. Fatty Acid Composition of Salami Made by Meat from Different Commercial Categories of Indigenous Dairy Cattle. Animals 2021, 11, 1060. [Google Scholar] [CrossRef]
- Coppa, M.; Gorlier, A.; Lonati, M.; Martin, B.; Russo, E.M.; Lombardi, G. The management of the transition from hay to pasture-based diets affects milk fatty acid kinetics. Dairy Sci. Technol. 2012, 92, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Nudda, A.; Battacone, G.; Boaventura Neto, O.; Cannas, A.; Francesconi, A.H.D.; Atzori, A.S.; Pulina, G. Feeding strategies to design the fatty acid profile of sheep milk and cheese. R. Bras. Zootec. 2014, 43, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Griinari, J.M.; Corl, B.A.; Lacy, S.H.; Chouinard, P.Y.; Nurmela, K.V.V.; Bauman, D.E. Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by Δ9-desaturase. J. Nutr. 2000, 130, 2285–2291. [Google Scholar] [CrossRef]
- Prandini, A.; Sigolo, S.; Cerioli, C.; Piva, G. Survey on conjugated linoleic acid (CLA) content and fatty acid composition of Grana Padano cheese produced in different seasons and areas. Ital. J. Anim. Sci. 2009, 8, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Chion, A.R.; Tabacco, E.; Giaccone, D.; Peiretti, P.G.; Battelli, G.; Borreani, G. Variation of fatty acid and terpene profiles in mountain milk and “Toma piemontese” cheese as affected by diet composition in different seasons. Food Chem. 2010, 121, 393–399. [Google Scholar] [CrossRef]
- Agradi, S.; Curone, G.; Negroni, D.; Vigo, D.; Brecchia, G.; Bronzo, V.; Panseri, S.; Chiesa, L.M.; Peric, T.; Danes, D.; et al. Determination of fatty acids profile in Original Brown cows dairy products and relationship with alpine pasture farming system. Animals 2020, 10, 1231. [Google Scholar] [CrossRef]
- Caredda, M.; Addis, M.; Ibba, I.; Leardi, R.; Scintu, M.F.; Piredda, G.; Sanna, G. Building of prediction models by using Mid-Infrared spectroscopy and fatty acid profile to discriminate the geographical origin of sheep milk. LWT 2017, 75, 131–136. [Google Scholar] [CrossRef]
- Coppa, M.; Farruggia, A.; Ravaglia, P.; Pomiès, D.; Borreani, G.; Le Morvan, A.; Ferlay, A. Frequent moving of grazing dairy cows to new paddocks increases the variability of milk fatty acid composition. Animal 2015, 9, 604–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buccioni, A.; Rapaccini, S.; Antongiovanni, M.; Minieri, S.; Conte, G.; Mele, M. Conjugated linoleic acid and C18:1 isomers content in milk fat of sheep and their transfer to Pecorino Toscano cheese. Int. Dairy J. 2010, 20, 190–194. [Google Scholar] [CrossRef]
- Khan, N.A.; Cone, J.W.; Fievez, V.; Hendriks, W.H. Causes of variation in fatty acid content and composition in grass and maize silages. Anim. Feed Sci. Technol. 2012, 174, 36–45. [Google Scholar] [CrossRef]
- Glasser, F.; Doreau, M.; Maxin, G.; Baumont, R. Fat and fatty acid content and composition of forages: A meta-analysis. Anim. Feed Sci. Technol. 2013, 185, 19–34. [Google Scholar] [CrossRef]
- Cabiddu, A.; Decandia, M.; Addis, M.; Piredda, G.; Pirisi, A.; Molle, G. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 2005, 59, 168–180. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosuè, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Couvreur, S.; Hurtaud, C.; Lopez, C.; Delaby, L.; Peyraud, J.L. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties. J. Dairy Sci. 2006, 89, 1956–1969. [Google Scholar] [CrossRef]
- Marino, V.M.; Schadt, I.; Carpino, S.; Caccamo, M.; La Terra, S.; Guardiano, C.; Licitra, G. Effect of Sicilian pasture feeding management on content of α-tocopherol and β-carotene in cow milk. J. Dairy Sci. 2014, 97, 543–551. [Google Scholar] [CrossRef]
- Di Grigoli, A.; Di Trana, A.; Alabiso, M.; Maniaci, G.; Giorgio, D.; Bonanno, A. Effects of grazing on the behaviour, oxidative and immune status, and production of organic dairy cows. Animals 2019, 9, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Official Methods of Analysis of Aoac International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012.
