
Dynamics and Diversity of Intrauterine Anaerobic Microbiota in Dairy Cows with Clinical and Subclinical Endometritis
 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Farm and Study Animals
2.2. Gynecological Examination and Diagnosis of Clinical and Subclinical Endometritis
2.3. Bacteriological Sampling
2.4. Statistical Analysis
3. Results
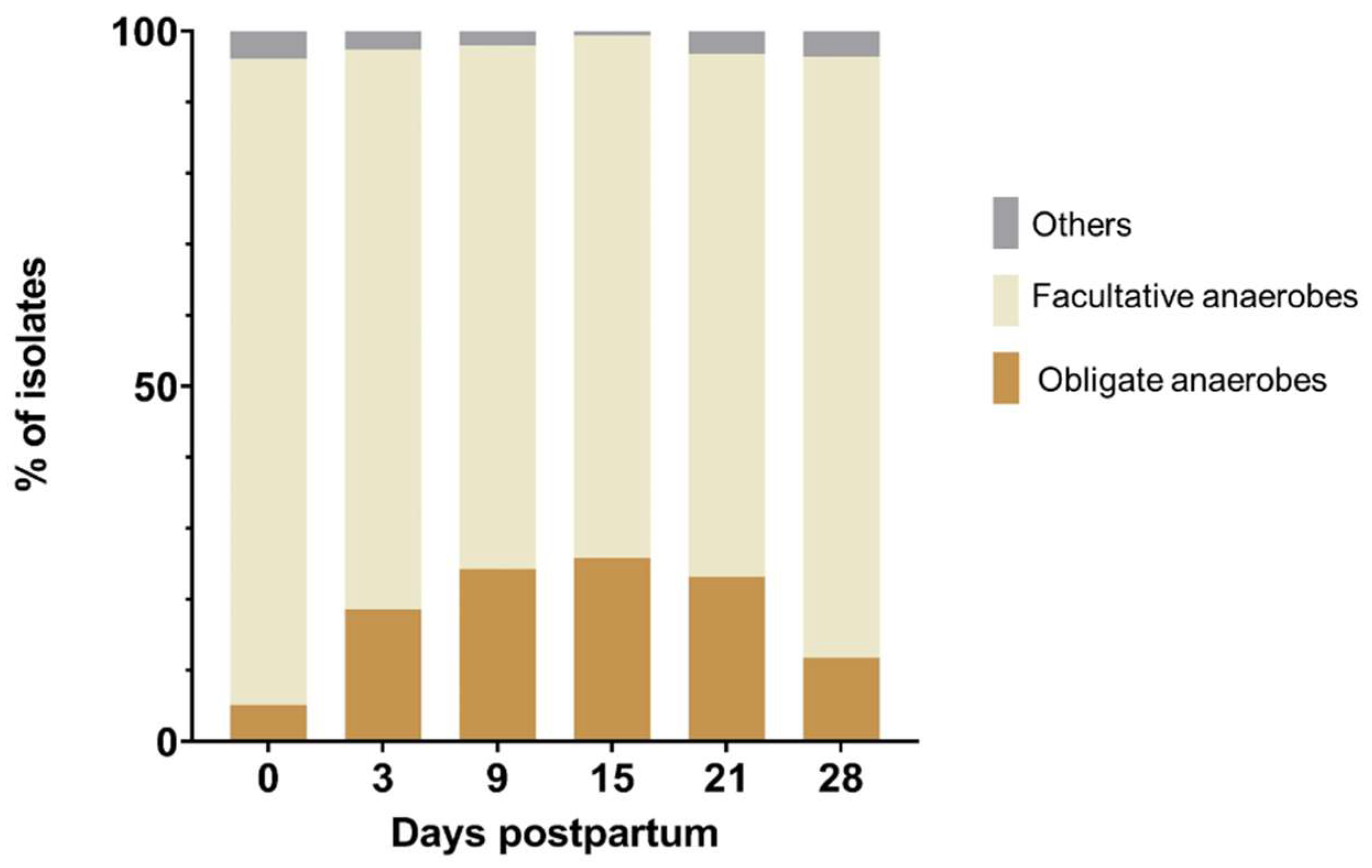
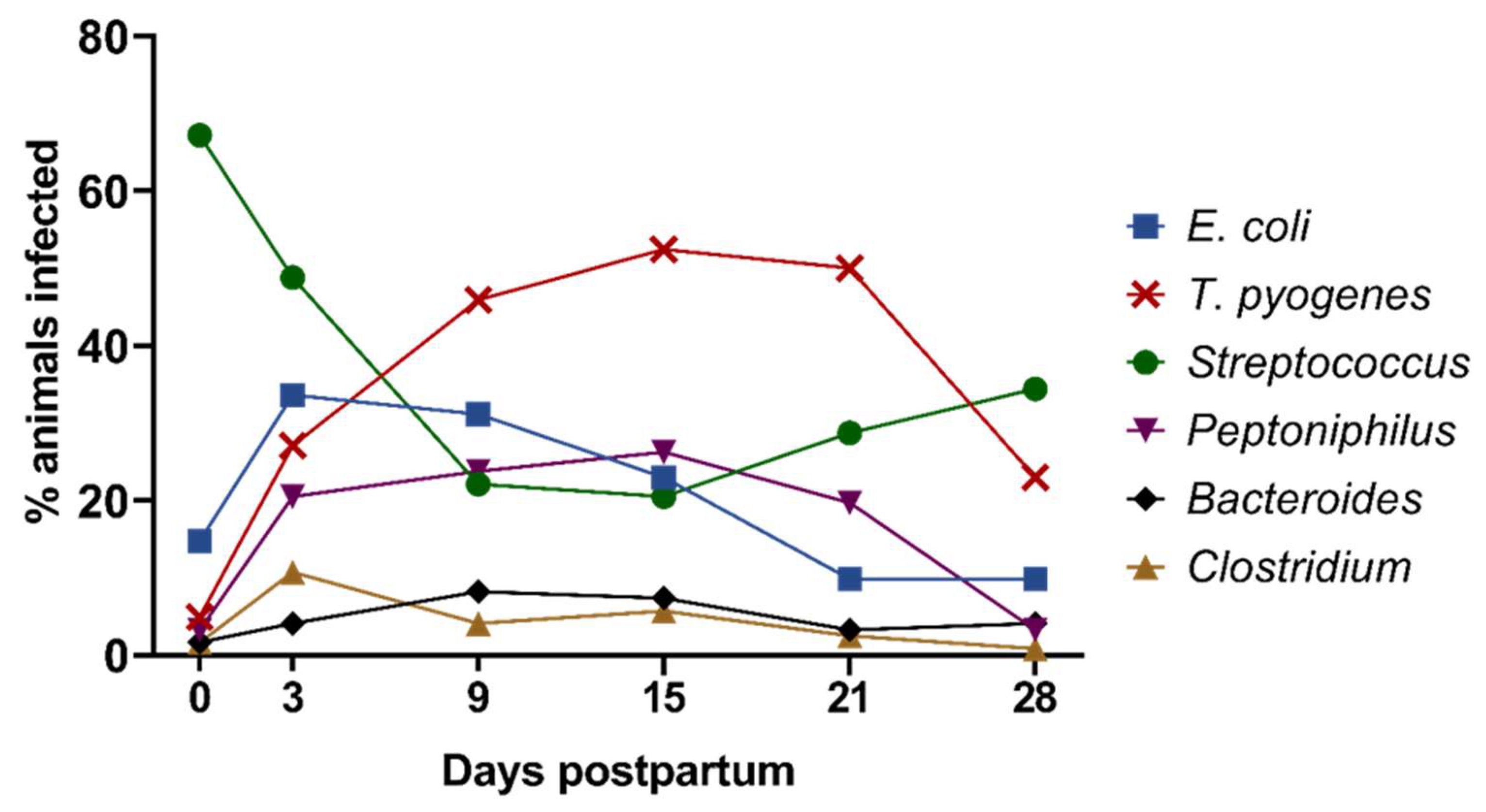
3.1. Composition and Dynamics of Cultivable Anaerobic Intrauterine Microbiota
3.2. Co-Occurrence of Different Bacterial Species and Genera
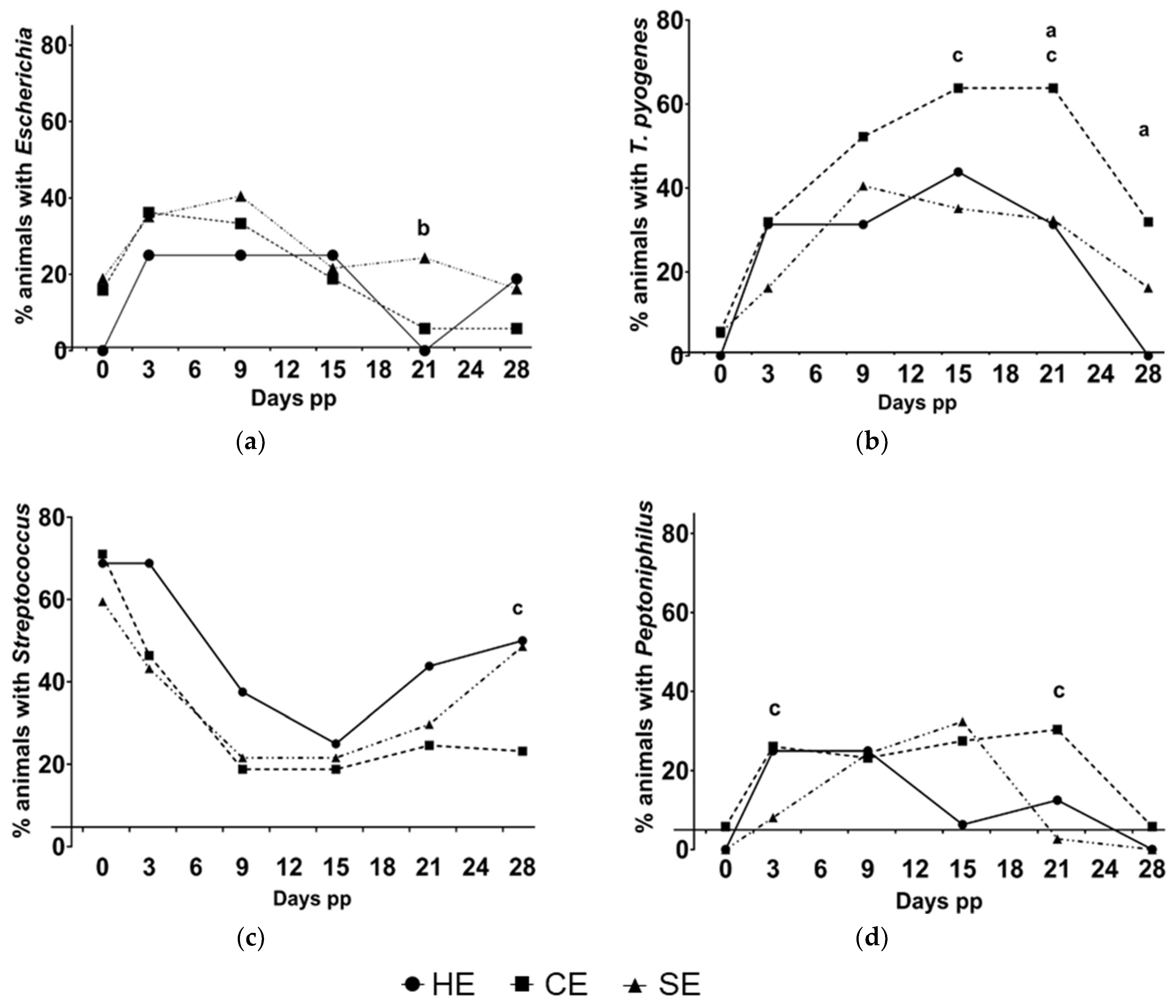
3.3. Prevalence of Bacteriological Findings in HE, CE, and SE Cows
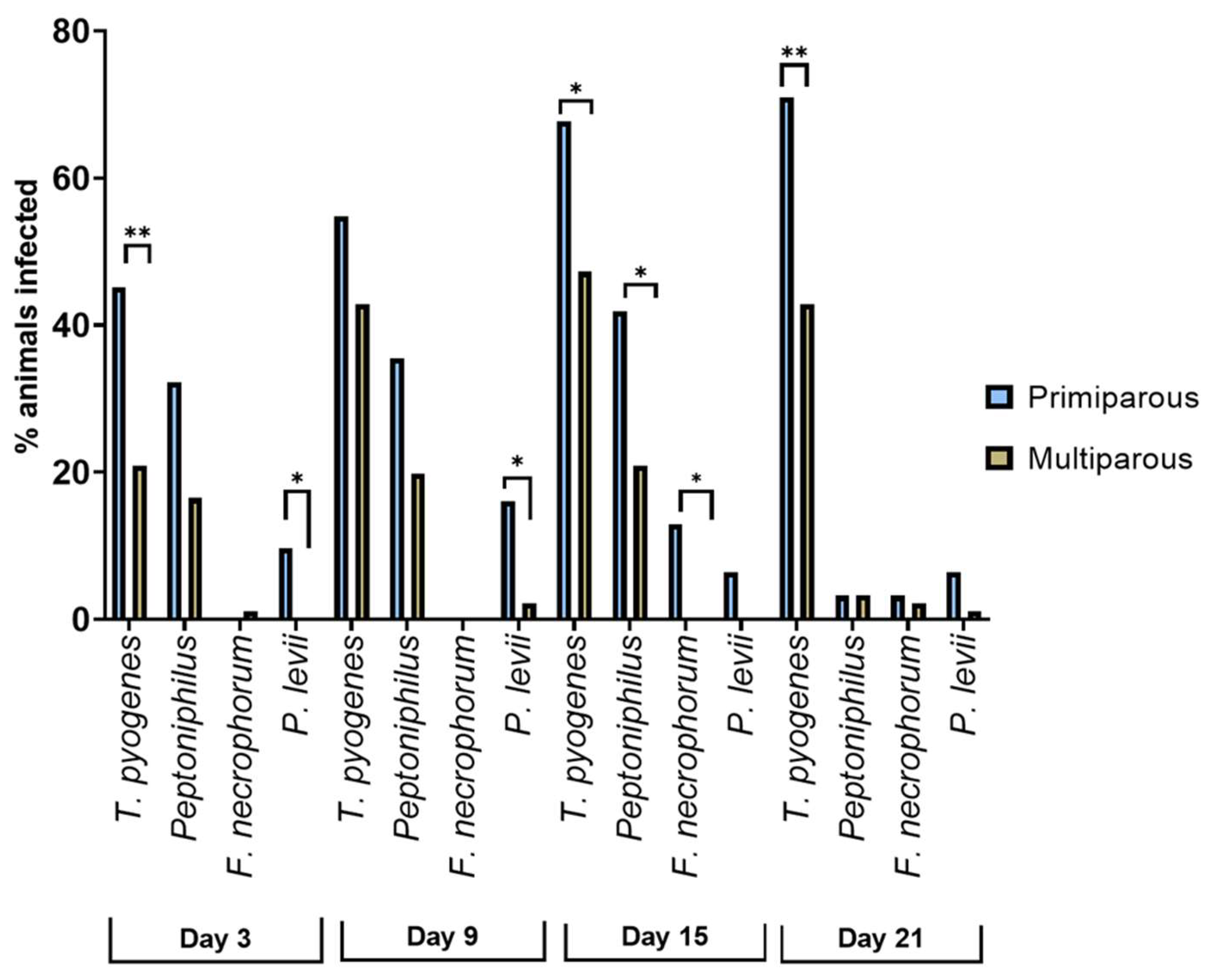
3.4. Prevalence of Bacteriological Findings in Primi- and Multiparous Cows
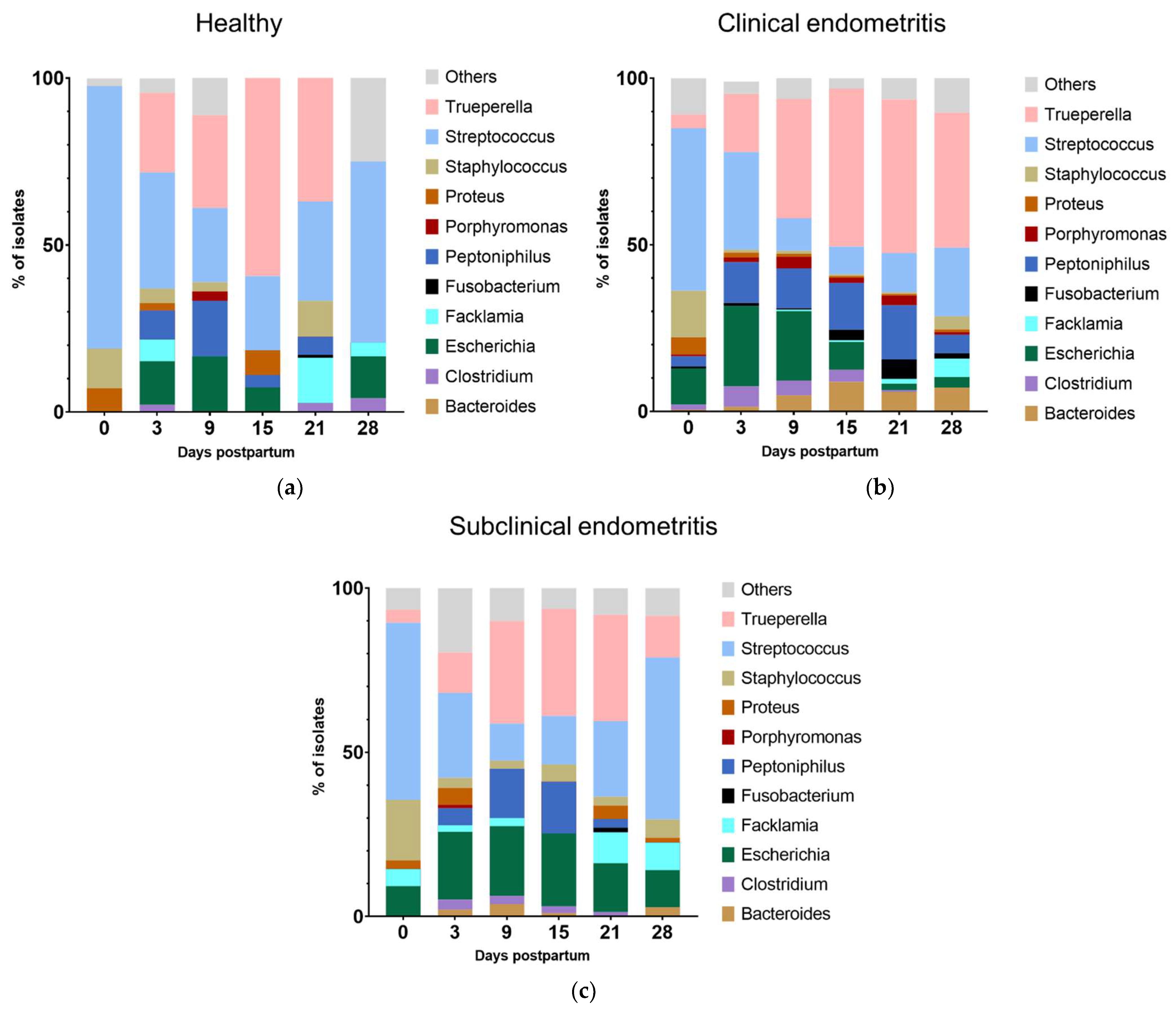
3.5. Dynamics of the Microbiota in Relation to the Uterine Hhealth Status
3.6. Relative Risk for Different Vaginal Discharge Scores and Odds Ratio for the Occurrence Subclinical Endometritis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carneiro, L.C.; Cronin, J.G.; Sheldon, I.M. Mechanisms linking bacterial infections of the bovine endometrium to disease and infertility. Reprod. Biol. 2016, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.G.; Ericsson, A.C.; Poock, S.E.; Melendez, P.; Lucy, M.C. Hot topic: 16S rRNA gene sequencing reveals the microbiome of the virgin and pregnant bovine uterus. J. Dairy Sci. 2017, 100, 4953–4960. [Google Scholar] [CrossRef] [Green Version]
