
Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares


Abstract
:Simple Summary
Abstract
1. Introduction
2. Susceptibility and Resistance to Endometritis
3. Post Breeding Endometritis
4. Delayed Uterine Clearance
4.1. Risk Factors
4.2. Intrauterine Fluid
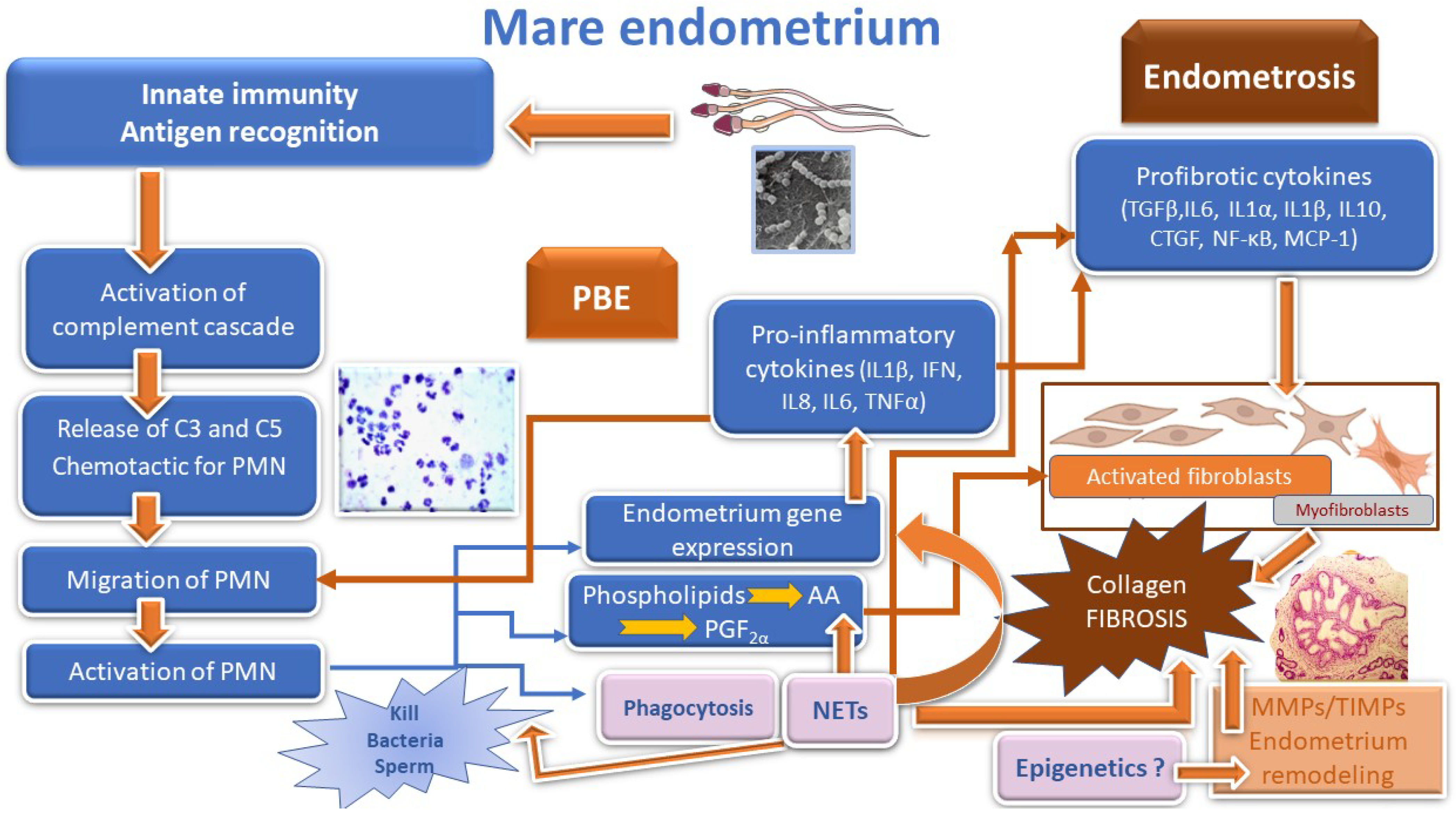
5. Physiology of Post Breeding Endometritis
Cytokines
6. Treatments
7. Etiology of Prolonged Post Breeding Endometritis
Occlusion of the Cervix
8. Endometrosis
8.1. Endometritis and Endometrosis Are Interconnected
8.2. Pathogenesis of Endometrosis
8.2.1. Cytokines and Other Fibrosis Mediators
8.2.2. Neutrophils and NETs
8.2.3. Prostaglandins
9. Epigenetics
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Hughes, J.P.; Loy, R.G. Investigations on the effect of intrauterine inoculations of Streptococcus zooepidemicus in the mare. Proc. Am. Assoc. Equine Pract. 1969, 15, 289–292. [Google Scholar]
- Dimock, W.W.; Snyder, E. Bacteria of the genital tract of mares and the semen of stallions and their relation to breeding efficiency. J. Am. Vet. Med. Assoc. 1924, 64, 288–297. [Google Scholar]
- Kenney, R.M. Cyclic and pathologic changes of the mare endometrium as detected by biopsy, with a note on early embryonic death. J. Am. Vet. Med. Assoc. 1978, 172, 241–262. [Google Scholar] [PubMed]
- Kenney, R.M.; Doig, P.A. Equine endometrial biopsy. In Current Therapy in Theriogenology; Morrow, D.A., Ed.; W.B. Saunders: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Katila, T.; Lock, T.F.; Hoffman, W.E.; Smith, A.R. Lysozyme, alkaline phosphatase, and neutrophils in uterine secretions of mares with differing resistance to endometritis. Theriogenology 1990, 33, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, M.H.T.; Liu, I.K.M. Measurement of total volume and protein concentration of intrauterine secretion after intrauterine inoculation of bacteria in mares that were either resistant or susceptible to chronic uterine infection. Am. J. Vet. Res. 1992, 53, 1641–1644. [Google Scholar]