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- IDF (International Dairy Federation). Cheese and Processed Cheese Product. Determination of the Total Solids Content; Standard FIL-IDF 4A:1982; International Dairy Federation: Brussels, Belgium, 1982. [Google Scholar]
- IDF (International Dairy Federation). Cheese and Processed Cheese Product. Determination of Fat Content-Gravimetric Method (Reference Method); Standard FIL-IDF 5B:1986; International Dairy Federation: Brussels, Belgium, 1986. [Google Scholar]
- IDF (International Dairy Federation). Determination of the Protein Content of Processed Cheese Products; Standard FIL-IDF 25:1964; International Dairy Federation: Brussels, Belgium, 1964. [Google Scholar]
- IDF (International Dairy Federation). Determination of the Ash Content of Processed Cheese Products; Standard FIL-IDF 27:1964; International Dairy Federation: Brussels, Belgium, 1964. [Google Scholar]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanno, A.; Di Grigoli, A.; Vitale, F.; Alabiso, M.; Giosuè, C.; Mazza, F.; Todaro, M. Legume grain-based supplements in dairy sheep diet: Effects on milk yield, composition and fatty acid profile. Anim. Prod. Sci. 2016, 56, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Alabiso, M.; Maniaci, G.; Giosuè, C.; Gaglio, R.; Francesca, N.; Di Grigoli, A.; Portolano, B.; Bonanno, A. Effect of muscle type and animal category on fatty acid composition of bresaola made from meat of Cinisara cattle: Preliminary investigation. CyTA-J. Food 2020, 18, 734–741. [Google Scholar] [CrossRef]
- Ashkezary, M.R.; Bonanno, A.; Todaro, M.; Settanni, L.; Gaglio, R.; Todaro, A.; Alabiso, M.; Maniaci, G.; Mazza, F.; Di Grigoli, A. Effects of adding solid and molten chocolate on the physicochemical, antioxidant, microbiological, and sensory properties of ewe’s milk cheese. J. Food Sci. 2020, 85, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Ulbricth, T.L.V.; Southgate, D.A.T. Coronary Heart Disease, Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- AS/STAT Qualification Tools User’s Guide in SAS 9; Version 9.2; SAS Institute Inc.: Campus Drive Cary, NC, USA, 2010.
