
Oxidative Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue and Uterine Discharge Collection
2.2. Cytological Smear Preparation and Cytological Assessment
2.3. Evaluation and Diagnosis of Dairy Endometritis
2.4. Cell Culture and Treatment
2.5. Total RNA Extraction and Quantitative Reverse Transcription PCR
2.6. Detection of Intracellular ROS
2.7. Detection of Mitochondrial Membrane Potential
2.8. Transmission Electron Microscopy
2.9. Protein Extraction and Western Blotting
2.10. Statistical Analysis
3. Results
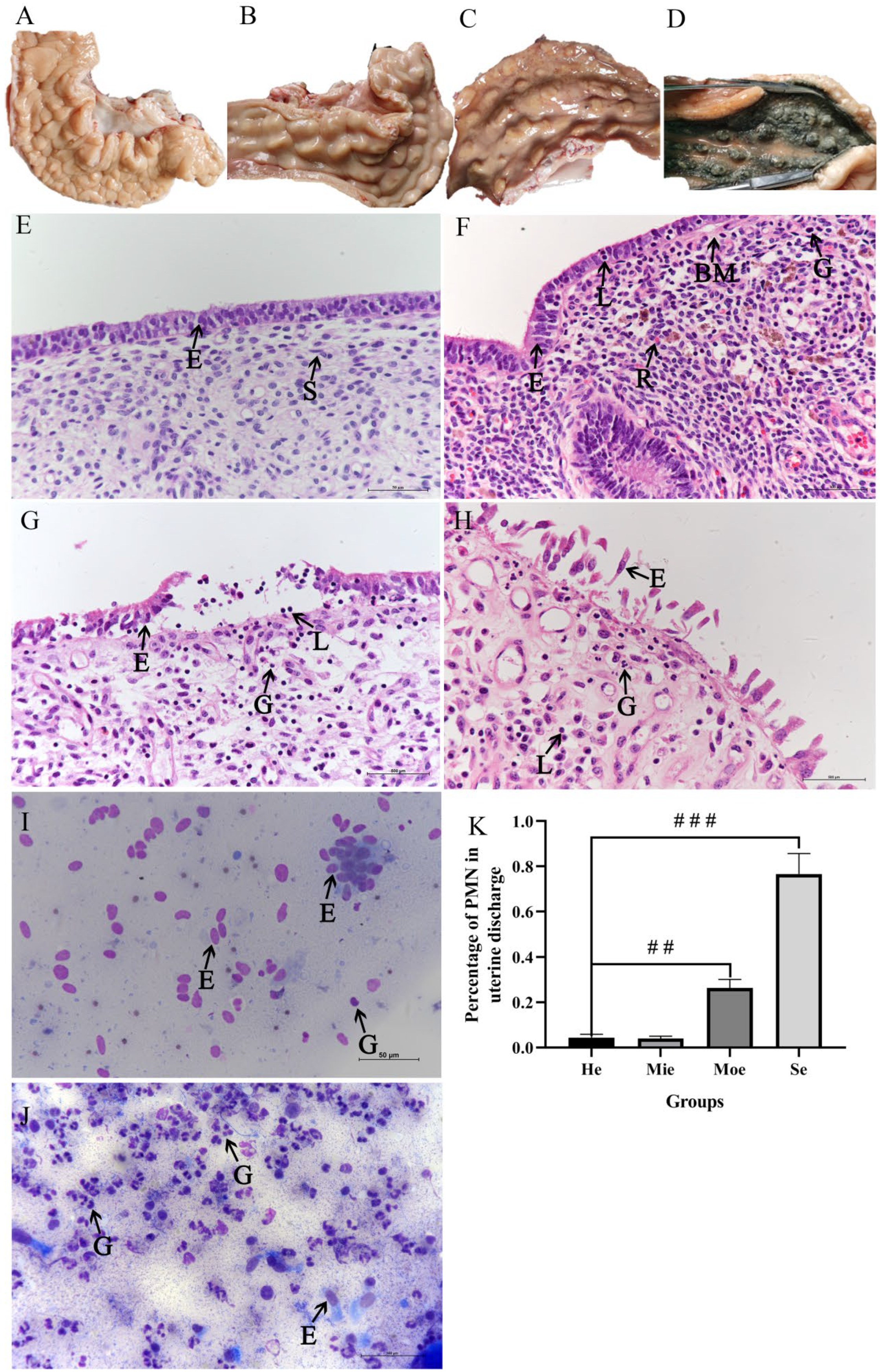
3.1. Examination of Bovine Uteri and Uterine Discharge
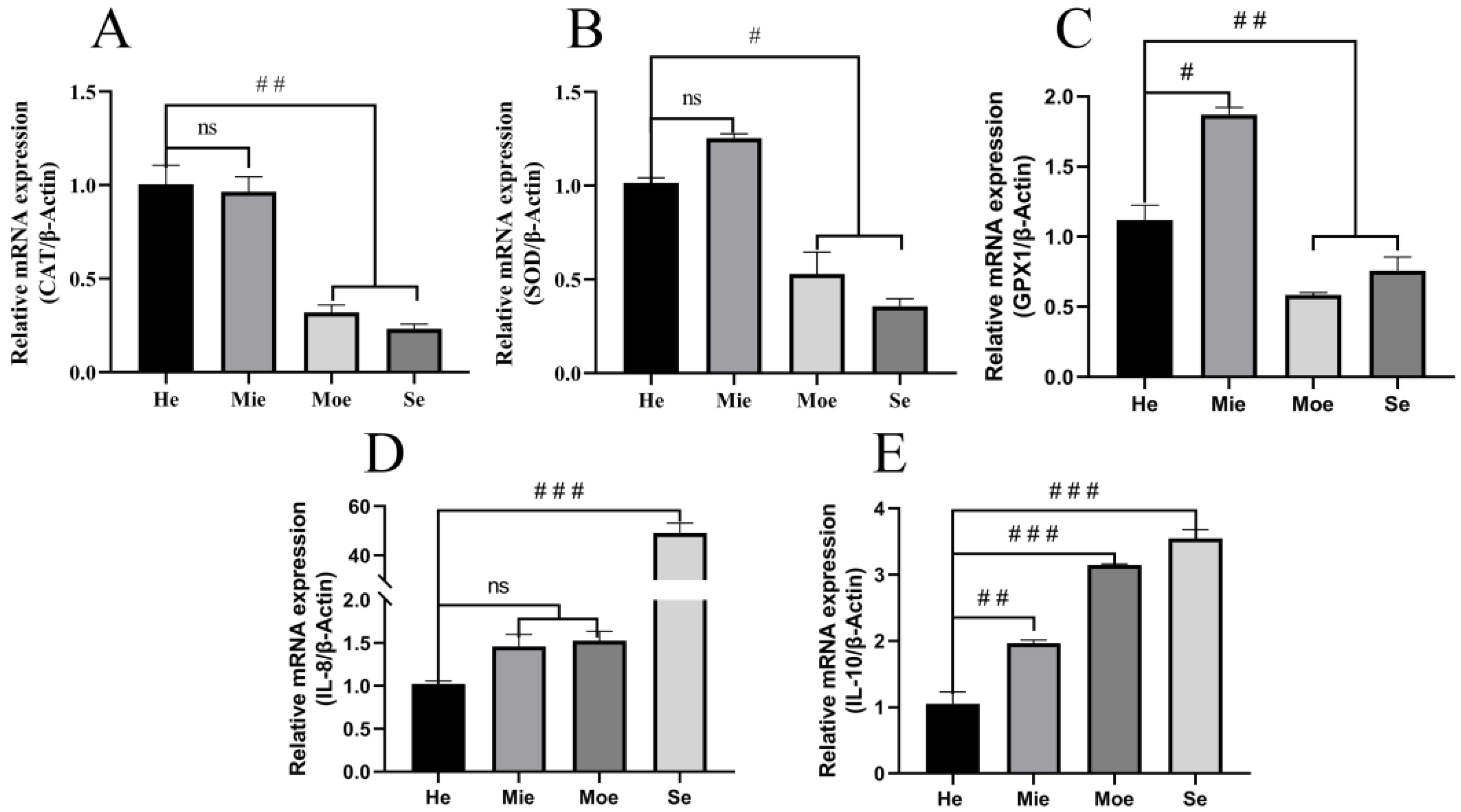
3.2. Glutathione Peroxidase, Superoxide Dismutase, Catalase Show Reduced Expression and IL-8, IL-10 Show Increased Expression in Bovine Uteri with Inflammation
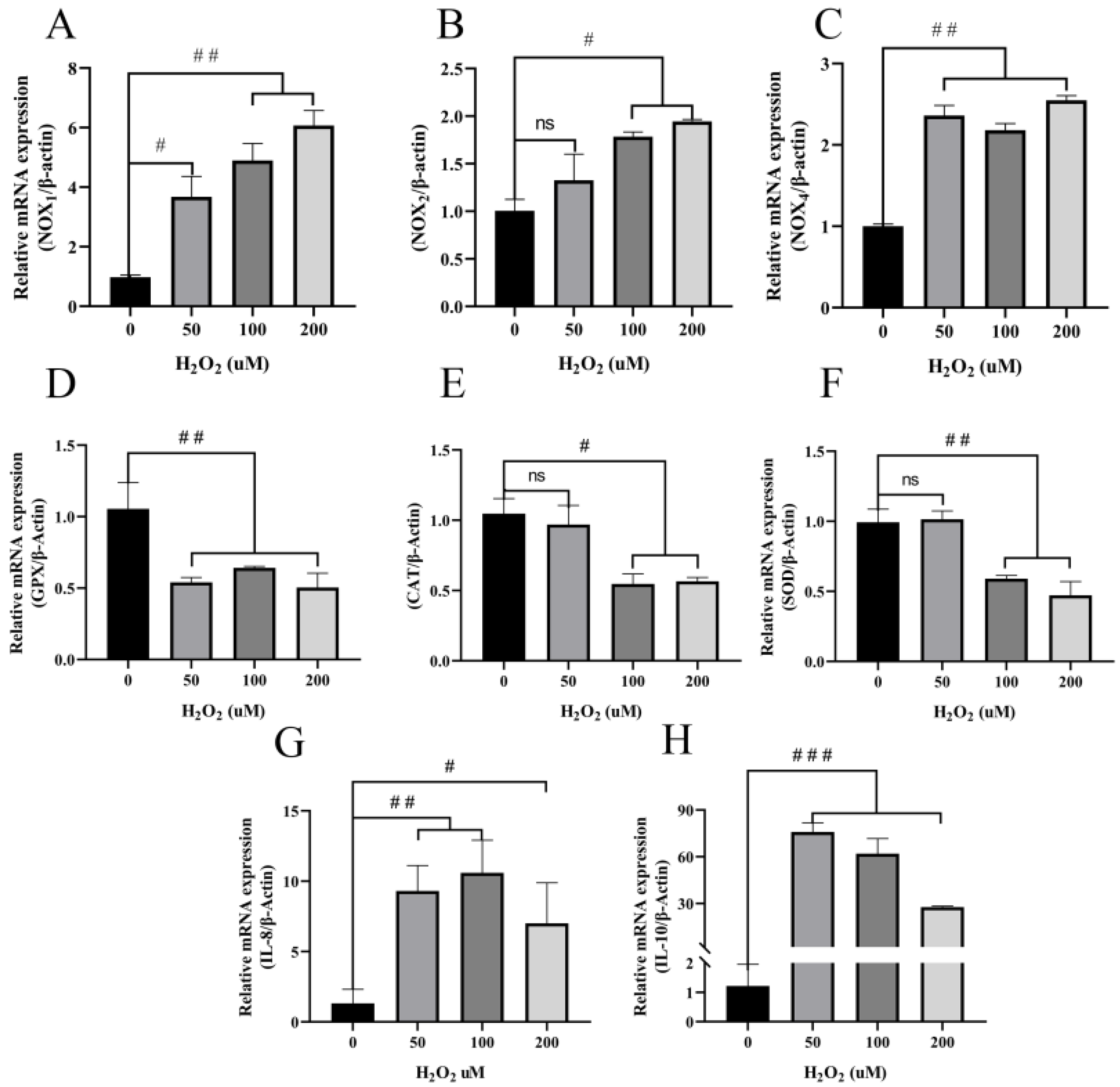
3.3. H2O2 Induces Increased Inflammatory Cytokines and Decreased Antioxidant Enzymes in BEECs
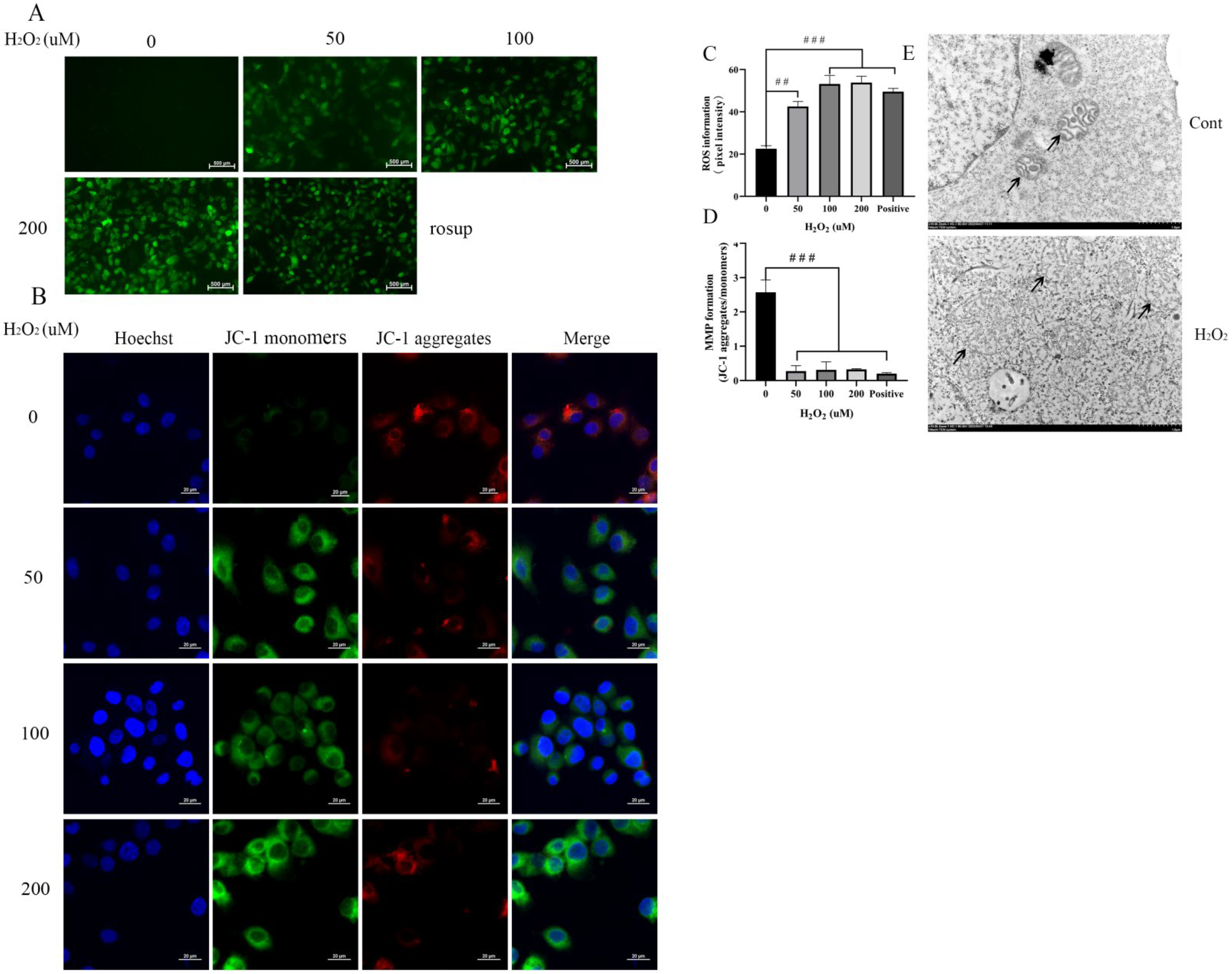
3.4. H2O2 Increases ROS and a Decrease of the MMP to Damage Mitochondria in BEECs
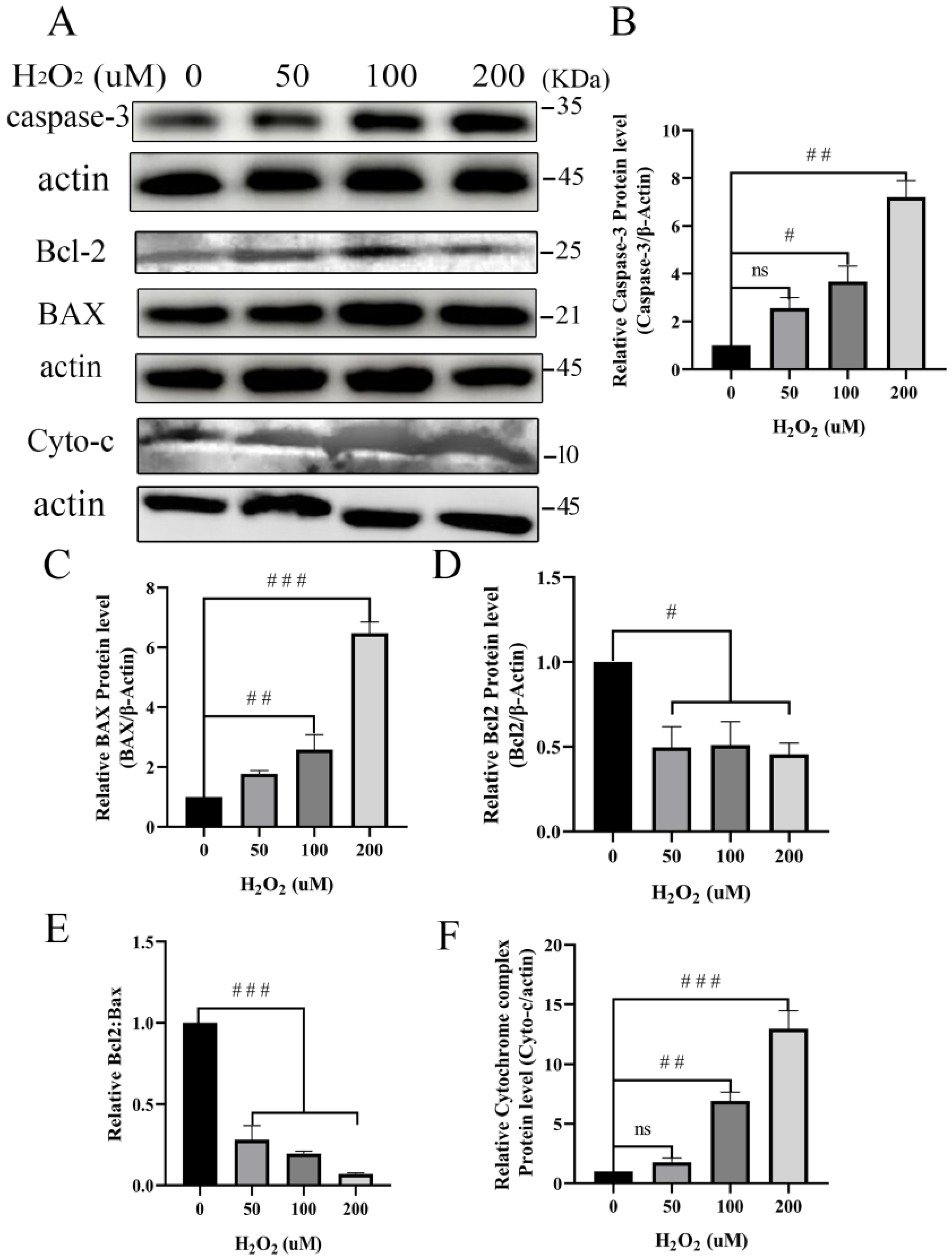
3.5. Pro-Apoptosis of Mitochondria-Dependent Proteins Increases after Treatment with H2O2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilbert, R.O. Management of reproductive disease in dairy cows. Vet. Clin. North Am. Food Anim. Pract. 2016, 32, 387–410. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.G.; Bromfield, J.J. Tolerance and innate immunity shape the development of postpartum uterine disease and the impact of endometritis in dairy cattle. Annu. Rev. Anim. Biosci. 2019, 7, 361–384. [Google Scholar] [CrossRef]
- Esposito, G.; Irons, P.C.; Webb, E.C.; Chapwanya, A. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim. Reprod. Sci. 2014, 144, 60–71. [Google Scholar] [CrossRef]
- Esposito, G.; Raffrenato, E.; Lukamba, S.D.; Adnane, M.; Irons, P.C.; Cormican, P.; Tasara, T.; Chapwanya, A. Characterization of metabolic and inflammatory profiles of transition dairy cows fed an energy-restricted diet. J. Anim. Sci. 2020, 98, skz391. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, D.; Wu, X.; He, B.; Cheng, Z.; Szenci, O.; Song, P.; Shao, D.; Zhang, S.; Yan, Z. Decreasing of S100A4 in bovine endometritis in vivo and in vitro. Theriogenology 2020, 153, 68–73. [Google Scholar] [CrossRef]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef]
- Bretzlaff, K. Rationale for treatment of endometritis in the dairy cow. Vet. Clin. North Am. Food Anim. Pract. 1987, 3, 593–607. [Google Scholar] [CrossRef]