- Nikolakopoulos, E.; Watson, E.D. Does artificial insemination with chilled, extended semen reduce the antigenic challenge to the mare’s uterus compared with natural service? Theriogenology 1997, 47, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Squires, E.L.; Troedsson, M.H.T. Susceptibility to persistent breeding-induced endometritis in the mare: Relationship to endometrial biopsy score and age, and variations between seasons. Theriogenology 2012, 78, 495–501. [Google Scholar] [CrossRef]
- Troedsson, M.H.T.; deMoraes, M.J.; Liu, I.K.M. Correlations between histologic endometrial lesions in mares and clinical response to intrauterine exposure with Streptococcus zooepidemicus. Am. J. Vet. Res. 1993, 54, 570–572. [Google Scholar]
- Troedsson, M.H.T.; Liu, I.K.M. Uterine clearance of non-antigenic markers (51Cr) in response to a bacterial challenge in mares potentially susceptible and resistant to chronic uterine infection. J. Reprod. Fertil. 1991, 44, 307–313. [Google Scholar] [PubMed]
- Fumuso, E.A.; Aguilar, J.; Giguère, S.; Rivulgo, M.; Wade, J.; Rogan, D. Immune parameters in mares resistant and susceptible to persistent post-breeding endometritis: Effects of immunomodulation. Vet. Immunol. Immunopathol. 2007, 118, 30–39. [Google Scholar] [CrossRef]
- LeBlanc, M.M.; Neuwirth, L.; Asbury, A.C.; Tran, T.; Mauragis, D.; Klapstein, E. Scintigraphic measurement of uterine clearance in normal mares and mares with recurrent endometritis. Equine Vet. J. 1994, 26, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Pierson, R.A. Ultrasonic anatomy and pathology of the equine uterus. Theriogenology 1984, 21, 505–516. [Google Scholar] [CrossRef]
- Pycock, J.F.; Newcombe, J.R. Assessment of the effect of three treatments to remove intrauterine fluid on pregnancy rate in the mare. Vet. Rec. 1996, 138, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Brinsko, S.P.; Rigby, S.L.; Varner, D.D. A practical method for recognizing mares susceptible to post-breeding endometritis. In Proceedings of the American Association Equine Practitioners, New Orleans, LA, USA, 21–25 November 2003; Volume 49, pp. 363–365.
