
Morphological Characteristics and Transcriptome Landscapes of Chicken Follicles during Selective Development

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Histological Observation of Follicles
2.3. ELISA
2.4. RNA Extraction, Library Construction, and Sequencing
2.5. Bioinformatics Analyses
2.6. Quantitative Reverse Transcription PCR (qRT-PCR)
2.7. Statistical Analyses
3. Results
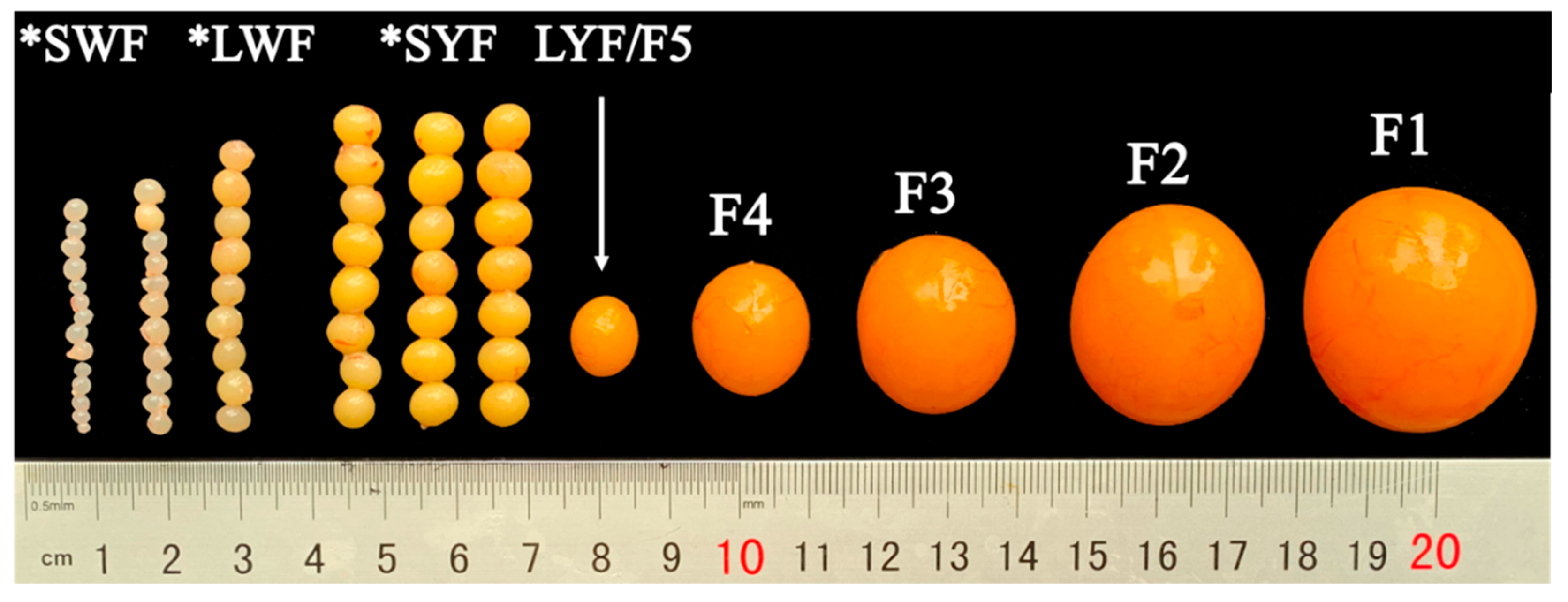
3.1. Number and Size of Follicles at Development Stage
3.2. Histological Observation and Follicle Progesterone Concentrations
3.3. Experimental Design and RNA-Seq Overview
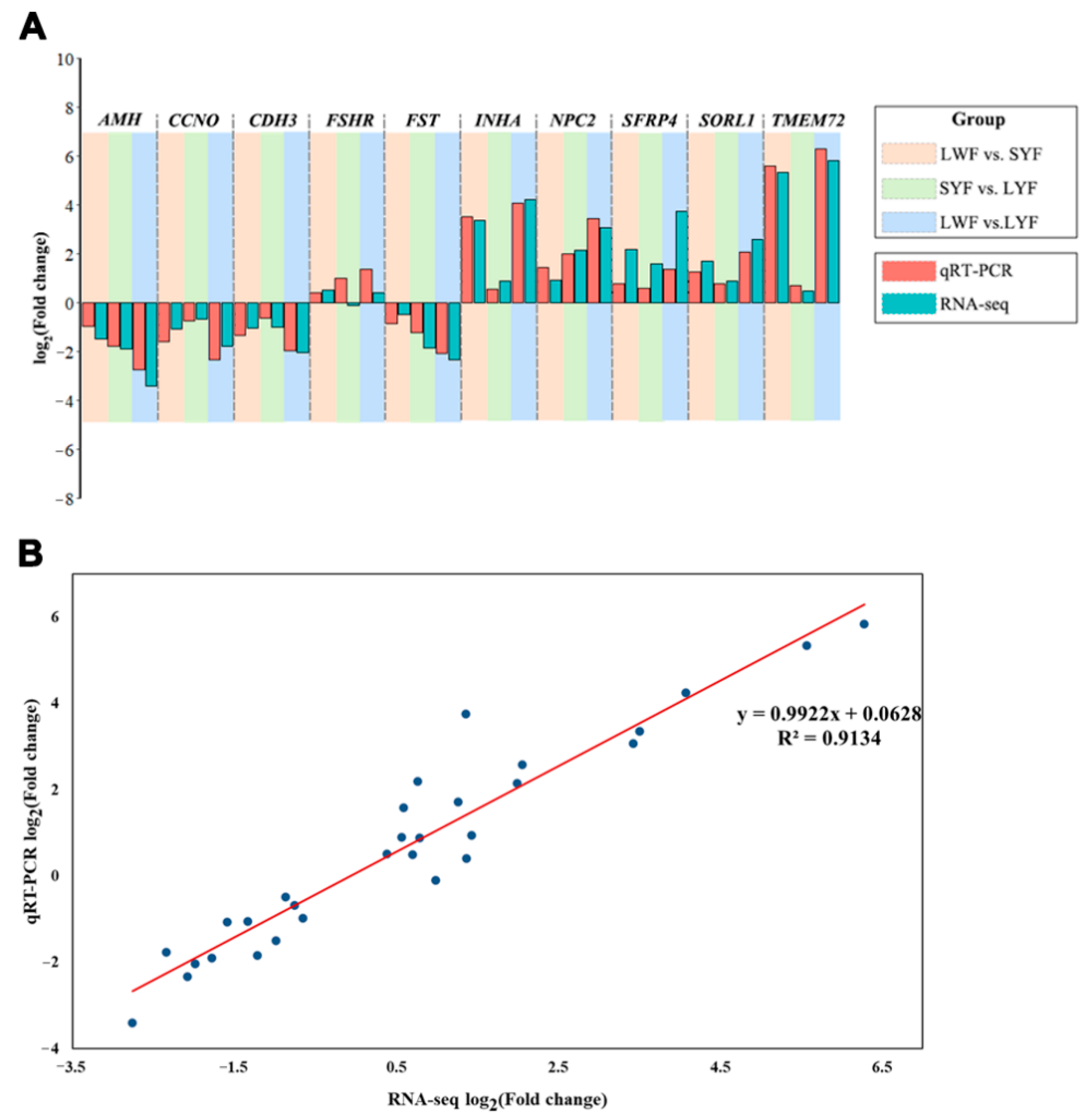
3.4. Validation of RNA-Seq Data by qRT-PCR
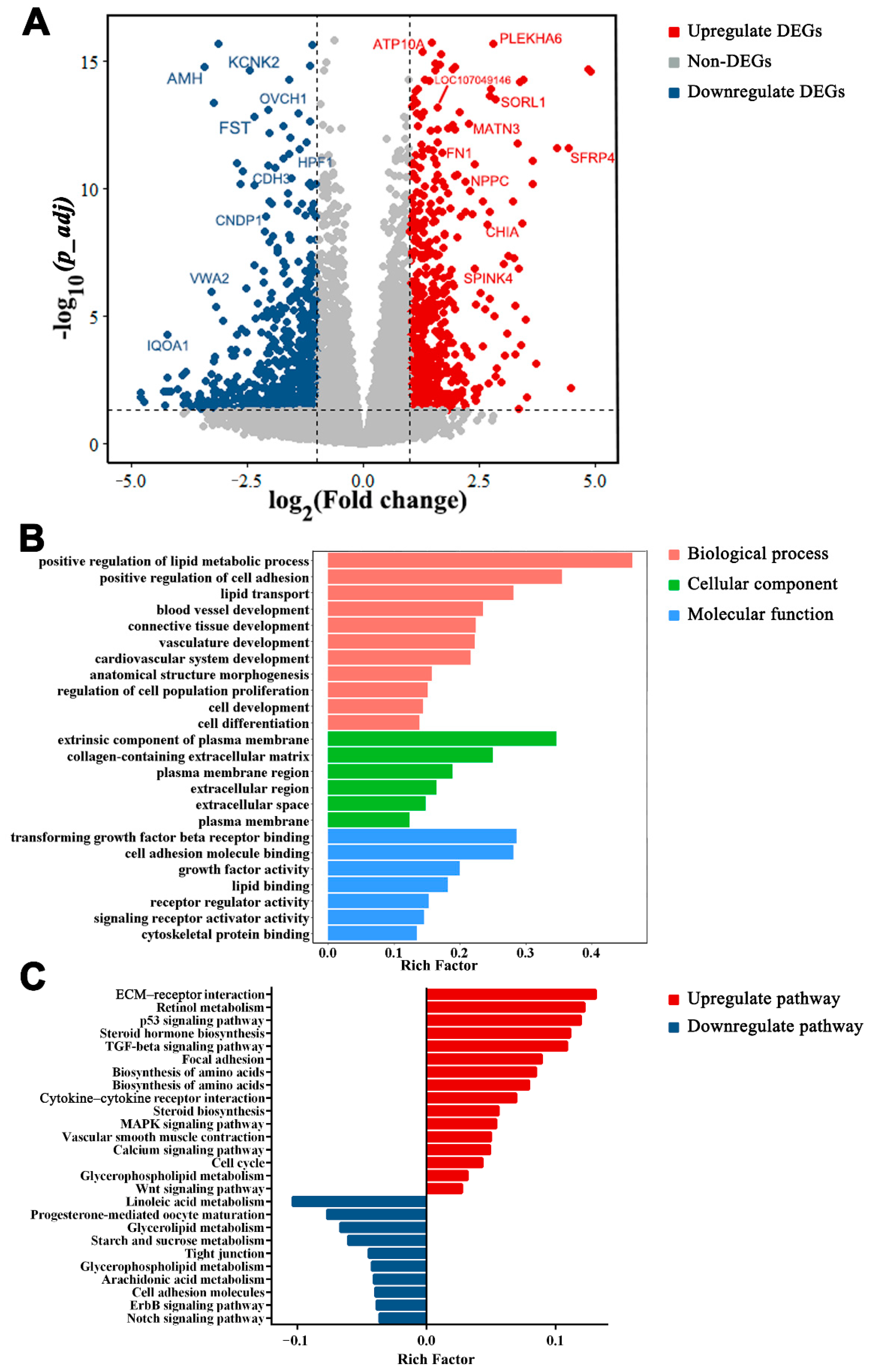
3.5. Functions of DEGs between LWF and SYF
3.6. Functions of DEGs between SYF and LYF
3.7. Functions of DEGs between LWF and LYF
3.8. Gene Expression Dynamics and Transcriptional Profiles around Follicle Selection
3.9. Key Signaling Pathways in Follicle Selection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, P. Follicle Selection in the Avian Ovary. Reprod. Domest. Anim. 2012, 47, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Onagbesan, O.; Bruggeman, V.; Decuypere, E. Intra-ovarian growth factors regulating ovarian function in avian species: A review. Anim. Reprod. Sci. 2009, 111, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L. Ovarian follicle selection and granulosa cell differentiation. Poult. Sci. 2015, 94, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Woods, D.C. Dynamics of avian ovarian follicle development: Cellular mechanisms of granulosa cell differentiation. Gen. Comp. Endocrinol. 2009, 163, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A.; Stephens, C.S.; Giles, J.R. The domestic chicken: Causes and consequences of an egg a day. Poult. Sci. 2015, 94, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods. 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.-X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Software, G. One-Way ANOVA Followed by Dunnett’s Multiple Comparisons Test Was Performed Using GraphPad Prism Version 8.0.0 for Windows; Graph Pad Software Inc.: San Diego, CA, USA, 2020. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Rangel, P.L.; Rodríguez, A.; Gutiérrez, K.; Sharp, P.J.; Gutierrez, C.G. Subdominant hierarchical ovarian follicles are needed for steroidogenesis and ovulation in laying hens (Gallus domesticus). Anim. Reprod. Sci. 2014, 147, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Steroidogenesis: Unanswered Questions. Trends Endocrinol. Metab. 2017, 28, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Q.; Liu, Z.; Guo, X.; Du, Y.; Yuan, Z.; Guo, M.; Kang, L.; Sun, Y.; Jiang, Y. Transcriptome Analysis on Single Small Yellow Follicles Reveals That Wnt4 Is Involved in Chicken Follicle Selection. Front. Endocrinol. 2017, 8, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Wang, Y.; Liu, Z.; Guo, X.; Sun, Y.; Kang, L.; Jiang, Y. Transcriptomic and proteomic analyses of ovarian follicles reveal the role of VLDLR in chicken follicle selection. BMC Genom. 2020, 21, 486. [Google Scholar] [CrossRef] [PubMed]
- Orisaka, M.; Tajima, K.; Tsang, B.K.; Kotsuji, F. Oocyte-granulosa-theca cell interactions during preantral follicular development. J. Ovarian Res. 2009, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braw-Tal, R. The initiation of follicle growth: The oocyte or the somatic cells? Mol. Cell. Endocrinol. 2002, 187, 11–18. [Google Scholar] [CrossRef]
- van der Klein, S.A.S.; Zuidhof, M.J.; Bédécarrats, G.Y. Diurnal and seasonal dynamics affecting egg production in meat chickens: A review of mechanisms associated with reproductive dysregulation. Anim. Reprod. Sci. 2020, 213, 106257. [Google Scholar] [CrossRef] [PubMed]
- Stephens, C.S.; Johnson, P.A. Occludin expression and regulation in small follicles of the layer and broiler breeder hen. Gen. Comp. Endocrinol. 2017, 248, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Hsieh, M.; Doyle, K.H.; Falender, A.E.; Lo, Y.K.; Sharma, S.C. Novel signaling pathways that control ovarian follicular development, ovulation, and luteinization. Rcent. Prog. Horm. Res. 2002, 57, 195–220. [Google Scholar] [CrossRef] [PubMed]