- Altomonte, I.; Salari, F.; Neglia, A.; Martini, M. Milk yield and quality characteristics of Cinisara and Modicana cows reared on a farm in the province of Palermo (Sicily-Italy). Large Anim. Rev. 2016, 22, 251–254. [Google Scholar]
- Bonanno, A.; Di Grigoli, A.; Tornambè, G.; Formoso, B.; Alicata, M.L.; Procida, G.; Pizzoferrato, L. Effects of feeding on nutritional and aromatic characteristics of caciocavallo palermitano cheese. In Proceedings of the 6th International Meeting on Mountain Cheese , Ragusa, Italy, 1–6 June 2004. [Google Scholar]
- Todaro, M.; Bonanno, A.; Scatassa, M.L. The quality of Valle del Belice sheep’s milk and cheese produced in the hot summer season in Sicily. Dairy Sci. Technol. 2014, 94, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Masucci, F.; Napolitano, F.; Braghieri, A.; Romano, R.; Manzo, N.; Di Francia, A. Fatty acid and sensory profiles of Caciocavallo cheese as affected by management system. J. Dairy Sci. 2014, 97, 1918–1928. [Google Scholar] [CrossRef] [Green Version]
- Bargo, F.; Delahoy, J.E.; Schroeder, G.F.; Muller, L.D. Milk fatty acid composition of dairy cows grazing at two pasture allowances and supplemented with different levels and sources of concentrate. Anim. Feed Sci. Technol. 2006, 125, 17–31. [Google Scholar] [CrossRef]
- Sinclair, A.J. Dietary fat and cardiovascular disease: The significance of recent developments for the food industry. Food Aust. 1993, 45, 226. [Google Scholar]
- Coppa, M.; Ferlay, A.; Monsallier, F.; Verdier-Metz, I.; Pradel, P.; Didienne, R.; Farruggia, A.; Montel, M.C.; Martin, B. Milk fatty acid composition and cheese texture and appearance from cows fed hay or different grazing systems on upland pastures. J. Dairy Sci. 2011, 94, 1132–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreu-Coll, L.; Cano-Lamadrid, M.; Sendra, E.; Carbonell-Barrachina, A.; Legua, P.; Hernández, F. Fatty acid profile of fruits (pulp and peel) and cladodes (young and old) of prickly pear Opuntia ficus-indica (L.) Mill. from six Spanish cultivars. J. Food Compost. Anal. 2019, 84, 103294. [Google Scholar] [CrossRef]
- El Otmani, S.; Chebli, Y.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Effects of Olive Cake and Cactus Cladodes as Alternative Feed Resources on Goat Milk Production and Quality. Agriculture 2021, 11, 3. [Google Scholar] [CrossRef]
