
Overexpression of DGAT2 Stimulates Lipid Droplet Formation and Triacylglycerol Accumulation in Bovine Satellite Cells

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Adenovirus Generation and Overexpression of DGAT2
2.3. Design of shRNAs and Recombinant Adenovirus Production
2.4. Optimization of Optimal Conditions for Adenoviral Overexpression/Interference of the DGAT2 Gene
2.5. Cell Culture and Treatment
2.6. RNA Extraction and Quantitative Real-Time PCR
2.7. Oil Red O Staining
2.8. Cellular TAG Assay
2.9. Adiponectin Assay in Culture Medium
2.10. Western Blotting
2.11. Transcriptome Sequencing (RNA-Seq) Research
2.12. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis of DEGs
2.13. KEGG Enrichment Analysis
2.14. Statistical Analysis
3. Results
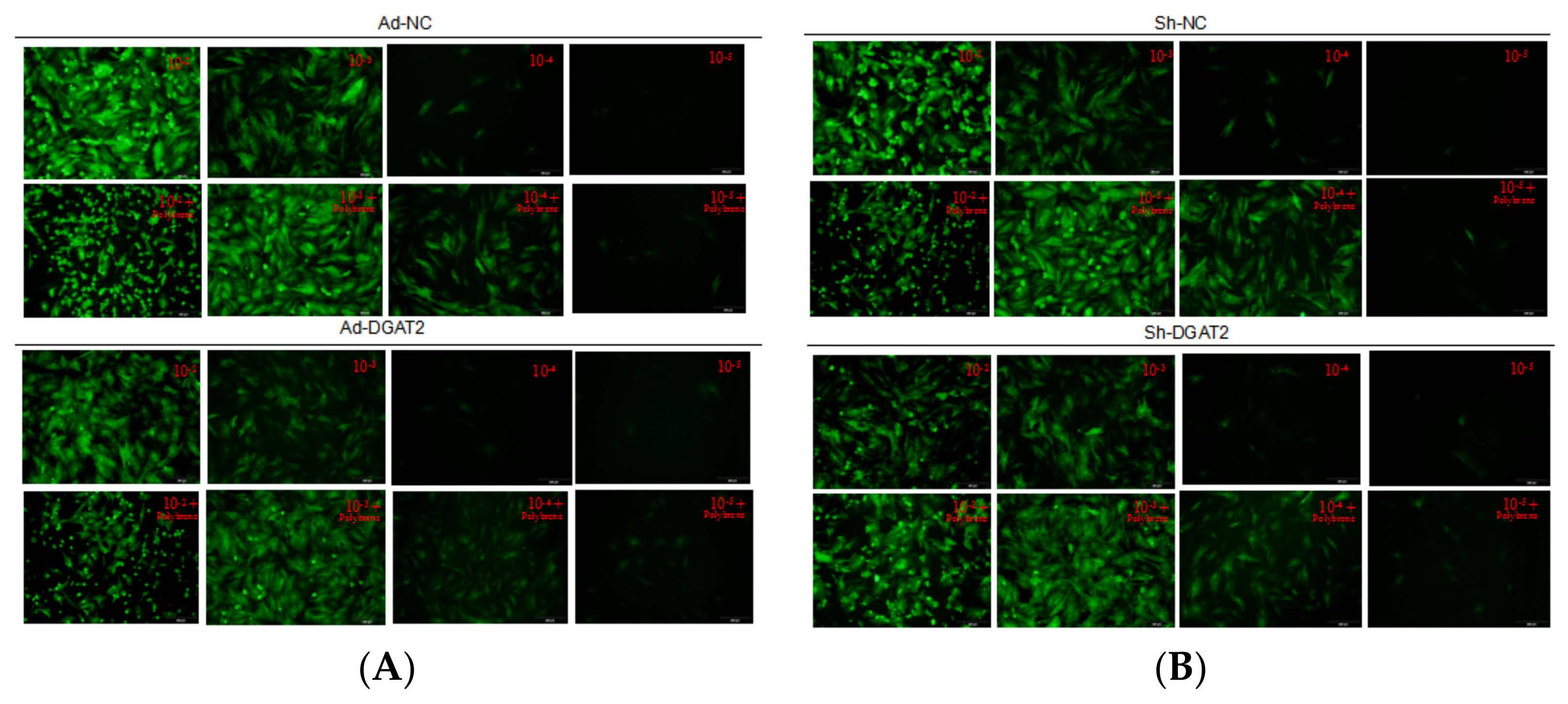
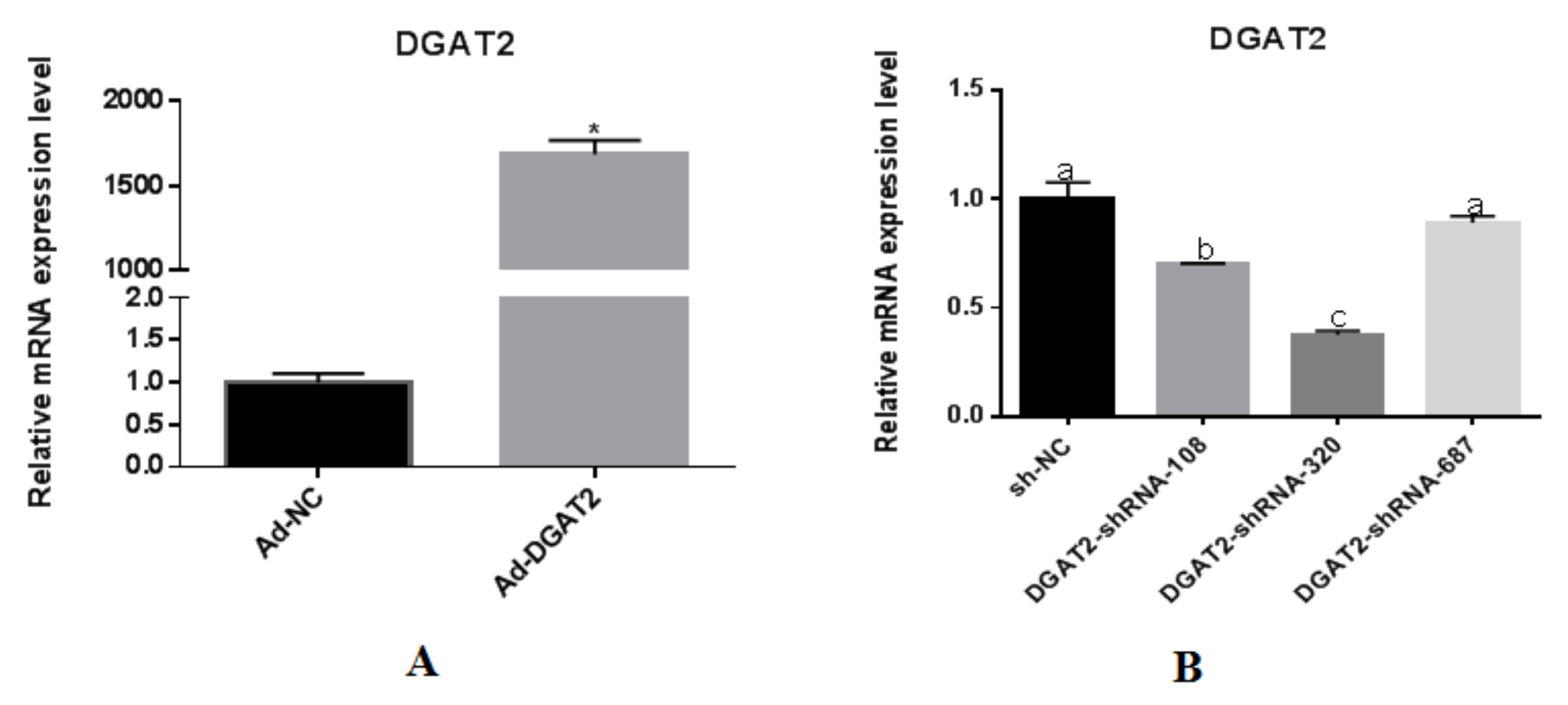
3.1. Detection of Optimal MOI Value and Infection Efficiency in Yanbian Bovine Skeletal Muscle Satellite Cells Infected with Adenovirus Overexpressing or Interfering with DGAT2
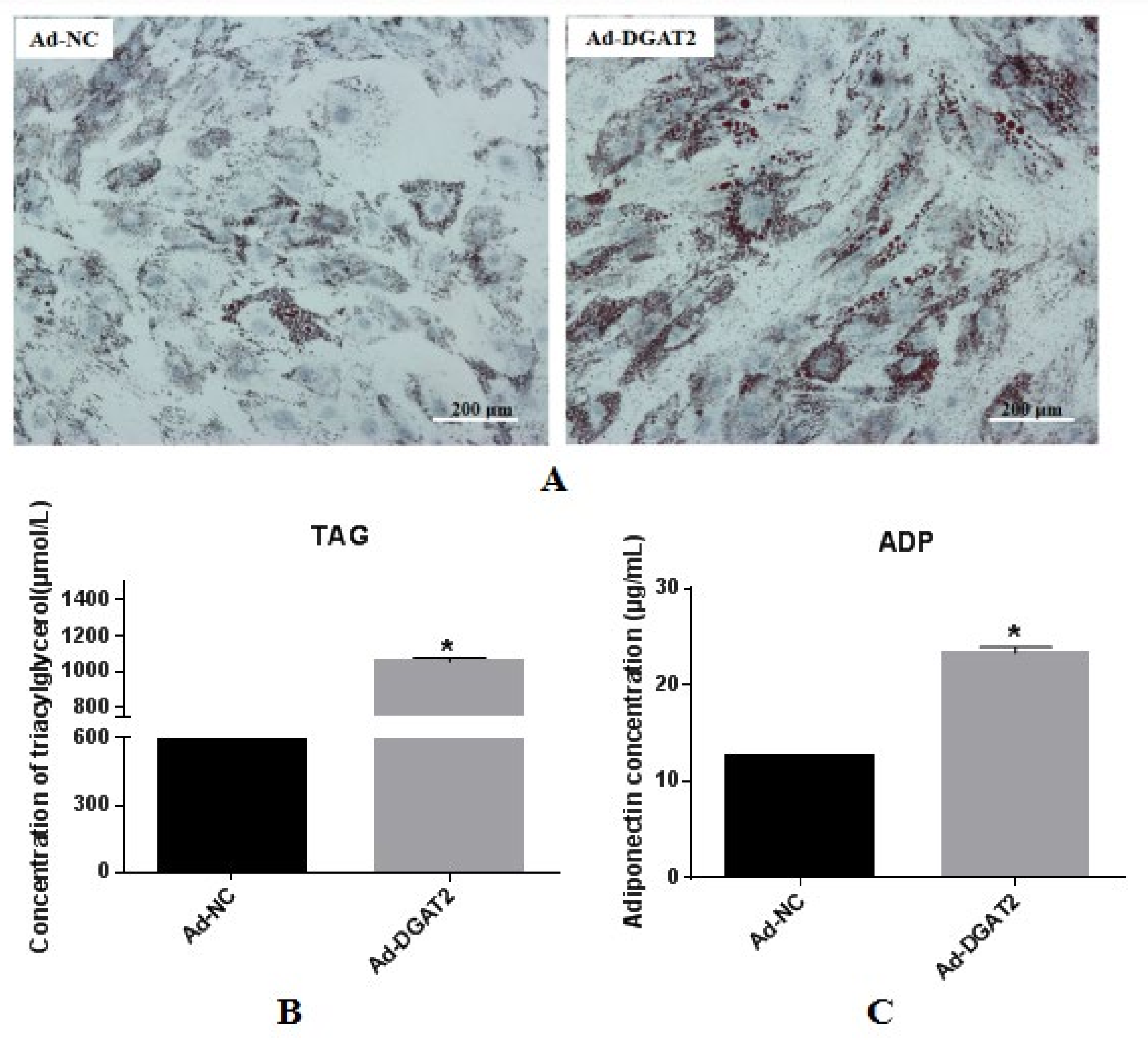