- Cásseres de Borba, E.V.; Casanova Comozzato, G.; Malschitzky, E.; Bustamante-Filho, I.; Afonso Martins, A.; Costa Mattos, R.; Pires Neves, A. Is the presence of uterine fluid a reliable indicator of endometrial inflammation? Pferdeheilkunde 2012, 28, 27–29. [Google Scholar] [CrossRef] [Green Version]
- Zent, W.W.; Troedsson, M.H.T.; Xue, J.-L. Postbreeding uterine fluid accumulation in a normal population of thoroughbred mares: A field study. In Proceedings of the 44th America Association Equine Practitioners, Baltimore, MD, USA, 9 December 1998; Volume 44, pp. 64–65. [Google Scholar]
- Dimock, W.W.; Edwards, P.R. The pathology and bacteriology of the reproductive organs of mares in relation to sterility. Ky. Agric. Exp. Stn. Bull. 1928, 286, 157–237. [Google Scholar]
- Kotilainen, T.; Huhtinen, M.; Katila, T. Sperm-induced leukocytosis in the equine uterus. Theriogenology 1994, 41, 629–636. [Google Scholar] [CrossRef]
- Katila, T. Onset and duration of uterine inflammatory response of mares after insemination with fresh semen. Biol. Reprod. Mono 1995, 52, 515–517. [Google Scholar] [CrossRef]
- Troedsson, M.H.T. Uterine clearance and resistance to persistent endometritis in the mare. Theriogenology 1999, 52, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Katila, T. Evaluation of diagnostic methods in equine endometritis. Reprod. Biol. 2016, 16, 189–196. [Google Scholar] [CrossRef]
- Reilas, T.; Rivera del Alamo, M.M.; Liepina, E.; Yeste, M.; Katila, T. Effects on the equine endometrium of cervical occlusion after insemination. Theriogenology 2016, 85, 617–624. [Google Scholar] [CrossRef]
- Morris, L.H.A.; McCue, P.M.; Aurich, C. Equine endometritis: A review of challenges and new approaches. Reproduction 2020, 160, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Rasch, K.; Schoon, H.A.; Sieme, H.; Klug, E. Histomorphological endometrial status and influence of oxytocin on the uterine drainage and pregnancy rate in mares. Equine Vet. J. 1996, 28, 455–460. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, M.M. Advances in the diagnosis and treatment of chronic infectious and post-mating-induced endometritis in the mare. Reprod. Domest. Anim. 2010, 45, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Hamer, J.M.; Gason, L.M.; Graham, C.S.; Asbury, A.C.; Irvine, C.H. Clearance of bacteria and non-antigenic markers following intra-uterine inoculation into maiden mares: Effect of steroid hormone environment. Theriogenology 1986, 26, 37–50. [Google Scholar] [CrossRef]
- Evans, M.J.; Hamer, J.M.; Gason, L.M. Factors affecting uterine clearance of inoculated materials in mares. J. Reprod. Fertil. 1987, 35, 327–334. [Google Scholar] [PubMed]
- Caslick, E.A. The vulva and the vulvovaginal orifice and its relation to genital health of the thoroughbred mare. Cornell Vet. 1937, 27, 178–187. [Google Scholar]
- LeBlanc, M.M.; Neuwirth, L.; Jones, L.; Mauragis, D. Differences in uterine position of reproductively normal mares and those with delayed uterine clearance detected by scintigraphy. Theriogenology 1998, 50, 49–54. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, M.L.; Schlafer, D.H. Selected topics in reproductive pathology: Mare II. In Proceedings of the 61st Annual Convention of the American Association of Equine Practitioners, Las Vegas, NV, USA, 5–9 December 2015; pp. 276–296. [Google Scholar]
- Knudsen, O. Partial dilatation of the uterus as a cause of sterility in the mare. Cornell Vet. 1964, 54, 423–438. [Google Scholar] [PubMed]
- Reilas, T.; Katila, T.; Mäkelä, O.; Huhtinen, M.; Koskinen, E. Intrauterine fluid accumulation in oestrous mares. Acta Vet. Scand. 1997, 38, 69–78. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, M.M.; Johnson, R.D.; Calderwood-Mays, M.B.; Valderrama, C. Lymphatic clearance of India ink in reproductively normal mares and mares susceptible to endometritis. Biol. Reprod. Monogr. 1 1995, 52, 109–113. [Google Scholar]