- Karstrup, C.C.; Klitgaard, K.; Jensen, T.K.; Agerholm, J.S.; Pedersen, H.G. Presence of bacteria in the endometrium and placentomes of pregnant cows. Theriogenology 2017, 99, 41–47. [Google Scholar] [CrossRef]
- Ballas, P.; Reinländer, U.; Schlegl, R.; Ehling-Schulz, M.; Drillich, M.; Wagener, K. Characterization of intrauterine cultivable aerobic microbiota at the time of insemination in dairy cows with and without mild endometritis. Theriogenology 2021, 159, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Paiano, R.B.; Moreno, L.Z.; Gomes, V.T.M.; Parra, B.M.; Barbosa, M.R.; Sato, M.I.Z.; Bonilla, J.; Pugliesi, G.; Baruselli, P.S.; Moreno, A.M. Assessment of the main pathogens associated with clinical and subclinical endometritis in cows by culture and MALDI-TOF mass spectrometry identification. J. Dairy Sci. 2022, 105, 3367–3376. [Google Scholar] [CrossRef] [PubMed]
- Sicsic, R.; Goshen, T.; Dutta, R.; Kedem-Vaanunu, N.; Kaplan-Shabtai, V.; Pasternak, Z.; Gottlieb, Y.; Shpigel, N.Y.; Raz, T. Microbial communities and inflammatory response in the endometrium differ between normal and metritic dairy cows at 5–10 days post-partum. Vet. Res. 2018, 49, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicalho, M.L.S.; Machado, V.S.; Higgins, C.H.; Lima, F.S.; Bicalho, R.C. Genetic and functional analysis of the bovine uterine microbiota. Part I: Metritis versus healthy cows. J. Dairy Sci. 2017, 100, 3850–3862. [Google Scholar] [CrossRef]
- Bicalho, M.L.S.; Lima, S.; Higgins, C.H.; Machado, V.S.; Lima, F.S.; Bicalho, R.C. Genetic and functional analysis of the bovine uterine microbiota. Part II: Purulent vaginal discharge versus healthy cows. J. Dairy Sci. 2017, 100, 3863–3874. [Google Scholar] [CrossRef] [Green Version]