3.2. Overexpression of DGAT2 Increases TAG Accumulation, ADP Content, and Lipid Droplet Formation in BSCs Undergoing Adipogenic Transdifferentiation
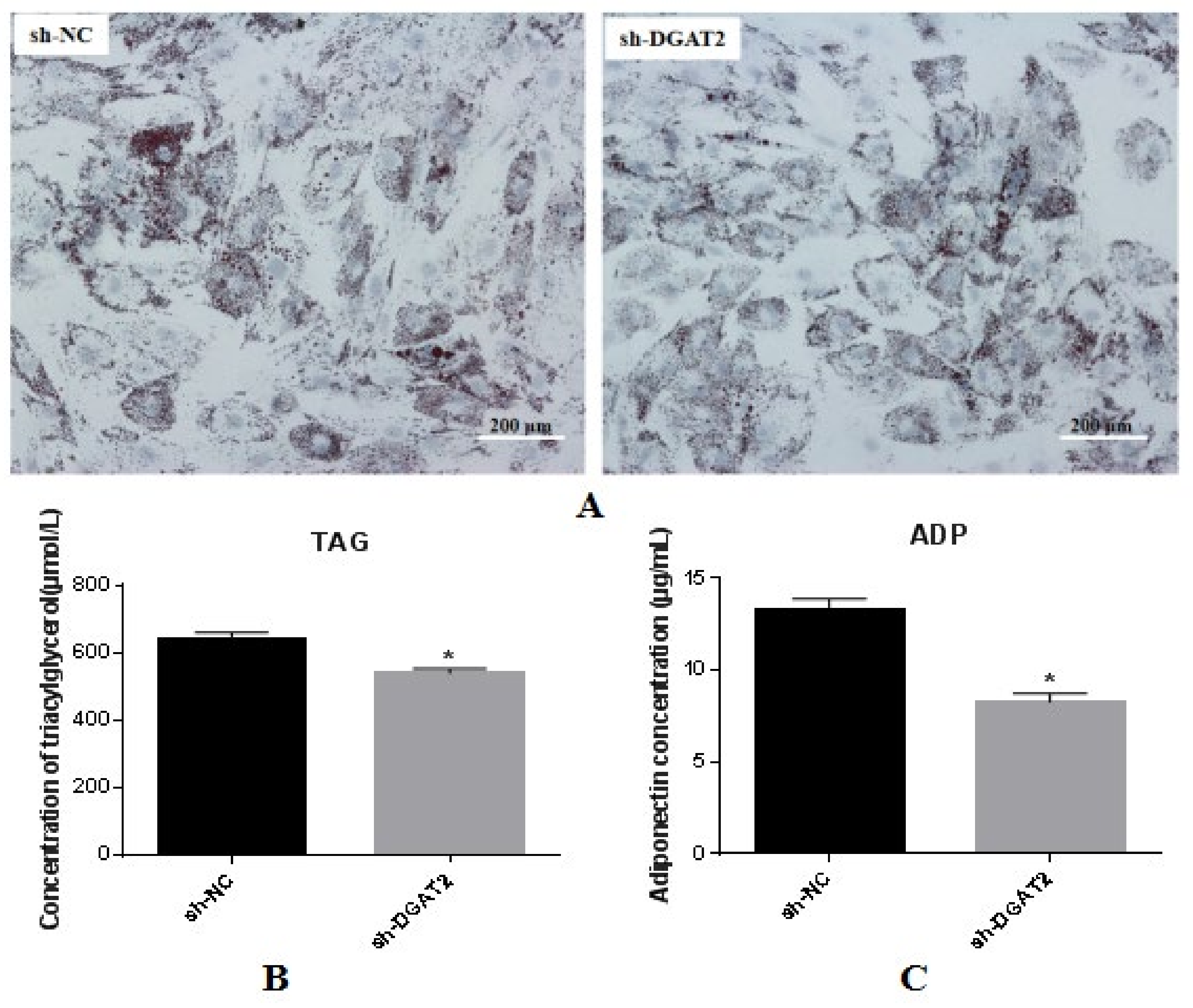
3.3. Interference of DGAT2 Affects TAG Accumulation, ADP Content, and Lipid Droplet Formation in BSCs Undergoing Adipogenic Transdifferentiation
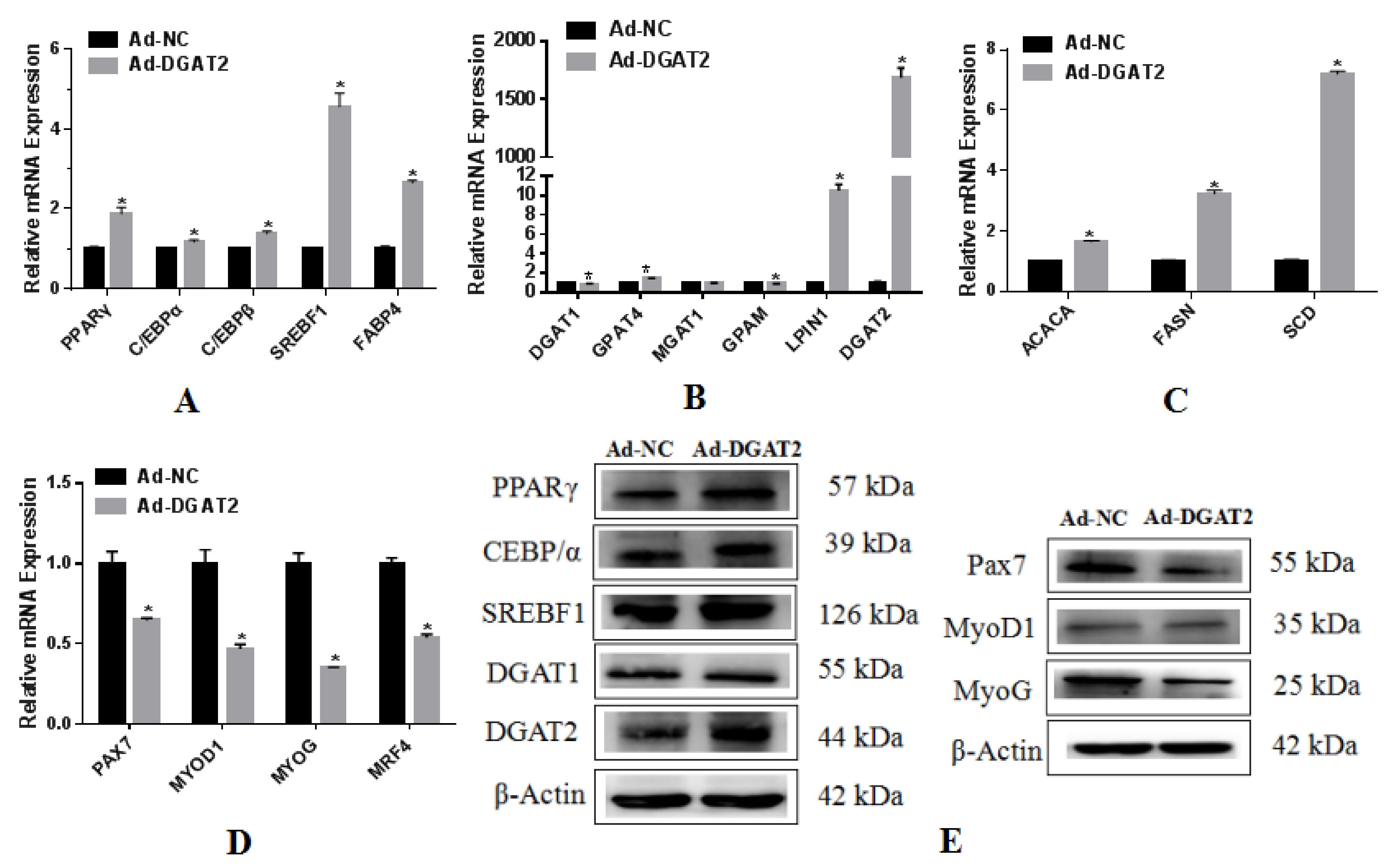
3.4. The Expression and Function of DGAT2 Overexpression in BSCs Adipogenic Differentiation
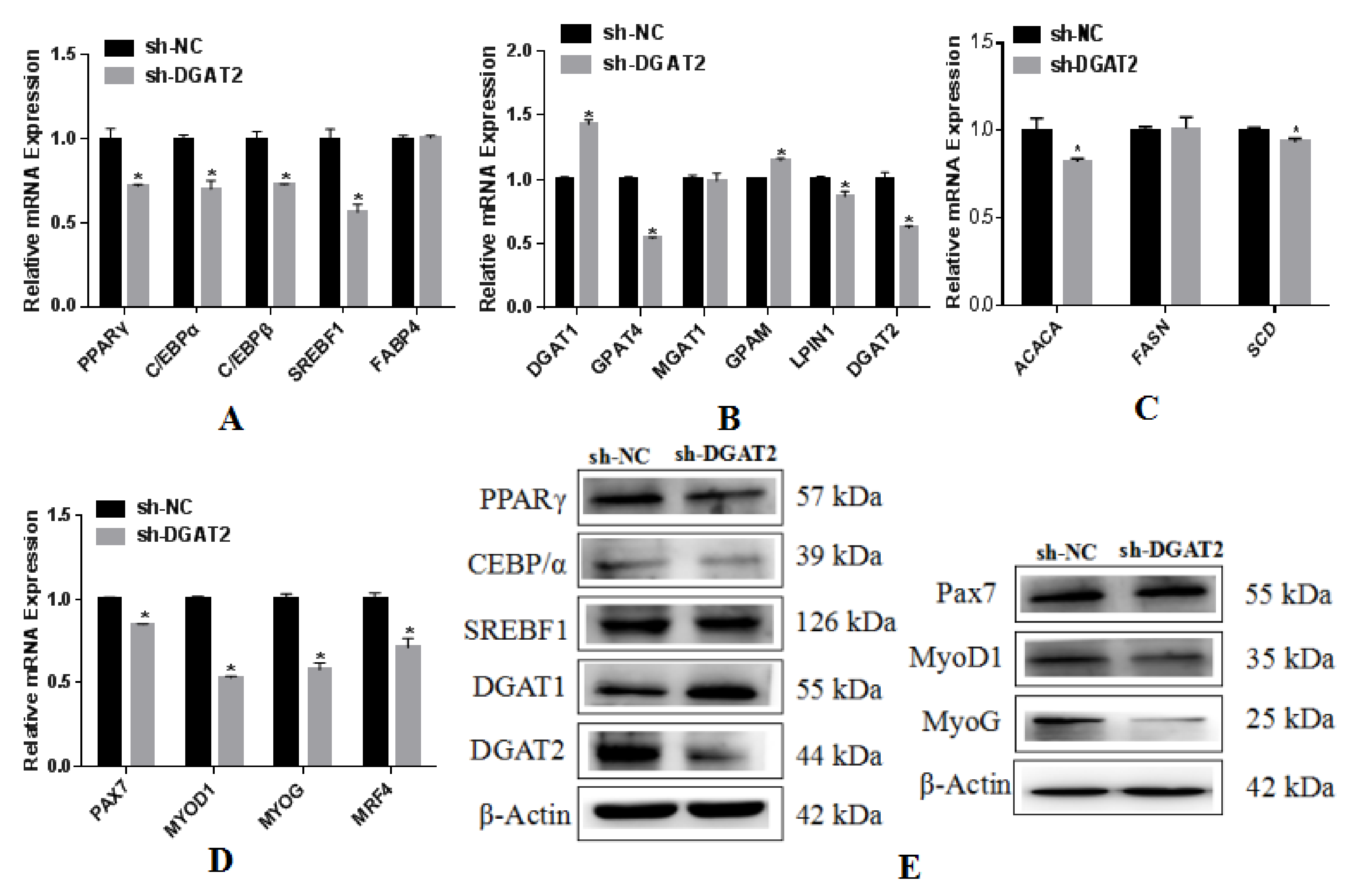
3.5. Effects of Interference with DGAT2 Expression on BSCs Undergoing Adipogenic Differentiation
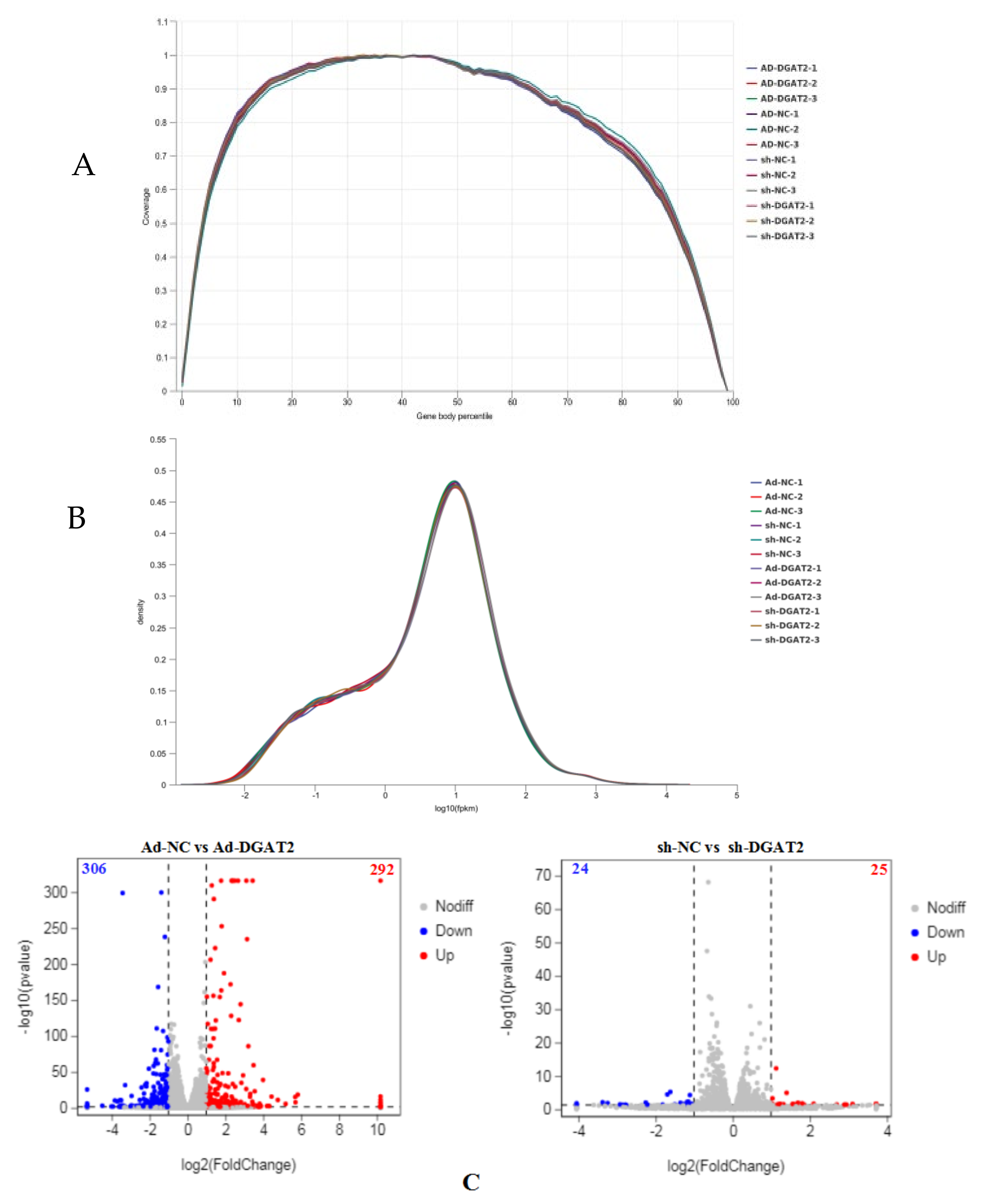
3.6. DEG Screening in BSCs Infected with Ad-DGAT2/sh-DGAT2
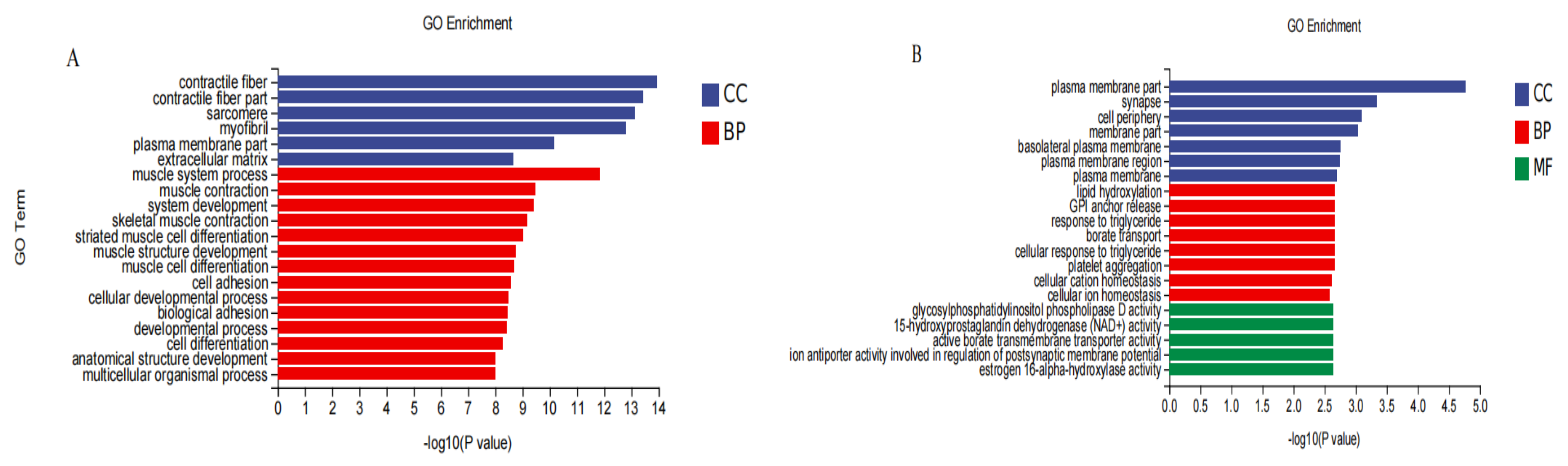
3.7. GO Functional Classification
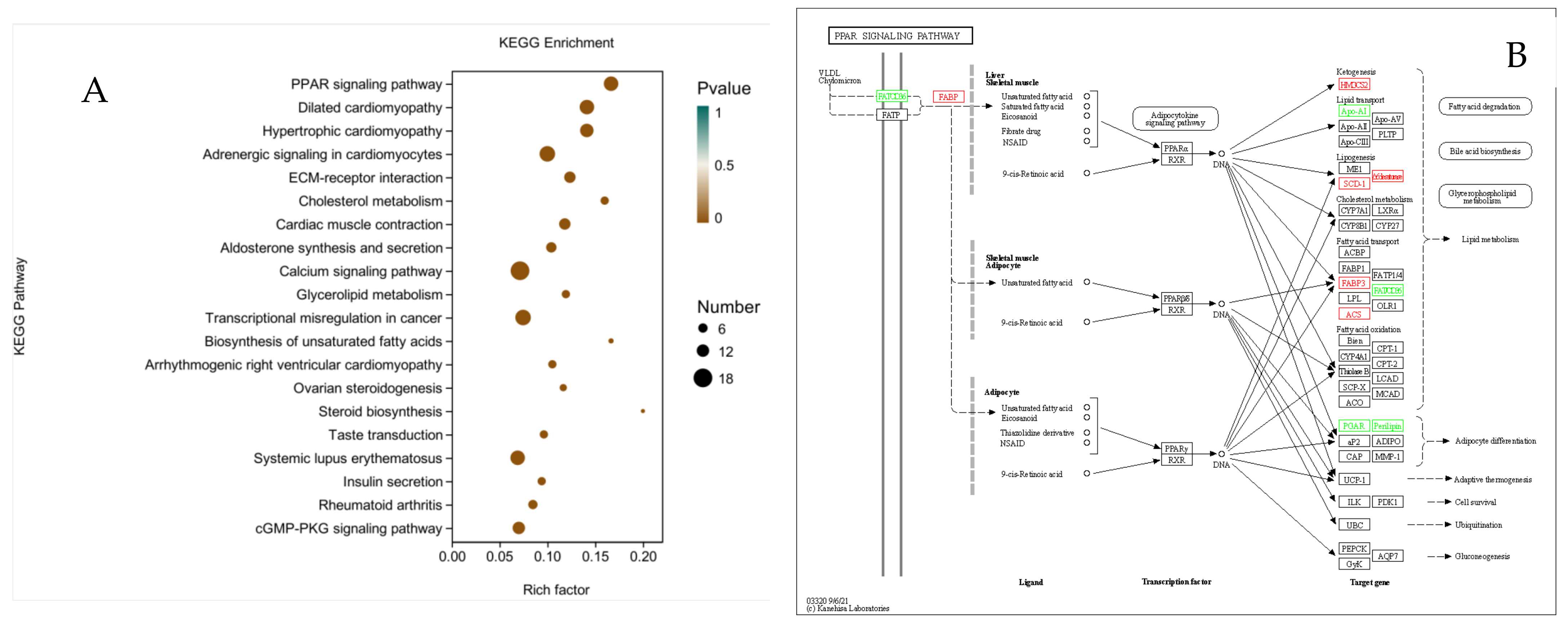
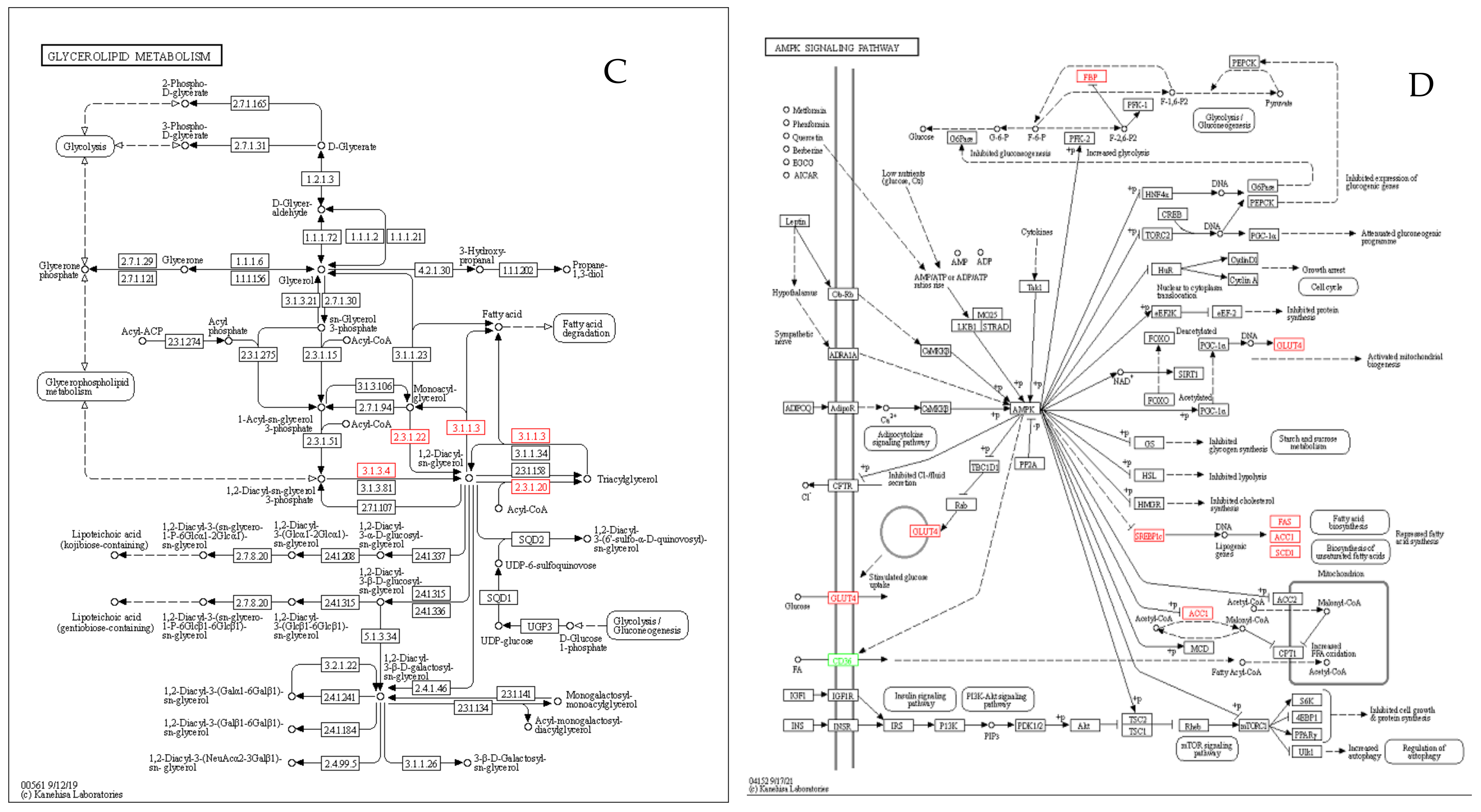
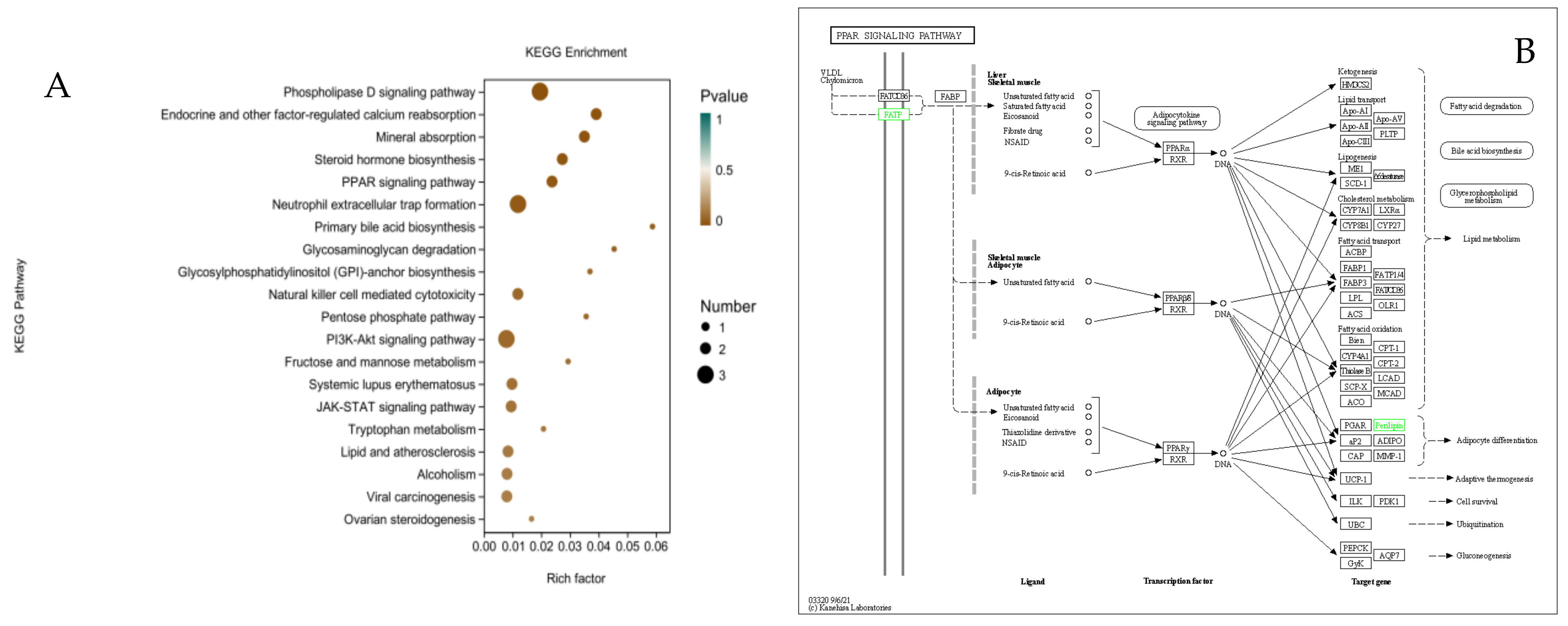