- Sheldon, I.M. Diagnosing postpartum endometritis in dairy cattle. Vet. Rec. 2020, 186, 88–90. [Google Scholar] [CrossRef]
- Mcdougall, S.; Aberdein, D.; Bates, A.; Burke, C.R. Prevalence of endometritis diagnosed by vaginal discharge scoring or uterine cytology in dairy cows and herds. J. Dairy Sci. 2020, 103, 6511–6521. [Google Scholar] [CrossRef]
- Dubuc, J.; Duffield, T.F.; Leslie, K.E.; Walton, J.S.; Leblanc, S.J. Definitions and diagnosis of postpartum endometritis in dairy cows. J. Dairy Sci. 2010, 93, 5225–5233. [Google Scholar] [CrossRef]
- Fine, N.; Tasevski, N.; Mcculloch, C.A.; Tenenbaum, H.C.; Glogauer, M. The neutrophil: Constant defender and first responder. Front. Immunol. 2020, 11, 571085. [Google Scholar] [CrossRef] [PubMed]
- Almyroudis, N.G.; Grimm, M.J.; Davidson, B.A.; Röhm, M.; Urban, C.F.; Segal, B.H. NETosis and NADPH oxidase: At the intersection of host defense, inflammation, and injury. Front. Immunol. 2013, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Huang, J.; Li, Y.; Lv, X.; Zhou, H.; Wang, H.; Xu, Y.; Wang, C.; Wang, J.; Liu, Z. ROS-scavenging hydrogel to promote healing of bacteria infected diabetic wounds. Biomaterials 2020, 258, 120286. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Mae, K.; Nagatani, H.; Horie, M.; Nagamori, E. Effect of hydrogen peroxide concentration on the maintenance and differentiation of cultured skeletal muscle cells. J. Biosci. Bioeng. 2021, 131, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Konno, T.; Melo, E.P.; Chambers, J.E.; Avezov, E. Intracellular sources of ROS/H(2)O(2) in health and neurodegeneration: Spotlight on endoplasmic reticulum. Cells 2021, 10, 233. [Google Scholar] [CrossRef]
- Wanschel, A.; Guizoni, D.M.; Lorza-Gil, E.; Salerno, A.G.; Paiva, A.A.; Dorighello, G.G.; Davel, A.P.; Balkan, W.; Hare, J.M.; Oliveira, H. The presence of cholesteryl ester transfer protein (CETP) in endothelial cells generates vascular oxidative stress and endothelial dysfunction. Biomolecules 2021, 11, 69. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Brenner, C.; Galluzzi, L.; Kepp, O.; Kroemer, G. Decoding cell death signals in liver inflammation. J. Hepatol. 2013, 59, 583–594. [Google Scholar] [CrossRef]
- Valacchi, G.; Virgili, F.; Cervellati, C.; Pecorelli, A. OxInflammation: From subclinical condition to pathological biomarker. Front. Physiol. 2018, 9, 858. [Google Scholar] [CrossRef]