- Madoz, L.V.; Giuliodori, M.J.; Migliorisi, A.L.; Jaureguiberry, M.; de la Sota, R.L. Endometrial cytology, biopsy, and bacteriology for the diagnosis of subclinical endometritis in grazing dairy cows. J. Dairy Sci. 2014, 97, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Prunner, I.; Wagener, K.; Pothmann, H.; Ehling-Schulz, M.; Drillich, M. Risk factors for uterine diseases on small- and medium-sized dairy farms determined by clinical, bacteriological, and cytological examinations. Theriogenology 2014, 82, 857–865. [Google Scholar] [CrossRef]
- Baranski, W.; Podhalicz-Dziegielewska, M.; Zdunczyk, S.; Janowski, T. The diagnosis and prevalence of subclinical endometritis in cows evaluated by different cytologic thresholds. Theriogenology 2012, 78, 1939–1947. [Google Scholar] [CrossRef] [PubMed]
- Prunner, I.; Pothmann, H.; Wagener, K.; Giuliodori, M.; Huber, J.; Ehling-Schulz, M.; Drillich, M. Dynamics of bacteriologic and cytologic changes in the uterus of postpartum dairy cows. Theriogenology 2014, 82, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Wagener, K.; Prunner, I.; Pothmann, H.; Drillich, M.; Ehling-Schulz, M. Diversity and health status specific fluctuations of intrauterine microbial communities in postpartum dairy cows. Vet. Microbiol. 