- El-Derany, M.O.; Said, R.S.; El-Demerdash, E. Bone Marrow-Derived Mesenchymal Stem Cells Reverse Radiotherapy-Induced Premature Ovarian Failure: Emphasis on Signal Integration of TGF-β, Wnt/β-Catenin and Hippo Pathways. Stem Cell Rev. Rep. 2021, 17, 1429–1445. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-Y.; O’Connor, A.; Shitanaka, M.; Shimada, M.; Liu, Z.; Richards, J.S. β-Catenin (CTNNB1) Promotes Preovulatory Follicular Development but Represses LH-Mediated Ovulation and Luteinization. Mol. Endocrinol. 2010, 24, 1529–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.-U.; Yoon, J.D.; Kim, M.; Cai, L.; Choi, H.; Oh, D.; Kim, E.; Hyun, S.-H. R-Spondin 2 and WNT/CTNNB1 Signaling Pathways Are Required for Porcine Follicle Development and In Vitro Maturation. Animals 2021, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Vainio, S.; Heikkilä, M.; Kispert, A.; Chin, N.; McMahon, A.P. Female development in mammals is regulated by Wnt-4 signalling. Nature 1999, 397, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Descamps, B.; Sewduth, R.; Ferreira Tojais, N.; Jaspard, B.; Reynaud, A.; Sohet, F.; Lacolley, P.; Allières, C.; Lamazière, J.-M.D.; Moreau, C.; et al. Frizzled 4 Regulates Arterial Network Organization Through Noncanonical Wnt/Planar Cell Polarity Signaling. Circ. Res. 2012, 110, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Wang, Y.; Dabdoub, A.; Smallwood, P.M.; Williams, J.; Woods, C.; Kelley, M.W.; Jiang, L.; Tasman, W.; Zhang, K.; et al. Vascular Development in the Retina and Inner Ear: Control by Norrin and Frizzled-4, a High-Affinity Ligand-Receptor Pair. Cell 2004, 116, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, T.; Kashiwagi, E.; Ide, H.; Li, Y.; Zheng, Y.; Miyamoto, Y.; Netto, G.J.; Ishiguro, H.; Miyamoto, H. Cyclosporine A and tacrolimus inhibit bladder cancer growth through down-regulation of NFATc1. Oncotarget 2015, 6, 1582–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, T.; Kashiwagi, E.; Ide, H.; Li, Y.; Zheng, Y.; Ishiguro, H.; Miyamoto, H. The role of NFATc1 in prostate cancer progression: Cyclosporine A and tacrolimus inhibit cell proliferation, migration, and invasion. Prostate 2015, 75, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Li, J.; Zhang, W.; Ren, F.; Yue, W. NFATc1 activation promotes the invasion of U251 human glioblastoma multiforme cells through COX-2. Int. J. Mol. Med. 2015, 35, 1333–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocón-Grove, O.M.; Poole, D.H.; Johnson, A.L. Bone morphogenetic protein 6 promotes FSH receptor and anti-Müllerian hormone mRNA expression in granulosa cells from hen prehierarchal follicles. Reproduction 2012, 143, 825–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Ocón-Grove, O.; Johnson, A.L. Bone Morphogenetic Protein 4 Supports the Initial Differentiation of Hen (Gallus gallus) Granulosa Cells. Biol. Reprod. 