- Özgen, S.; Schoon, H.-A.; Aupperle, H.; Sieme, H.; Klug, E. Etiopathogenesis of equine intrauterine fluid accumulation. Pferdeheilkunde 2002, 18, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Ricketts, S.W.; Alonso, S. The effect of age and parity on the development of equine chronic endometrial disease. Equine Vet. J. 1991, 23, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, E.M.; Ginther, O.J. Relationships of age to uterine function and reproductive efficiency in mares. Theriogenology 1992, 37, 1101–1115. [Google Scholar] [CrossRef] [PubMed]
- Barbacini, S.; Necchi, D.; Zavaglia, G.; Squires, E.L. Retrospective study on the incidence of postinsemination uterine fluid in mares inseminated with frozen/thawed semen. J. Equine Vet. Sci. 2003, 23, 493–496. [Google Scholar] [CrossRef]
- Sinnemaa, L.; Järvimaa, T.; Lehmonen, N.; Mäkelä, O.; Reilas, T.; Sankari, S.; Katila, T. Effect of insemination volume on uterine contractions and inflammatory response and on elimination of semen in the mare uterus–scintigraphic and ultrasonographic studies. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2005, 52, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Nikolakopoulos, E.; Kindahl, H.; Watson, E.D. Oxytocin and PGF2alpha release in mares resistant and susceptible to persistent mating-induced endometritis. J. Reprod. Fertil. 2000, 56, 363–372. [Google Scholar] [PubMed]
- Troedsson, M.H.T.; Liu, I.K.M.; Ing, M.; Pascoe, J.; Thurmond, M. Multiple site electromyography recordings of uterine activity following and intrauterine bacterial challenge in mares susceptible and resistant to chronic uterine infection. J. Reprod. Fertil. 1993, 99, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, A.S.; Foster, D.N.; Carlson, C.S.; Troedsson, M.H. Nitric oxide levels and nitric oxide synthase expression in uterine samples from mares susceptible and resistant to persistent breeding-induced endometritis. Am. J. Reprod. Immunol. 2005, 53, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Horohov, D.W.; Scoggin, K.E.; Squires, E.; Troedsson, M.H.T. An investigation of uterine nitric oxide production in mares susceptible and resistant to persistent breeding induced endometritis, and the effects of immunomodulation. Reprod. Domest. Anim. 2013, 48, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, M.H.T. Problems after breeding. J. Equine Vet. Sci. 2008, 28, 635–639. [Google Scholar] [CrossRef]
- LeBlanc, M.M.; Causey, R.C. Clinical and subclinical endometritis in the mare: Both threats to fertility. Reprod. Dom. Anim. 2009, 44, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Malschitzky, E.; Rodrigues Trein, C.; Cunha Bustamante Filho, I.; Garbade, P.; Macedo Gregory, R.; Costa Mattos, R. Young maiden mares can also be susceptible to a persistent mating-induced endometritis. Pferdeheilkunde 2006, 22, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Tunòn, A.-M.; Ekwall, H.; Nummijärvi, A.; Rodriguez-Martinez, H. X-ray microanalysis of the secretory epithelium of the endometrial glands and intraluminal uterine fluid in oestrus mares. Reprod. Domest. Anim. 2000, 35, 221–227. [Google Scholar] [CrossRef]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, E. Persistent breeding-induced endometritis in mares–a multifaceted challenge: From clinical aspects to immunopathogenesis and pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elweza, A.E.; Ezz, M.A.; Acosta, T.J.; Talukder, A.K.; Shimizu, T.; Hayakawa, H.; Shimada, M.; Imakawa, K.; Zaghlouls, A.H.; Miyamoto, A. A proinflammatory response of bovine endometrial epithelial cells to active sperm in vitro. Mol. Reprod. Dev. 2018, 85, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Palm, F.; Walter, I.; Budik, S.; Kolodziejek, J.; Nowotny, N.; Aurich, C. Influence of different semen extenders and seminal plasma on PMN migration and on expression of IL-1β, IL-6, TNF-αand COX-2 mRNA in the equine endometrium. Theriogenology 2008, 70, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Nash, D.M.; Sheldon, I.M.; Herath, S.; Lane, E.A. Markers of the uterine innate immune response of the mare. Anim. Reprod. Sci. 2010, 119, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Pereira, C.; Galvão, A.; Pinto Bravo, P.; Szóstek, A.; Skarzynski, D.J.; Ferreira-Dias, G. What is new on the physiopathology of endometrosis in the mare? Pferdeheilkunde 2014, 30, 15–18. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Skarzynski, D.J.; Szóstek-Mioduchowska, A.Z.; Rebordão, M.R.; Jalali, B.M.; Piotrowska-Tomala, K.K.; Leciejewska, N.; Łazarczyk, M.; Ferreira-Dias, G.M. Neutrophils, monocytes and other immune components in the equine endometrium: Friends or foes? Theriogenology 2020, 150, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Woodward, E.M.; Troedsson, M.H.T. Inflammatory mechanisms of endometritis. Equine Vet. J. 2015, 47, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Betancourt, A.; Horohov, D.W.; Scoggin, K.E.; Squires, E.; Troedsson, M.H.T. Endometrial inflammatory markers of the early immune response in mares susceptible or resistant to persistent breeding-induced endometritis. Reproduction 2013, 145, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, W.E. Investigations into the use of exogenous oxytocin for promoting uterine drainage in mares susceptible to endometritis. Vet. Rec. 1991, 128, 593–594. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, M.H.; Scott, M.A.; Liu, I.K. Comparative treatment of mares susceptible to chronic uterine infection. Am. J. Vet. Res. 1995, 56, 468–472. [Google Scholar] [PubMed]
- Causey, R.C. Mucus and the mare: How little we know. Theriogenology 2007, 68, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Dell Aqua, J.; Papa, F.; Araùjo, J.; Alvarenga, M.; Zahn, F.; Lopes, M. Modulation of acute uterine inflammatory response after artificial insemination with equine frozen semen. Anim. Reprod. Sci. 2006, 94, 270–273. [Google Scholar]
- Bucca, S.; Carli, A.; Buckley, T.; Dolci, G.; Fogarty, U. The use of dexamethasone administered to mares at breeding time in the modulation of persistent mating induced endometritis. Theriogenology 2008, 70, 1093–1100. [Google Scholar] [CrossRef]
- Christoffersen, M.; Woodward, E.M.; Bojesen, A.M.; Petersen, M.R.; Squires, E.L.; Lehn-Jensen, H.; Troedsson, M.H.T. Effect of immunomodulatory therapy on the endometrial inflammatory response to induced infectious endometritis in susceptible mares. Theriogenology 2012, 78, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Woodward, E.M.; Christoffersen, M.; Horohov, D.W.; Squires, E.; Troedsson, M.H.T. The effect of treatment with immune modulators on endometrial cytokine expression in mares susceptible to persistent breeding-induced endometritis. Equine Vet. J. 2015, 47, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Güvenc, K.; Reilas, T.; Katila, T. Effect of insemination dose and site on uterine inflammatory response of mares. Theriogenology 2005, 63, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Güvenc, K.; Reilas, T.; Katila, T. Effect of frozen semen on the uterus of mares with pathological uterine changes. Reprod. Nutr. Dev. 2004, 44, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, R.M. Chronic degenerative endometritis (CDE) (endometrosis). Equine Vet. J. 1993, 25, 184–193. [Google Scholar] [CrossRef]
- Schöniger, S.; Schoon, H.-A. The healthy and diseased equine endometrium: A review of morphological features and molecular analyses. Animals 2020, 10, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteller-Vico, A.; Liu, I.K.; Brosnan, R.J. Uterine vascular elastosis, endometrial grade, age and parity. Theriogenology 2010, 121, 87–88. [Google Scholar]
- Esteller-Vico, A.; Liu, I.K.; Vaughan, B.; Steffey, E.P.; Brosnan, R.J. Effects of vascular elastosis on uterine blood flow and perfusion in anesthetized mares. Theriogenology 2015, 83, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Esteller-Vico, A.; Liu, I.K.; Couto, S. Uterine vascular degeneration is present throughout the uterine wall of multiparous mares. Colinearity between elastosis, endometrial grade, age and parity. Theriogenology 2012, 78, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A. Tissue injury and repair in the female human reproductive tract. Reproduction 2003, 125, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Diel de Amorim, M.; Khan, F.A.; Chenier, T.S.; Scholtz, E.L.; Hayes, M.A. Analysis of the uterine flush fluid proteome of healthy mares and mares with endometritis or fibrotic endometrial degeneration. Reprod. Fertil. Dev. 2020, 32, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.A.; Bartol, F.F.; Tarleton, B.J.; Wiley, A.A.; Johnson, G.A.; Bazer, F.W.; Spencer, T.E. Developmental biology of uterine glands. Biol. Reprod. 2001, 65, 1311–1323. [Google Scholar] [CrossRef]