- Szumacher-Strabel, M.; Cieslak, A.; Nowakowska, A. Effect of oils rich in linoleic acid on in vitro rumen fermentation parameters of sheep, goats and dairy cows. J. Anim. Feed Sci. 2009, 18, 440–452. [Google Scholar] [CrossRef]
- Collomb, M.; Bisig, W.; Butikofer, U.; Sieber, R.; Bregy, M.; Etter, L. Fatty acid composition of mountain milk from Switzerland: Comparison of organic and integrated farming systems. Int. Dairy J. 2008, 18, 976–982. [Google Scholar] [CrossRef]
- Ferlay, A.; Martin, B.; Pradel, P.; Coulon, J.B.; Chilliard, Y. Influence of grass-based diets on milk fatty acid composition and milk lipolytic system in Tarentaise and Montbe´liarde cow breeds. J. Dairy Sci. 2006, 89, 4026–4041. [Google Scholar] [CrossRef] [Green Version]
- Milewski, S.; Ząbek, K.; Antoszkiewicz, Z.; Tański, Z.; Sobczak, A. Impact of production season on the chemical composition and health properties of goat milk and rennet cheese. Emir. J. Food Agric. 2018, 107–114. [Google Scholar] [CrossRef]
- Romanzin, A.; Corazzin, M.; Piasentier, E.; Bovolenta, S. Effect of rearing system (mountain pasture vs. indoor) of Simmental cows on milk composition and Montasio cheese characteristics. J. Dairy Res. 2013, 80, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Mele, M. Designing milk fat to improve healthfulness and functional properties of dairy products: From feeding strategies to a genetic approach. Ital. J. Anim. Sci. 2009, 8, 365–374. [Google Scholar] [CrossRef]
- Fieser, B.G.; Vanzant, E.S. Interactions between supplement energy source and tall fescue hay maturity on forage utilization by beef steers. J. Anim. Sci. 2004, 82, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Ba, N.X.; Van Huu, N.; Ngoan, L.D.; Leddin, C.M.; Doyle, P.T. Effects of amount of concentrate supplement on forage intake, diet digestibility and live weight gain in yellow cattle in Vietnam. Asian-Australas. J. Anim. Sci. 2008, 21, 1736–1744. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Season | Autumn | Winter | Spring | Summer | ||||
|---|---|---|---|---|---|---|---|---|
| Farm | A | B | A | B | A | B | A | B |