- Duilio, C.; Ambrosio, G.; Kuppusamy, P.; Dipaula, A.; Becker, L.C.; Zweier, J.L. Neutrophils are primary source of O2 radicals during reperfusion after prolonged myocardial ischemia. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2649–H2657. [Google Scholar] [CrossRef] [Green Version]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben, M.M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Gabai, G.; De Luca, E.; Miotto, G.; Zin, G.; Stefani, A.; Da, D.L.; Barberio, A.; Celi, P. Relationship between Protein Oxidation Biomarkers and Uterine Health in Dairy Cows during the Postpartum Period. Antioxidants 2019, 8, 21. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial metabolism of reactive oxygen species. Biochem. Biokhimiia 2005, 70, 200. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Ramirez, F.; Shu-I, T.U.; Chatterji, P.R.; Okazaki, H.; Mckeever, B. The nature of the energy transduction links in mitochondrial oxidative phosphorylation. Acs Sympos. 1981, 171, 205–209. [Google Scholar]
- Correia-álvarez, E.; Keating, J.E.; Glish, G.; Tarran, R.; Sassano, M.F. Reactive oxygen species, mitochondrial membrane potential, and cellular membrane potential are predictors of E-Liquid induced cellular toxicity. Nicotine Tob. Res. 2020, 22, S4–S13. [Google Scholar] [CrossRef]
- Rehfeldt, S.; Laufer, S.; Goettert, M.I. A highly selective in vitro JNK3 inhibitor, FMU200, restores mitochondrial membrane potential and reduces oxidative stress and apoptosis in SH-SY5Y cells. Int. J. Mol. Sci. 2021, 22, 3701. [Google Scholar] [CrossRef]
- Santos, J.H.; Hunakova, L.; Chen, Y.; Bortner, C.; Van Houten, B. Cell sorting experiments link persistent mitochondrial DNA damage with loss of mitochondrial membrane potential and apoptotic cell death. J. Biol. Chem. 2003, 278, 1728–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.K. Mitochondria damage checkpoint in apoptosis and genome stability. FEMS Yeast Res. 2004, 5, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Bedoui, S.; Herold, M.J.; Strasser, A. Emerging connectivity of programmed cell death pathways and its physiological implications. Nat. Rev. Mol. Cell. Biol. 2020, 21, 678–695. [Google Scholar] [CrossRef]
- Xiong, S.; Mu, T.; Wang, G.; Jiang, X. Mitochondria-mediated apoptosis in mammals. Protein Cell 2014, 5, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Hockenbery, D.; Nuñez, G.; Milliman, C.; Schreiber, R.D.; Korsmeyer, S.J. Bcl-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature 1990, 348, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, J.M.; Soane, L. Multiple functions of BCL-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008722. [Google Scholar] [CrossRef]