3.8. KEGG Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hamrick, M.W.; McGee-Lawrence, M.E.; Frechette, D.M. Fatty infiltration of skeletal muscle: Mechanisms and comparisons with bone marrow adiposity. Front. Endocrinol. 2016, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uezumi, A.; Fukada, S.-I.; Yamamoto, N.; Takeda, S.; Tsuchida, K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nat. Cell Biol. 2010, 12, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Yada, E.; Yamanouchi, K.; Nishihara, M. Adipogenic potential of satellite cells from distinct skeletal muscle origins in the rat. J. Vet. Med. Sci. 2006, 68, 479–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Billecke, N.; Bosma, M.; Rock, W.; Fleissner, F.; Best, G.; Schrauwen, P.; Kersten, S.; Bonn, M.; Hesselink, M.K.C.; Parekh, S.H. Perilipin 5 mediated lipid droplet remodelling revealed by coherent Raman imaging. Integr. Biol. 2015, 7, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, K.; Lee, Y.K.; Londos, C.; Raaka, B.M.; Dalen, K.T.; Kimmel, A.R. Perilipin family members preferentially sequester to either triacylglycerol-specific or cholesteryl-ester-specific intracellular lipid storage droplets. J. Cell Sci. 2012, 125 Pt 17, 4067–4076. [Google Scholar] [CrossRef] [Green Version]