2015, 175, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Miranda-CasoLuengo, R.; Lu, J.; Williams, E.J.; Miranda-CasoLuengo, A.A.; Carrington, S.D.; Evans, A.C.O.; Meijer, W.G. Delayed differentiation of vaginal and uterine microbiomes in dairy cows developing postpartum endometritis. PLoS ONE 2019, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sens, A.; Heuwieser, W. Presence of Escherichia coli, Trueperella pyogenes, alpha-hemolytic streptococci, and coagulase-negative staphylococci and prevalence of subclinical endometritis. J. Dairy Sci. 2013, 96, 6347–6354. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Suthar, V.; Plöntzke, J.; Heuwieser, W. Relationship between bacteriological findings in the second and fourth weeks postpartum and uterine infection in dairy cows considering bacteriological results. J. Dairy Sci. 2012, 95, 7105–7114. [Google Scholar] [CrossRef]
- Santos, T.M.; Bicalho, R.C. Diversity and succession of bacterial communities in the uterine fluid of postpartum metritic, endometritic and healthy dairy cows. PLoS ONE 2012, 7, e53048. [Google Scholar] [CrossRef] [Green Version]
- Pascottini, O.B.; Van Schyndel, S.J.; Spricigo, J.F.W.; Rousseau, J.; Weese, J.S.; LeBlanc, S.J. Dynamics of uterine microbiota in postpartum dairy cows with clinical or subclinical endometritis. Sci. Rep. 2020, 10, 12353. [Google Scholar] [CrossRef]
- Ballas, P.; Gabler, C.; Wagener, K.; Drillich, M.; Ehling-Schulz, M. Streptococcus uberis strains originating from bovine uteri provoke upregulation of pro-inflammatory factors mRNA expression of endometrial epithelial cells in vitro. Vet. Microbiol. 2020, 245, 108710. [Google Scholar] [CrossRef]