- Kasimanickam, R.; Duffield, T.F.; Foster, R.A.; Gartley, C.J.; Leslie, K.E.; Walton, J.S.; Johnson, W.H. A comparison of the cytobrush and uterine lavage techniques to evaluate endometrial cytology in clinically normal postpartum dairy cows. Can. Vet. J. 2005, 46, 255–259. [Google Scholar]
- Wagener, K.; Gabler, C.; Drillich, M. A review of the ongoing discussion about definition, diagnosis and pathomechanism of subclinical endometritis in dairy cows. Theriogenology 2017, 94, 21–30. [Google Scholar] [CrossRef]
- de Boer, M.W.; Leblanc, S.J.; Dubuc, J.; Meier, S.; Heuwieser, W.; Arlt, S.; Gilbert, R.O.; Mcdougall, S. Invited review: Systematic review of diagnostic tests for reproductive-tract infection and inflammation in dairy cows. J. Dairy Sci. 2014, 97, 3983–3999. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, D.; Yan, Z. Increasing of matrix metalloproteinase 3 in bovine endometritis. Theriogenology 2021, 175, 83–88. [Google Scholar] [CrossRef]
- Sadeghi, H.; Braun, H.S.; Panti, B.; Opsomer, G.; Bogado, P.O. Validation of a deep learning-based image analysis system to diagnose subclinical endometritis in dairy cows. PLoS ONE 2022, 17, e263409. [Google Scholar] [CrossRef] [PubMed]
- Barański, W.; Podhalicz-Dzięgielewska, M.; Zduńczyk, S.; Janowski, T. The diagnosis and prevalence of subclinical endometritis in cows evaluated by different cytologic thresholds. Theriogenology 2012, 78, 1939–1947. [Google Scholar] [CrossRef] [PubMed]
- Sens, A.; Heuwieser, W. Presence of Escherichia coli, Trueperella pyogenes, α-hemolytic streptococci, and coagulase-negative staphylococci and prevalence of subclinical endometritis. J. Dairy Sci. 2013, 96, 6347–6354. [Google Scholar] [CrossRef] [PubMed]
- Mcdougall, S.; Hussein, H.; Aberdein, D.; Buckle, K.; Roche, J.; Burke, C.; Mitchell, M.; Meier, S. Relationships between cytology, bacteriology and vaginal discharge scores and reproductive performance in dairy cattle. Theriogenology 2011, 76, 229–240. [Google Scholar] [CrossRef]
- Ribeiro, E.S.; Lima, F.S.; Greco, L.F.; Bisinotto, R.S.; Monteiro, A.P.; Favoreto, M.; Ayres, H.; Marsola, R.S.; Martinez, N.; Thatcher, W.W.; et al. Prevalence of periparturient diseases and effects on fertility of seasonally calving grazing dairy cows supplemented with concentrates. J. Dairy Sci. 2013, 96, 5682–5697. [Google Scholar] [CrossRef]
- Hartmann, D.; Rohkohl, J.; Merbach, S.; Heilkenbrinker, T.; Klindworth, H.P.; Schoon, H.A.; Hoedemaker, M. Prevalence of cervicitis in dairy cows and its effect on reproduction. Theriogenology 2016, 85, 247–253. [Google Scholar] [CrossRef]