| Farm characteristics | ||||||||
| Available grazing areas, ha | 26 | 30 | 26 | 30 | 26 | 30 | 26 | 30 |
| Lactating cows, n. | 15 | 17 | 18 | 25 | 18 | 26 | 10 | 15 |
| Indoor diet ingredients, kg/animal per day | ||||||||
| Wheat bran | 7.0 | 7.0 | 7.8 | 7.9 | 4.9 | 4.9 | 5.6 | 5.0 |
| Wheat straw | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| Opuntia ficus-indica cladodes | 9.0 | 11.0 | ||||||
| Pastures | Feed | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Autumn | Winter | Spring | Summer | Wheat Bran | Wheat Straw | Cladodes 1 | ||||
| Farm | A | B | A | B | A | B | A | B | |||
| Dry Matter, % | 19.16 | 18.70 | 16.26 | 15.52 | 18.79 | 19.66 | 78.15 | 76.32 | 89.54 | 92.71 | 8.64 |
| Crude Protein | 19.44 | 23.37 | 21.32 | 23.75 | 17.93 | 16.12 | 9.94 | 9.87 | 18.11 | 6.37 | 8.31 |
| Ether extract | 3.62 | 4.30 | 5.00 | 4.44 | 3.87 | 3.35 | 8.81 | 7.07 | 4.65 | 1.73 | 2.41 |
| Ash | 11.86 | 14.75 | 12.40 | 13.87 | 9.32 | 10.11 | 8.94 | 10.58 | 4.39 | 6.96 | 27.35 |
| NDF | 49.71 | 47.88 | 38.45 | 35.61 | 44.48 | 40.89 | 51.59 | 53.27 | 26.22 | 73.22 | 31.08 |
| Total FA, % DM | 3.23 | 3.85 | 4.50 | 4.22 | 3.44 | 2.85 | 7.98 | 6.41 | 3.86 | 1.56 | 2.15 |
| C12:0 | 0.53 | 0.49 | 0.57 | 0.53 | 0.44 | 0.42 | 0.41 | 0.56 | 0.00 | 2.14 | 0.00 |
| C14:0 | 2.17 | 2.23 | 2.21 | 2.61 | 1.80 | 1.91 | 0.85 | 0.73 | 10.37 | 4.55 | 0.00 |
| C14:1 | 0.38 | 0.37 | 0.41 | 0.4 | 0.35 | 0.31 | 0.3 | 0.31 | 0.00 | 0.00 | 0.00 |
| C16:0 | 17.60 | 17.70 | 15.62 | 15.32 | 20.1 | 21.54 | 27.71 | 27.92 | 14.45 | 32.74 | 17.43 |
| C16:1 | 1.40 | 1.51 | 1.45 | 1.66 | 1.46 | 1.05 | 1.41 | 1.41 | 0.21 | 0.00 | 0.00 |
| C18:0 | 2.38 | 2.43 | 1.96 | 1.76 | 3.58 | 2.79 | 6.90 | 7.01 | 0.79 | 18.10 | 5.04 |
| C18:1 c9 | 2.79 | 2.95 | 2.25 | 2.75 | 3.17 | 3.01 | 18.04 | 17.85 | 16.65 | 7.52 | 11.14 |
| C18:2 n-6 | 15.77 | 15.81 | 12.21 | 12.01 | 22.70 | 20.95 | 35.18 | 35.02 | 51.71 | 11.31 | 35.40 |
| C18:3 n-3 | 56.75 | 56.91 | 62.79 | 62.73 | 46.30 | 47.90 | 9.15 | 9.19 | 5.81 | 23.64 | 16.20 |
| C20:0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 14.70 |
| Season 1 | SEM 2 | Significance | ||||
|---|---|---|---|---|---|---|
| Parameters | Autumn | Winter | Spring | Summer | Season | |
| Milk | ||||||