- Allen, W.R.; Wilsher, S. A review of implantation and early placentation in the mare. Placenta 2009, 30, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Ellenberger, C.; Mattos, R.C.; Aupperle, H.; Dhein, S.; Stief, B.; Schoon, H.A. The equine endometrosis: New insights into the pathogenesis. Anim. Reprod. Sci. 2009, 111, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Aresu, L.; Benali, S.; Giannuzzi, D.; Mantovani, R.; Castagnaro, M.; Falomo, M.E. The role of inflammation and matrix metalloproteinases in equine endometrosis. J. Vet. Sci. 2012, 13, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanada, M.; Maeda, Y.; Oikawa, M.A. Histopathological characteristics of endometrosis in thoroughbred mares in Japan: Results from 50 necropsy cases. J. Equine Sci. 2014, 25, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Alpoim-Moreira, J.; Fernandes, C.; Rebordão, M.R.; Amaral, A.; Pinto-Bravo, P.; Bliebernicht, M.; Skarzynski, D.J.; Ferreira-Dias, G. Collagens and DNA methyltransferases in mare endometrosis. Reprod. Domest. Anim. 2019, 54, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crociati, M.; Capomaccio, S.; Mandara, M.T.; Stradaioli, G.; Sylla, L.; Monaci, M.; Cappelli, K. Different expression of Defensin-B gene in the endometrium of mares of different age during the breeding season. BMC Vet. Res. 2019, 15, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto-Bravo, P.; Rebordão, M.R.; Amaral, A.; Fernandes, C.; Cuello, C.; Parrilla, I.; Martínez, E.; Roberto da Costa, R.P.; Skarzynski, D.J.; Ferreira-Dias, G. Is mare endometrosis linked to oviduct fibrosis? Pferdeheilkunde 2018, 34, 43–46. [Google Scholar] [CrossRef]
- Bracher, V.; Mathias, S.; Allen, W.R. Influence of chronic degenerative endometritis (endometrosis) on placental development in the mare. Equine Vet. J. 1996, 28, 180–188. [Google Scholar] [CrossRef]
- Wilsher, S.; Allen, W.R. Factors influencing placental development and function in the mare. Equine Vet. J. 2012, 41, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Neto da Silva, A.C.; Costa, A.L.; Teixeira, A.; Alpoim-Moreira, J.; Fernandes, C.; Fradinho, M.J.; Rebordão, M.R.; Silva, E.; Ferreira da Silva, J.; Bliebernicht, M.; et al. Collagen and microvascularization in placentas from young and older mares. Front. Vet. Sci. 2022, 8, 772658. [Google Scholar] [CrossRef]
- Wilsher, S.; Allen, W.R. The effects of maternal age and parity on placental and fetal development in the mare. Equine Vet. J. 2003, 35, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szóstek-Mioduchowska, A.Z.; Lukasik, K.; Skarzynski, D.J.; Okuda, K. Effect of transforming growth factor-β1 on α-smooth muscle actin and collagen expression in equine endometrial fibroblasts. Theriogenology 2019, 15, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.Z.; Baclawska, A.; Okuda, K.; Skarzynski, D.J. Effect of proinflammatory cytokines on endometrial collagen and metallopeptidase expression during the course of equine endometrosis. Cytokine 2019, 123, 154767. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.; Słowinska, M.; Pacewicz, J.; Skarzynski, D.J.; Okuda, K. Matrix metallopeptidase expression and modulation by transforming growth factor-β1 in equine endometrosis. Sci. Rep. 2020, 10, 1119. [Google Scholar] [CrossRef] [Green Version]
- Szóstek, A.Z.; Lukasik, K.; Galvão, A.M.; Ferreira-Dias, G.M.; Skarzynski, D.J. Impairment of the interleukin system in equine endometrium during the course of endometrosis. Biol. Reprod. 2013, 89, 1–13. [Google Scholar] [CrossRef]
- de Holanda, A.G.B.; da Silva Leite, J.; Consalter, A.; da Silva, K.V.G.C.; Dos Santos Batista, B.P.; Fonseca, A.B.M.; Brandão, F.Z.; Ferreira, A.M.R. Expression of interleukins 6 and 10 and population of inflammatory cells in the equine endometrium: Diagnostic implications. Mol. Biol. Rep. 2019, 46, 2485–2491. [Google Scholar] [CrossRef]