- Barlund, C.S.; Carruthers, T.D.; Waldner, C.L.; Palmer, C.W. A comparison of diagnostic techniques for postpartum endometritis in dairy cattle. Theriogenology 2008, 69, 714–723. [Google Scholar] [CrossRef]
- Hu, X.; Li, D.; Wang, J.; Guo, J.; Li, Y.; Cao, Y.; Zhang, N.; Fu, Y. Melatonin inhibits endoplasmic reticulum stress-associated TXNIP/NLRP3 inflammasome activation in lipopolysaccharide-induced endometritis in mice. Int. Immunopharmacol. 2018, 64, 101–109. [Google Scholar] [CrossRef]
- Bogado, P.O.; Hostens, M.; Sys, P.; Vercauteren, P.; Opsomer, G. Cytological endometritis at artificial insemination in dairy cows: Prevalence and effect on pregnancy outcome. J. Dairy Sci. 2017, 100, 588–597. [Google Scholar] [CrossRef]
- Zal, F.; Khademi, F.; Taheri, R.; Mostafavi-Pour, Z. Antioxidant ameliorating effects against H(2)O(2)-induced cytotoxicity in primary endometrial cells. Toxicol. Mech. Methods 2018, 28, 122–129. [Google Scholar] [CrossRef]
- Sun, X.; Chang, R.; Tang, Y.; Luo, S.; Jiang, C.; Jia, H.; Xu, Q.; Dong, Z.; Liang, Y.; Loor, J.J.; et al. Transcription factor EB (TFEB)-mediated autophagy protects bovine mammary epithelial cells against H(2)O(2)-induced oxidative damage in vitro. J. Anim. Sci. Biotechnol. 2021, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Ballas, P.; Reinländer, U.; Schlegl, R.; Ehling-Schulz, M.; Drillich, M.; Wagener, K. Characterization of intrauterine cultivable aerobic microbiota at the time of insemination in dairy cows with and without mild endometritis. Theriogenology 2021, 159, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, R.; Duffield, T.F.; Foster, R.A.; Gartley, C.J.; Leslie, K.E.; Walton, J.S.; Johnson, W.H. Endometrial cytology and ultrasonography for the detection of subclinical endometritis in postpartum dairy cows. Theriogenology 2004, 62, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Meira, E.J.; Henriques, L.C.; Sá, L.R.; Gregory, L. Comparison of ultrasonography and histopathology for the diagnosis of endometritis in Holstein-Friesian cows. J. Dairy Sci. 2012, 95, 6969–6973. [Google Scholar] [CrossRef]
- Bogado, P.O.; Hostens, M.; Dini, P.; Vandepitte, J.; Ducatelle, R.; Opsomer, G. Comparison between cytology and histopathology to evaluate subclinical endometritis in dairy cows. Theriogenology 2016, 86, 1550–1556. [Google Scholar] [CrossRef]