| Daily yield, kg/head | 11.7 a | 11.5 a | 12.7 a | 9.8 b | 0.635 | 0.035 |
| Fat | 3.42 ABa | 3.52 Aa | 3.12 Bb | 3.14 Bb | 0.081 | 0.005 |
| Protein | 3.46 ABb | 3.70 Aa | 3.59 Aab | 3.27 Bc | 0.063 | 0.002 |
| Casein | 2.93 ABb | 3.22 Aa | 2.94 Ab | 2.66 Bc | 0.073 | 0.001 |
| Lactose | 4.60 b | 4.73 a | 4.60 b | 4.65 ab | 0.029 | 0.024 |
| Cheese | ||||||
| Yield at 24 h, kg/100 kg | 11.4 ABa | 11.8 Aa | 10.8 BCb | 10.2 Cb | 0.195 | <0.001 |
| Dry Matter, % | 65.3 | 65.1 | 67.5 | 70.4 | 1.543 | 0.096 |
| Fat | 45.3 Aa | 44.9 ABa | 41.0 Bb | 44.2 ABa | 0.749 | 0.002 |
| Protein | 44.6 Bb | 46.7 ABab | 48.4 Aa | 45.1 ABb | 0.856 | 0.007 |
| Ash | 8.02 a | 6.46 b | 7.89 b | 7.84 b | 0.308 | 0.017 |
| Season 1 | SEM 2 | Significance | ||||
|---|---|---|---|---|---|---|
| Parameters | Autumn | Winter | Spring | Summer | Season | |
| C4:0 | 2.54 a | 2.54 a | 1.59 b | 2.29 ab | 0.249 | 0.019 |
| C6:0 | 2.03 | 2.32 | 1.76 | 1.80 | 0.146 | 0.072 |
| C8:0 | 1.27 | 1.58 | 0.81 | 1.11 | 0.252 | 0.214 |
| C10:0 | 2.58 | 3.41 | 2.36 | 2.21 | 0.327 | 0.131 |
| C12:0 | 2.93 | 3.92 | 2.13 | 2.46 | 0.572 | 0.214 |
| C14:0 | 11.0 B | 12.9 A | 10.9 B | 10.4 B | 0.457 | <0.001 |
| C14:1 | 0.96 AB | 1.12 A | 0.93 AB | 0.76 B | 0.062 | 0.012 |
| C15:0iso | 0.34 | 0.32 | 0.32 | 0.32 | 0.009 | 0.385 |
| C15:0 | 1.19 | 1.24 | 1.35 | 1.26 | 0.047 | 0.073 |
| C16:0iso | 0.32 | 0.30 | 0.31 | 0.28 | 0.024 | 0.638 |
| C16:0 | 28.4 AB | 31.7 A | 29.0 AB | 27.0 B | 0.789 | 0.011 |
| C16:1 | 0.22 ab | 0.19 b | 0.33 a | 0.24 ab | 0.034 | 0.025 |
| C17:0iso | 0.51 ab | 0.45 b | 0.54 a | 0.54 a | 0.021 | 0.047 |
| C17:0anteiso | 1.83 | 1.86 | 1.68 | 1.82 | 0.080 | 0.303 |
| C17:0 | 0.72 | 0.72 | 0.79 | 0.79 | 0.019 | 0.060 |
| C17:1 | 0.25 ab | 0.22 b | 0.25 ab | 0.29 a | 0.015 | 0.007 |
| C18:0 | 10.3 AB | 8.56 B | 12.4 A | 12.2 A | 0.530 | 0.003 |
| C18:1 c9 OA | 22.9 ABab | 18.4 Bc | 19.8 Bbc | 24.2 Aa | 0.981 | 0.002 |
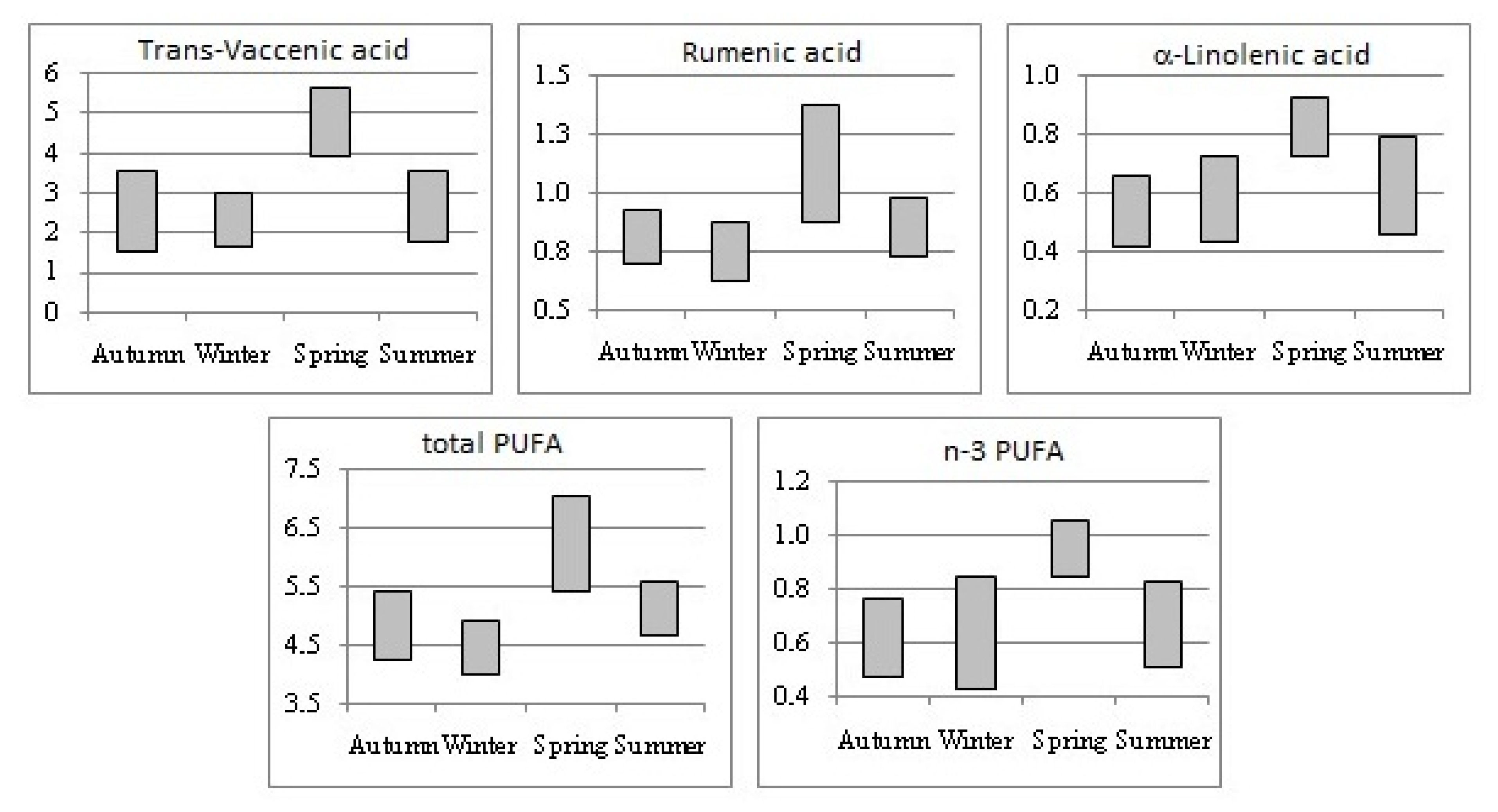
| C18:1 t11 TVA | 3.15 B | 2.82 B | 4.14 A | 3.09 B | 0.218 | <0.001 |
| Other C18:1 | 1.37 ABb | 0.89 Bb | 2.12 Aa | 1.39 ABb | 0.174 | <0.001 |
| Other C18:2 | 0.75 | 0.62 | 0.81 | 0.60 | 0.092 | 0.070 |
| C18:2 n-6 LA | 2.18 | 2.01 | 2.52 | 2.54 | 0.179 | 0.143 |