- Moolenaar, W.H.; Van Meeteren, L.A.; Giepmans, B.N.G. The ins and outs of lysophosphatidic acid signaling. Bioessays 2004, 26, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.; Leciejewska, N.; Zelmańska, B.; Staszkiewicz-Chodor, J.; Ferreira-Dias, G.; Skarzynski, D. Lysophosphatidic acid as a regulator of endometrial connective tissue growth factor and prostaglandin secretion during estrous cycle and endometrosis in the mare. BMC Vet. Res. 2020, 16, 343. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Chun, J.; Duffield, J.S.; Wada, T.; Luster, A.D.; Tager, A.M. LPA1-induced cytoskeleton reorganization drives fibrosis through CTGF-dependent fibroblast proliferation. FASEB J. 2013, 27, 1830–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domino, M.; Jasinski, T.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Zabielski, R.; Sady, M.; Gajewski, Z. Expression of genes involved in the NF-κB-dependent pathway of the fibrosis in the mare endometrium. Theriogenology 2020, 147, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Jasiński, T.; Zdrojkowski, Ł.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Domino, M. Equine endometrosis pathological features: Are they dependent on NF-κB Signaling pathway? Animals 2021, 11, 3151. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V. Neutrophil extracellular traps in microbial infections. In Neutrophils in Infectious Diseases; Tacchini-Cottier, F., Zandbergen, G., Eds.; Bentham eBooks: Dubais, United Arab Emirates, 2011; pp. 3–10. [Google Scholar]
- Souza, D.G.; Alves-Filho, J.C.; Cunha, F.Q.; Lima, C.X.; Teixeira, M.M. Neutrophils in the context of polymicrobial sepsis. In Neutrophils in Infectious Diseases; Tacchini-Cottier, F., van Zandbergen, G., Eds.; Bentham eBooks: Dubais, United Arab Emirates, 2011; pp. 20–36. [Google Scholar]
- Neeli, I.; Dwivedi, N.; Khan, S.; Radic, M. Regulation of extracellular chromatin release from neutrophils. J. Innate Immun. 2009, 1, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Carneiro, C.; Alexandre-Pires, G.; Brito, P.; Pereira, C.; Nunes, T.; Galvão, A.; Leitão, A.; Vilela, C.; Ferreira-Dias, G. Neutrophil Extracellular Traps formation by bacteria causing endometritis in the mare. J Reprod. Immunol. 2014, 106, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Lögters, T.; Margraf, S.; Altrichter, J.; Cinatl, J.; Mitzner, S.; Windolf, J.; Scholz, M. The clinical value of neutrophil extracellular traps. Med. Microbiol. Immunol. 2009, 198, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Chrysanthopoulou, A.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Mikroulis, D.; Konstantinidis, T.; Sivridis, E.; Koffa, M.; Giatromanolaki, A.; Boumpas, D.T.; et al. Neutrophil extracellular traps promote differentiation and function of fibroblasts. J. Pathol. 2014, 233, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Constituents of neutrophil extracellular traps induce fibrosis formation in mare endometrium. Theriogenology 2018, 113, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Foster, D.N. Seminal DNase frees spermatozoa entangled in neutrophil extracellular traps. Biol. Reprod. 2005, 73, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Lovaas, B.J.; Bird, S.L.; Lamb, G.C.; Rendahl, A.K.; Taube, P.C.; Foster, D.N. Species-specific interaction of seminal plasma on sperm-neutrophil binding. Anim. Reprod. Sci. 2009, 114, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Otero, Y.; Zambrano, F.; Catalán, J.; Sánchez, R.; Yeste, M.; Miro, J.; Fernandez-Fuertes, B. Seminal plasma, and not sperm, induces time and concentration-dependent neutrophil extracellular trap release in donkeys. Equine Vet. J. 2022, 54, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, H.N.; Sales, K.J.; Milling Smith, O.P.; Battersby, S.; Boddy, S.C. Prostaglandin receptors are mediators of vascular function in endometrial pathologies. Mol. Cell. Endocrinol. 2006, 252, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.K.; Fisher, A.S.; Scruggs, A.M.; White, E.S.; Hogaboam, C.M.; Richardson, B.C.; Peters-Golden, M. Hypermethylation of PTGER2 confers prostaglandin E2 resistance in fibrotic fibroblasts from humans and mice. Am. J. Pathol. 