- Nielsen, J.M. Endometritis in the mare: A diagnostic study comparing cultures from swab and biopsy. Theriogenology 2005, 64, 510–518. [Google Scholar] [CrossRef]
- Bondurant, R.H. Inflammation in the bovine female reproductive tract. J. Anim. Sci. 1999, 77 (Suppl. S2), 101–110. [Google Scholar] [CrossRef]
- Bonnett, B.N.; Miller, R.B.; Etherington, W.G.; Martin, S.W.; Johnson, W.H. Endometrial biopsy in Holstein-Friesian dairy cows. I. Technique, histological criteria and results. Can. J. Vet. Res. 1991, 55, 155–161. [Google Scholar]
- Bogado, P.O.; Hostens, M.; Dini, P.; Vandepitte, J.; Ducatelle, R.; Opsomer, G. Distribution of inflammation and association between active and chronic alterations within the endometrium of dairy cows. Reprod. Domest. Anim. 2016, 51, 751–757. [Google Scholar] [CrossRef]
- Helfrich, A.L.; Reichenbach, H.D.; Meyerholz, M.M.; Schoon, H.A.; Arnold, G.J.; Fröhlich, T.; Weber, F.; Zerbe, H. Novel sampling procedure to characterize bovine subclinical endometritis by uterine secretions and tissue. Theriogenology 2020, 141, 186–196. [Google Scholar] [CrossRef]
- Rimessi, A.; Previati, M.; Nigro, F.; Wieckowski, M.R.; Pinton, P. Mitochondrial reactive oxygen species and inflammation: Molecular mechanisms, diseases and promising therapies. Int. J. Biochem. Cell Biol. 2016, 81, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Roy, A.; Sil, P.C. Mechanism of the protective action of taurine in toxin and drug induced organ pathophysiology and diabetic complications: A review. Food Funct. 2012, 3, 1251–1264. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Sil, P.C. Taurine ameliorates alloxan-induced diabetic renal injury, oxidative stress-related signaling pathways and apoptosis in rats. Amino Acids 2012, 43, 1509–1523. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.T.; Deng, J.S.; Huang, W.C.; Shieh, P.C.; Chung, M.I.; Huang, G.J. Salvianolic acid c against Acetaminophen-Induced acute liver injury by attenuating inflammation, oxidative stress, and apoptosis through inhibition of the Keap1/Nrf2/HO-1 signaling. Oxid. Med. Cell. Longev. 2019, 2019, 9056845. [Google Scholar] [CrossRef]
- Ransy, C.; Vaz, C.; Lombès, A.; Bouillaud, F. Use of H(2)O(2) to cause oxidative stress, the catalase issue. Int. J. Mol. Sci. 2020, 21, 9149. [Google Scholar] [CrossRef]
- Simpson, D.; Oliver, P.L. ROS generation in microglia: Understanding oxidative stress and inflammation in neurodegenerative disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef]