| CLA C18:2 c9t11 RA | 0.93 b | 0.74 b | 1.24 a | 0.82 b | 0.083 | 0.010 |
| Other CLA c isomers | 0.10 AB | 0.04 B | 0.13 A | 0.06 AB | 0.021 | 0.011 |
| C18:3 n-3 ALA | 0.55 B | 0.50 B | 0.84 A | 0.58 B | 0.049 | <0.001 |
| C18:3 n-6 GLA | 0.15 ABb | 0.12 Bb | 0.18 Aa | 0.13 Bb | 0.007 | <0.001 |
| C20:0 | 0.19 B | 0.16 B | 0.20 B | 0.27 A | 0.010 | <0.001 |
| C20:2 n-6 | 0.04 | 0.03 | 0.04 | 0.05 | 0.010 | 0.719 |
| C20:3 n-6 | 0.07 | 0.07 | 0.10 | 0.08 | 0.008 | 0.078 |
| C20:4 n-6 AA | 0.14 | 0.14 | 0.15 | 0.15 | 0.008 | 0.439 |
| C20:5 n-3 EPA | 0.02 | 0.02 | 0.05 | 0.05 | 0.008 | 0.099 |
| C22:0 | 0.07 | 0.06 | 0.08 | 0.08 | 0.008 | 0.059 |
| C22:5 n-3 DPA | 0.04 | 0.05 | 0.08 | 0.07 | 0.012 | 0.070 |
| Season 1 | SEM 2 | Significance | ||||
|---|---|---|---|---|---|---|
| Parameters | Autumn | Winter | Spring | Summer | Season | |
| Total FA, % DM | 41.0 Aa | 40.6 ABa | 37.0 Bb | 40.0 ABa | 0.711 | <0.001 |
| SFA 3 | 66.2 ABb | 72.1 Aa | 66.3 ABb | 64.9 Bb | 1.166 | 0.006 |
| MUFA 4 | 28.8 ABa | 23.6 Bb | 27.5 ABab | 30.0 Aa | 0.984 | 0.004 |
| PUFA 5 | 4.94 Bb | 4.31 Bb | 6.19 Aa | 5.16 ABb | 0.242 | <0.001 |
| MUFA/SFA | 0.44 ABa | 0.33 Bb | 0.42 ABab | 0.46 Aa | 0.022 | 0.007 |
| PUFA/SFA | 0.07 ab | 0.06 b | 0.09 a | 0.08 ab | 0.005 | 0.021 |
| n-6 | 2.57 | 2.37 | 2.94 | 2.99 | 0.200 | 0.134 |
| n-3 | 0.59 B | 0.56 B | 0.97 A | 0.70 B | 0.053 | <0.001 |
| n-6/n-3 | 4.46 | 4.38 | 3.06 | 4.44 | 0.477 | 0.071 |
| HPI 6 | 0.44 Aa | 0.32 Bc | 0.38 Bb | 0.47 Aa | 0.028 | 0.002 |
| TI 7 | 2.79 B | 3.42 A | 2.84 B | 2.67 B | 0.091 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maniaci, G.; Di Grigoli, A.; Bonanno, A.; Giosuè, C.; Ilardi, V.; Alabiso, M. Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese. Animals 2021, 11, 2675. https://doi.org/10.3390/ani11092675
Maniaci G, Di Grigoli A, Bonanno A, Giosuè C, Ilardi V, Alabiso M. Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese. Animals. 2021; 11(9):2675. https://doi.org/10.3390/ani11092675
Chicago/Turabian StyleManiaci, Giuseppe, Antonino Di Grigoli, Adriana Bonanno, Cristina Giosuè, Vincenzo Ilardi, and Marco Alabiso. 2021. "Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese" Animals 11, no. 9: 2675. https://doi.org/10.3390/ani11092675