2010, 177, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Oka, T.; Yamagami, K.; Lee, J.K.; Akazawa, H.; Naito, A.T.; Yasui, T.; Ishizu, T.; Nakaoka, Y.; Sakata, Y.; et al. An EP4 receptor agonist inhibits cardiac fibrosis through activation of PKA signaling in hypertrophied heart. Int. Heart J. 2017, 58, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olman, M.A. Beyond TGF-β: A prostaglandin promotes fibrosis. Nat. Med. 2009, 15, 1360–1361. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Impairment of anti-fibrotic PGE2 pathway might influence neutrophil extracellular traps-induced fibrosis in mare endometrium. Domest. Anim. Endocrinol. 2019, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oga, T.; Matsuoka, T.; Yao, C.; Nonomura, K.; Kitaoka, S.; Sakata, D.; Kita, Y.; Tanizawa, K.; Taguchi, Y.; Chin, K.; et al. Prostaglandin F2α receptor signaling facilitates bleomycin induced pulmonary fibrosis independently of transforming growth factor-β. Nat. Med. 2009, 15, 1426–1430. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.Y.; Ti, Y.; Wang, J.; Wang, Z.H.; Xie, G.L.; Shang, Y.Y.; Tang, M.-X.; Zhang, Y.; Zhang, W.; Zhong, M. The Prostaglandin F2α facilitates collagen synthesis in cardiac fibroblasts via an F-prostanoid receptor/protein kinase C/Rho kinase pathway independent of transforming growth factor β1. Int. J. Biochem. Cell Biol. 2012, 44, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Fernandes, C.; Silva, E.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Enzymes present in neutrophil extracellular traps may stimulate the fibrogenic PGF2α pathway in the mare endometrium. Animals 2021, 11, 2615. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.Z.; Baclawska, A.; Rebordão, M.R.; Ferreira-Dias, G.; Skarzynski, D.J. Prostaglandins effect on matrix metallopeptidases and collagen in mare endometrial fibroblasts. Theriogenology 2020, 153, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Fernandes, C.; Lukasik, K.; Szóstek-Mioduchowska, A.; Baclawska, A.; Rebordão, M.R.; Aguiar-Silva, J.; Pinto-Bravo, P.; Skarzynski, D.J.; Ferreira-Dias, G. Elastase inhibition affects collagen transcription and prostaglandin secretion in mare endometrium during the estrous cycle. Reprod. Domest. Anim. 2018, 53, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Szóstek, A.Z.; Siemienuch, M.J.; Galvão, A.M.; Kukasik, K.; Ferreira-Dias, G.; Skarzynski, D.J. mRNA transcription of Prostaglandin synthases and their products in the equine endometrium in the course of fibrosis. Theriogenology 2012, 78, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Sanders, Y.Y.; Pardo, A.; Selman, M.; Nuovo, G.J.; Tollefsbol, T.O.; Siegal, G.P.; Hagood, J.S. Thy-1 promoter hypermethylation: A novel epigenetic pathogenic mechanism in pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2008, 39, 610–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, B. Impact of aging on DNA methylation. Ageing Res. Rev. 2003, 3, 245–261. [Google Scholar] [CrossRef]


{kind=link}
| Condition | Duration | Treatment | Justification for Treatment |
|---|---|---|---|
| post-breeding endometritis (PBE) | ≤24 h | none | physiological defense mechanism |
| prolonged post-breeding endometritis (PPBE) | >24 h | ecbolics, lavage, immune modulators | presence of intrauterine fluid and neutrophils |
| acute infectious endometritis | days | antimicrobials, ecbolics, lavage, mucolytics | bacteria or fungi cultured |
| chronic infectious endometritis | from one to several weeks | antimicrobials, ecbolics, lavage, mucolytics | bacteria or fungi cultured |
| endometrosis | years | none | degenerative changes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katila, T.; Ferreira-Dias, G. Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares. Animals 2022, 12, 779. https://doi.org/10.3390/ani12060779
Katila T, Ferreira-Dias G. Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares. Animals. 2022; 12(6):779. https://doi.org/10.3390/ani12060779
Chicago/Turabian StyleKatila, Terttu, and Graça Ferreira-Dias. 2022. "Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares" Animals 12, no. 6: 779. https://doi.org/10.3390/ani12060779