- Hu, C.; Wu, Z.; Huang, Z.; Hao, X.; Wang, S.; Deng, J.; Yin, Y.; Tan, C. Nox2 impairs VEGF-A-induced angiogenesis in placenta via mitochondrial ROS-STAT3 pathway. Redox Biol. 2021, 45, 102051. [Google Scholar] [CrossRef]
- Lin, B.; Liu, Y.; Zhang, X.; Fan, L.; Shu, Y.; Wang, J. Membrane-Activated fluorescent probe for High-Fidelity imaging of mitochondrial membrane potential. ACS Sens. 2021, 6, 4009–4018. [Google Scholar] [CrossRef]
- Sun, J.; Tian, M.; Lin, W. Monitoring mitochondrial membrane potential by FRET: Development of fluorescent probes enabling ΔΨ(m)-Dependent subcellular migration. Anal. Chim. Acta 2020, 1097, 196–203. [Google Scholar] [CrossRef]
- Early, J.O.; Fagan, L.E.; Curtis, A.M.; Kennedy, O.D. Mitochondria in injury, inflammation and disease of articular skeletal joints. Front. Immunol. 2021, 12, 695257. [Google Scholar] [CrossRef]
- Kelso, G.F.; Porteous, C.M.; Coulter, C.V.; Hughes, G.; Porteous, W.K.; Ledgerwood, E.C.; Smith, R.A.; Murphy, M.P. Selective targeting of a redox-active ubiquinone to mitochondria within cells: Antioxidant and antiapoptotic properties. J. Biol. Chem. 2001, 276, 4588–4596. [Google Scholar] [CrossRef] [PubMed]
- Neitemeier, S.; Jelinek, A.; Laino, V.; Hoffmann, L.; Eisenbach, I.; Eying, R.; Ganjam, G.K.; Dolga, A.M.; Oppermann, S.; Culmsee, C. BID links ferroptosis to mitochondrial cell death pathways. Redox Biol. 2017, 12, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Li, H.; Xiao, Y.; Tang, L.; Zhong, F.; Huang, G.; Xu, J.M.; Xu, A.M.; Dai, R.P.; Zhou, Z.G. Adipocyte fatty Acid-Binding protein promotes Palmitate-Induced mitochondrial dysfunction and apoptosis in macrophages. Front. Immunol. 2018, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, X.; Chi, R.; Qi, J.; Xu, T. Moderate mechanical stress suppresses the IL-1β-induced chondrocyte apoptosis by regulating mitochondrial dynamics. J. Cell. Physiol. 2021, 236, 7504–7515. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, T.; Schinzel, A.; Borner, C. Bcl-w(edding) with mitochondria. Trends Cell Biol. 2004, 14, 8–12. [Google Scholar] [CrossRef]
- Korsmeyer, S.J.; Shutter, J.R.; Veis, D.J.; Merry, D.E.; Oltvai, Z.N. Bcl-2/Bax: A rheostat that regulates an anti-oxidant pathway and cell death. Semin. Cancer Biol. 1993, 4, 327–332. [Google Scholar]
- Yang, N.; Guan, Q.W.; Chen, F.H.; Xia, Q.X.; Yin, X.X.; Zhou, H.H.; Mao, X.Y. Antioxidants targeting mitochondrial oxidative stress: Promising neuroprotectants for epilepsy. Oxid. Med. Cell. Longev. 2020, 2020, 6687185. [Google Scholar] [CrossRef]