- Danesh Mesgaran, S.; Gärtner, M.A.; Wagener, K.; Drillich, M.; Ehling-Schulz, M.; Einspanier, R.; Gabler, C. Different inflammatory responses of bovine oviductal epithelial cells in vitro to bacterial species with distinct pathogenicity characteristics and passage number. Theriogenology 2018, 106, 237–246. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Westermann, S.; Drillich, M.; Kaufmann, T.B.; Madoz, L.V.; Heuwieser, W. A clinical approach to determine false positive findings of clinical endometritis by vaginoscopy by the use of uterine bacteriology and cytology in dairy cows. Theriogenology 2010, 74, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, R.; Duffield, T.F.; Foster, R.A.; Gartley, C.J.; Leslie, K.E.; Walton, J.S.; Johnson, W.H. A comparison of the cytobrush and uterine lavage techniques to evaluate endometrial cytology in clinically normal postpartum dairy cows. Can. Vet. J. 2005, 46, 255–259. [Google Scholar] [PubMed]
- Ballas, P.; Rückert, C.; Wagener, K.; Drillich, M.; Kämpfer, P.; Busse, H.J.; Ehling-Schulz, M. Corynebacterium endometrii sp. Nov., isolated from the uterus of a cow with endometritis. Int. J. Syst. Evol. Microbiol. 2020, 70, 146–152. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Machado, V.S.; Oikonomou, G.; Bicalho, M.L.S.; Knauer, W.A.; Gilbert, R.; Bicalho, R.C. Investigation of postpartum dairy cows’ uterine microbial diversity using metagenomic pyrosequencing of the 16S rRNA gene. Vet. Microbiol. 2012, 159, 460–469. [Google Scholar] [CrossRef]
- Wang, M.L.; Liu, M.C.; Xu, J.; An, L.G.; Wang, J.F.; Zhu, Y.H. Uterine microbiota of dairy cows with clinical and subclinical endometritis. Front. Microbiol. 2018, 9, 2691. [Google Scholar] [CrossRef]