2013, 88, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A.; Kent, T.R.; Urick, M.E.; Giles, J.R. Expression and Regulation of Anti-Mullerian Hormone in an Oviparous Species, the Hen1. Biol. Reprod. 2008, 78, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.J.; Purevsuren, L.; Jin, F.; Zhang, Y.P.; Liang, C.Y.; Zhu, M.Q.; Wang, F.; Jia, C.L.; Wei, Z.H. Effect of anti-müllerian hormone on the development and selection of ovarian follicle in hens. Poult. Sci. 2021, 100, 100959. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y. Control of ovarian primordial follicle activation. Clin. Exp. Reprod. Med. 2012, 39, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaivo-oja, N.; Jeffery, L.A.; Ritvos, O.; Mottershead, D.G. Smad signalling in the ovary. Reprod. Biol. Endocrinol. 2006, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.H.; Vanderhyden, B.C. Oocyte-granulosa cell interactions during mouse follicular development: Regulation of kit ligand expression and its role in oocyte growth. Reprod. Biol. Endocrinol. 2006, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.-M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell. 2018, 72, 1021–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L. Stage of Ovarian Follicular Development Associated with the Initiation of Steroidogenic Competence in Avian Granulosa Cells1. Biol. Reprod. 1991, 44, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Chandra, P.; Goyal, R.N.; Shim, Y.-B. A review on determination of steroids in biological samples exploiting nanobio-electroanalytical methods. Anal. Chim. Acta. 2013, 762, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Solovieva, E.V.; Bridgham, J.T. Relationship Between Steroidogenic Acute Regulatory Protein Expression and Progesterone Production in Hen Granulosa Cells During Follicle Development1. Biol. Reprod. 2002, 67, 1313–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, K.; Asiamah, C.A.; Lu, L.