- Wang, X.; Ni, H.; Xu, W.; Wu, B.; Xie, T.; Zhang, C.; Cheng, J.; Li, Z.; Tao, L.; Zhang, Y. Difenoconazole induces oxidative DNA damage and mitochondria mediated apoptosis in SH-SY5Y cells. Chemosphere 2021, 283, 131160. [Google Scholar] [CrossRef]
- Yang, B.; Johnson, T.S.; Thomas, G.L.; Watson, P.F.; Wagner, B.; Furness, P.N.; El, N.A. A shift in the Bax/Bcl-2 balance may activate caspase-3 and modulate apoptosis in experimental glomerulonephritis. Kidney Int. 2002, 62, 1301–1313. [Google Scholar] [CrossRef]
- Chimenti, M.S.; Sunzini, F.; Fiorucci, L.; Botti, E.; Fonti, G.L.; Conigliaro, P.; Triggianese, P.; Costa, L.; Caso, F.; Giunta, A.; et al. Potential role of cytochrome c and tryptase in psoriasis and psoriatic arthritis pathogenesis: Focus on resistance to apoptosis and oxidative stress. Front. Immunol. 2018, 9, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histopathologic Feature of Epithelium | Mononuclear in Lamina Propria (%) | Cytological PMN% in Mucus | Description |
|---|---|---|---|
| Columnar | Mononuclear < 3% | PMN < 2% | Normal |
| Cuboidal | 3–4% < Mononuclear < 5% | 5% < PMN <18% | Mild |
| Flattened | 5% < Mononuclear < 10% | 18% < PMN < 25% | Moderate |
| Necrosis and loss | 10% < Mononuclear | 25% < PMN | Severe |
| Gene Name | ID | Sequence | Size (bp) |
|---|---|---|---|
| CAT | NM_001035386.2 | F: AGAGGAAACGCCTGTGTGAG R: ATGCGGGAGCCATATTCAGG | 115 |
| SOD | NM_174615.2 | F: CTCTACTTGGTTGGGGCGTC R: TCGAAGTGGATGGTGCCTTG | 122 |
| GPx | NM_174076.3 | F: AACGTAGCATCGCTCTGAGG R: GATGCCCAAACTGGTTGCAG | 121 |
| NOX1 | NM_001191340.1 | F: TGTCTTTCCTGAGAGGCACC R: TTTGTGGAAGGCGAGGTTGT | 80 |
| NOX2 | NM_174035.4 | F: CAAGATGGAGGTGGGCCAAT R: GAGGTCAGGGTGAAAGGGTG | 81 |
| NOX4 | NM_001304775.1 | F: TCTGGACCTTTGTGCCT R: GACGGATGACTTGTGACTG | 95 |
| IL-8 | NM_173925.2 | F: CATTCCACACCTTTCCACCC R: AGGCAGACCTCGTTTCCATT | 116 |
| IL-10 | NM_174088.1 | F: CACAGGCTGAGAACCACG R: AGGGCAGAAAGCGATGA | 108 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, P.; Liu, C.; Sun, M.; Liu, J.; Lin, P.; Wang, A.; Jin, Y. Oxidative Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways. Animals 2022, 12, 2444. https://doi.org/10.3390/ani12182444
Song P, Liu C, Sun M, Liu J, Lin P, Wang A, Jin Y. Oxidative Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways. Animals. 2022; 12(18):2444. https://doi.org/10.3390/ani12182444
Chicago/Turabian StyleSong, Pengjie, Chen Liu, Mingkun Sun, Jianguo Liu, Pengfei Lin, Aihua Wang, and Yaping Jin. 2022. "Oxidative Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways" Animals 12, no. 18: 2444. https://doi.org/10.3390/ani12182444