- Wagener, K.; Grunert, T.; Prunner, I.; Ehling-Schulz, M.; Drillich, M. Dynamics of uterine infections with Escherichia coli, Streptococcus uberis and Trueperella pyogenes in post-partum dairy cows and their association with clinical endometritis. Vet. J. 2014, 202, 527–532. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Molinari, P.C.C.; Ormsby, T.J.R.; Bromfield, J.J. Preventing postpartum uterine disease in dairy cattle depends on avoiding, tolerating and resisting pathogenic bacteria. Theriogenology 2020, 150, 158–165. [Google Scholar] [CrossRef]
- Ibrahim, M.; Peter, S.; Wagener, K.; Drillich, M.; Ehling-Schulz, M.; Einspanier, R.; Gabler, C. Bovine endometrial epithelial cells scale their pro-inflammatory response in vitro to pathogenic Trueperella pyogenes isolated from the bovine uterus in a strain-specific manner. Front. Cell. Infect. Microbiol. 2017, 7, 264. [Google Scholar] [CrossRef]
- Piersanti, R.L.; Zimpel, R.; Molinari, P.C.C.; Dickson, M.J.; Ma, Z.; Jeong, K.C.; Santos, J.E.P.; Sheldon, I.M.; Bromfield, J.J. A model of clinical endometritis in Holstein heifers using pathogenic Escherichia coli and Trueperella pyogenes. J. Dairy Sci. 2019, 102, 2686–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicalho, M.L.S.; Machado, V.S.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. Association between virulence factors of Escherichia coli, Fusobacterium necrophorum, and Arcanobacterium pyogenes and uterine diseases of dairy cows. Vet. Microbiol. 