-L.; Liu, Y.; Pan, Y.; Chen, T.; Zhao, Z.; Su, Y. Ovarian transcriptomic analysis and follicular development of Leizhou black duck. Poult. Sci. 2020, 99, 6173–6187. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Gu, T.; Zhang, Y.; Huang, Z.; Wu, N.; Zhao, W.; Chang, G.; Xu, Q.; Chen, G. Transcriptome profiling to identify key mediators of granulosa cell proliferation upon FSH stimulation in the goose (Anser cygnoides). Br. Poult. Sci. 2018, 59, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Waterman Michael, R. A Rising StAR: An Essential Role in Cholesterol Transport. Science 1995, 267, 1780–1781. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.K.; Strauss, J.F. Steroidogenic acute regulatory protein (StAR) and the intramitochondrial translocation of cholesterol. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2000, 1529, 175–187. [Google Scholar] [CrossRef]
- Sugawara, T.; Lin, D.; Holt, J.A.; Martin, K.O.; Javitt, N.B.; Miller, W.L.; Strauss, J.F., 3rd. Structure of the human steroidogenic acute regulatory protein (StAR) gene: StAR stimulates mitochondrial cholesterol 27-hydroxylase activity. Biochemistry 1995, 34, 12506–12512. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.J.; Soo, S.C.; Caron, K.M.; Ikeda, Y.; Parker, K.L.; Stocco, D.M. Hormonal and developmental regulation of the steroidogenic acute regulatory protein. Mol. Endocrinol. 1995, 9, 1346–1355. [Google Scholar]
- Sugawara, T.; Kiriakidou, M.; McAllister, J.M.; Holt, J.A.; Arakane, F.; Strauss, J.F. Regulation of expression of the steroidogenic acute regulatory protein (StAR) gene: A central role for steroidogenic factor 1. Steroids 1997, 62, 5–9. [Google Scholar] [CrossRef]
- Wooton-Kee, C.R.; Clark, B.J. Steroidogenic Factor-1 Influences Protein-Deoxyribonucleic Acid Interactions within the Cyclic Adenosine 3′,5′-Monophosphate-Responsive Regions of the Murine Steroidogenic Acute Regulatory Protein Gene. Endocrinology 2000, 141, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Viger, R.S.; Mertineit, C.; Trasler, J.M.; Nemer, M. Transcription factor GATA-4 is expressed in a sexually dimorphic pattern during mouse gonadal development and is a potent activator of the Müllerian inhibiting substance promoter. Development 1998, 125, 2665–2675. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Holt, J.A.; Kiriakidou, M.; Strauss, J.F. Steroidogenic Factor 1-Dependent Promoter Activity of the Human Steroidogenic Acute Regulatory Protein (StAR) Gene. Biochemistry 1996, 35, 9052–9059. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, T.W.; Hales, D.B.; Hales, K.H.; McLean, M.P. Transcriptional Regulation of the Rat Steroidogenic Acute Regulatory Protein Gene by Steroidogenic Factor 1. Endocrinology 1998, 139, 4820–4831. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.; Liu, H.-C.; Hicks, J.A.; Long, J.A.; Porter, T.E. Transcriptome Analysis During Follicle Development in Turkey Hens With Low and High Egg Production. Front. Genet. 2021, 12, 619196. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Bridgham, J.T. Regulation of Steroidogenic Acute Regulatory Protein and Luteinizing Hormone Receptor Messenger Ribonucleic Acid in Hen Granulosa Cells. Endocrinology 2001, 142, 3116–3124. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LWF | SYF | LYF | p-Value | |
|---|---|---|---|---|
| Quantity (number) n 1 | 11.83 ± 2.79 a | 6.83 ± 2.23 b | 1 c | <0.001 |
| Weight (g) 2 | 0.07 ± 0.02 c | 0.16 ± 0.06 b | 0.93 ± 0.19 a | <0.001 |
| Major diameter (mm) 3 | 5.23 ± 0.53 c | 6.92 ± 0.91 b | 13.02 ± 0.61 a | <0.001 |
| Minor diameter (mm) 4 | 4.71 ± 0.49 c | 6.13 ± 0.81 b | 11.61 ± 0.84 a | <0.001 |
| Average diameter (mm) 5 | 4.97 ± 0.49 c | 6.55 ± 0.81 b | 12.31 ± 0.67 a | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, R.; Zheng, X.; Zhang, W.; Zhang, B.; Ling, Y.; Zhang, H.; Wu, C. Morphological Characteristics and Transcriptome Landscapes of Chicken Follicles during Selective Development. Animals 2022, 12, 713. https://doi.org/10.3390/ani12060713
Nie R, Zheng X, Zhang W, Zhang B, Ling Y, Zhang H, Wu C. Morphological Characteristics and Transcriptome Landscapes of Chicken Follicles during Selective Development. Animals. 2022; 12(6):713. https://doi.org/10.3390/ani12060713
Chicago/Turabian StyleNie, Ruixue, Xiaotong Zheng, Wenhui Zhang, Bo Zhang, Yao Ling, Hao Zhang, and Changxin Wu. 2022. "Morphological Characteristics and Transcriptome Landscapes of Chicken Follicles during Selective Development" Animals 12, no. 6: 713. https://doi.org/10.3390/ani12060713