2012, 157, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, L.R.; Karstrup, C.C.; Pedersen, H.G.; Angen, O.; Agerholm, J.S.; Rasmussen, E.L.; Jensen, T.K.; Klitgaard, K. An investigation of the microbiota in uterine flush samples and endometrial biopsies from dairy cows during the first 7 weeks postpartum. Theriogenology 2016, 86, 642–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledgard, A.M.; Smolenski, G.A.; Henderson, H.; Lee, R.S.F. Influence of pathogenic bacteria species present in the postpartum bovine uterus on proteome profiles. Reprod. Fertil. Dev. 2015, 27, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, G.; Barley, J.; Howie, F.; Falsen, E.; Moore, E.; Twomey, D.F.; Wragg, P.; Whatmore, A.M.; Stubberfield, E. Streptococcus pluranimalium in bovine reproductive disease. Vet. Rec. 2008, 163, 638. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, Y.; Hang, S.; Zhu, W. Microbial diversity in uterus of healthy and metritic postpartum Holstein dairy cows. Folia Microbiol. 2013, 58, 593–600. [Google Scholar] [CrossRef]
- Jeon, S.J.; Galvão, K.N. An advanced understanding of uterine microbial ecology associated with metritis in dairy cows. Genom. Inform. 2018, 16, e21. [Google Scholar] [CrossRef] [Green Version]
- Gärtner, M.A.; Bondzio, A.; Braun, N.; Jung, M.; Einspanier, R.; Gabler, C. Detection and characterisation of Lactobacillus spp. in the bovine uterus and their influence on bovine endometrial epithelial cells in vitro. PLoS ONE 2015, 10, e0119793. [Google Scholar] [CrossRef] [Green Version]
- Peter, S.; Gärtner, M.A.; Michel, G.; Ibrahim, M.; Klopfleisch, R.; Lübke-Becker, A.; Jung, M.; Einspanier, R.; Gabler, C. Influence of intrauterine administration of Lactobacillus buchneri on reproductive performance and pro-inflammatory endometrial mRNA expression of cows with subclinical endometritis. Sci. Rep. 2018, 8, 5473. [Google Scholar] [CrossRef] [Green Version]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Finding | VDS 1 | VDS 2 | VDS 3 | |||
|---|---|---|---|---|---|---|
| RRR | 95% CI | RRR | 95% CI | RRR | 95% CI | |
| Streptococcus, day 15 | 0.33 | 0.67–1.66 | 0.15 | 0.17–1.29 | 4.82 * | 1.23–18.87 |
| Peptoniphilus, day 21 | 2.25 | 0.44–11.67 | 3.14 | 0.66–14.95 | 12.44 ** | 2.34–66.10 |
| T. pyogenes, day 21 | 1.64 | 0.58–4.65 | 4.13 * | 1.37–12.44 | 5.08 ** | 1.34–19.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballas, P.; Pothmann, H.; Pothmann, I.; Drillich, M.; Ehling-Schulz, M.; Wagener, K. Dynamics and Diversity of Intrauterine Anaerobic Microbiota in Dairy Cows with Clinical and Subclinical Endometritis. Animals 2023, 13, 82. https://doi.org/10.3390/ani13010082
Ballas P, Pothmann H, Pothmann I, Drillich M, Ehling-Schulz M, Wagener K. Dynamics and Diversity of Intrauterine Anaerobic Microbiota in Dairy Cows with Clinical and Subclinical Endometritis. Animals. 2023; 13(1):82. https://doi.org/10.3390/ani13010082
Chicago/Turabian StyleBallas, Panagiotis, Harald Pothmann, Isabella Pothmann, Marc Drillich, Monika Ehling-Schulz, and Karen Wagener. 2023. "Dynamics and Diversity of Intrauterine Anaerobic Microbiota in Dairy Cows with Clinical and Subclinical Endometritis" Animals 13, no. 1: 82. https://doi.org/10.3390/ani13010082