- Coleman, R.A.; Lee, D.P. Enzymes of triacylglycerol synthesis and their regulation. Prog. Lipid Res. 2004, 43, 134–176. [Google Scholar] [CrossRef]
- Yen, C.L.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V., Jr. Thematic review series: Glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J. Lipid Res. 2008, 49, 2283–2301. [Google Scholar] [CrossRef] [Green Version]
- Chitraju, C.; Mejhert, N.; Haas, J.T.; Diaz-Ramirez, L.G.; Grueter, C.A.; Imbriglio, J.E.; Pinto, S.; Koliwad, S.K.; Walther, T.C.; Farese, R.V. Triglyceride synthesis by DGAT1 protects adipocytes from lipid-induced ER stress during lipolysis. Cell Metab. 2017, 26, 407–418.e3. [Google Scholar] [CrossRef]
- Smith, S.J.; Cases, S.; Jensen, D.R.; Chen, H.C.; Sande, E.; Tow, B.; Sanan, D.A.; Raber, J.; Eckel, R.H.; Farese, R.V. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nat. Genet. 2000, 25, 87–90. [Google Scholar] [CrossRef]
- Stone, S.J.; Myers, H.M.; Watkins, S.M.; Brown, B.E.; Feingold, K.R.; Elias, P.M.; Farese, R.V. Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J. Biol. Chem. 2004, 279, 11767–11776. [Google Scholar] [CrossRef] [Green Version]
- Stone, S.J.; Levin, M.C.; Zhou, P.; Han, J.; Walther, T.C.; Farese, R.V., Jr. The endoplasmic reticulum enzyme DGAT2 is found in mitochondria-associated membranes and has a mitochondrial targeting signal that promotes its association with mitochondria. J. Biol. Chem. 2009, 284, 5352–5361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilfling, F.; Wang, H.; Haas, J.T.; Krahmer, N.; Gould, T.J.; Uchida, A.; Cheng, J.-X.; Graham, M.; Christiano, R.; Fröhlich, F.; et al. Triacylglycerol synthesis enzymes mediate lipid droplet growth by relocalizing from the ER to lipid droplets. Dev. Cell 2013, 24, 384–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Z.; Yan, Y.; Zhang, J.F.; Sun, J.F.; Sun, B.; Yan, C.G.; Choi, S.H.; Johnson, B.J.; Kim, J.K.; Smith, S.B. Oleic acid in the absence of a PPARγ agonist increases adipogenic gene expression in bovine muscle satellite cells1. J. Anim. Sci. 2019, 97, 4114–4123. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Q.; Yan, Y.; Sun, B.; Wang, Y.; Tang, L.; Wang, E.; Yu, J.; Nogoy, K.M.C.; Li, X.Z.; et al. Effect of ciglitazone on adipogenic transdifferentiation of bovine skeletal muscle satellite cells. J. Anim. Sci. Technol. 2021, 63, 934–953. [Google Scholar] [CrossRef] [PubMed]
- Oppi-Williams, C.; Suagee, J.K.; Corl, B.A. Regulation of lipid synthesis by liver X receptor α and sterol regulatory element-binding protein 1 in mammary epithelial cells. J. Dairy Sci. 2013, 96, 112–121. [Google Scholar] [CrossRef]
- Vela-Vásquez, D.A.; Sifuentes-Rincón, A.M.; Delgado-Enciso, I.; Delgado-Enciso, O.G.; Ordaz-Pichardo, C.; Arellano-Vera, W.; Treviño-Alvarado, V. Improvement of serum lipid parameters in consumers of Mexican Wagyu-Cross beef: A randomized controlled trial. J. Food Sci. 2021, 86, 2713–2726. [Google Scholar] [CrossRef]
- Ruiz-Núñez, B.; Kuipers, R.S.; Luxwolda, M.F.; De Graaf, D.J.; Breeuwsma, B.B.; Dijck-Brouwer, D.J.; Muskiet, F.A. Saturated fatty acid (SFA) status and SFA intake exhibit different relations with serum total cholesterol and lipoprotein cholesterol: A mechanistic explanation centered around lifestyle-induced low-grade inflammation. J. Nutr. Biochem. 2014, 25, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Satija, A.; Blondin, S.; Janiszewski, M.; Emlen, E.; O’Connor, L.E.; Campbell, W.W.; Hu, F.B.; Willett, W.C.; Stampfer, M.J. Meta-analysis of randomized controlled trials of red meat consumption in comparison with various comparison diets on cardiovascular risk factors. Circulation 2019, 139, 1828–1845. [Google Scholar] [CrossRef]
- Williams, K.J.; Bensinger, S.J. Cellular fatty acid analysis in macrophage using stable isotope labeling. Methods Mol. Biol. 2020, 2184, 47–60. [Google Scholar] [CrossRef]
- Cases, S.; Smith, S.J.; Zheng, Y.-W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welch, C.B.; Lusis, A.J.; et al. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [Green Version]
- Asakura, A.; Komaki, M.; Rudnicki, M. Muscle satellite cells are multipotential stem cells that exhibit myogenic, osteogenic, and adipogenic differentiation. Differentiation 2001, 68, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Q.; Nogoy, K.M.; Sun, J.; Sun, B.; Wang, Y.; Tang, L.; Yu, J.; Jin, X.; Choi, S.H.; et al. Effect of palmitoleic acid on the differentiation of bovine skeletal muscle satellite cells. J. Anim. Sci. Technol. 2021, 63, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Belal, S.A.; Kang, D.R.; Sivakumar, A.S.; Choe, H.S.; Shim, K.S. Effect of long chain fatty acids on triacylglycerol accumulation, fatty acid composition and related gene expression in primary cultured bovine satellite cells. Anim. Biotechnol. 2019, 30, 323–331. [Google Scholar] [CrossRef]
- McFie, P.J.; Banman, S.L.; Kary, S.; Stone, S.J. Murine diacylglycerol acyltransferase-2 (DGAT2) can catalyze triacylglycerol synthesis and promote lipid droplet formation independent of its localization to the endoplasmic reticulum. J. Biol. Chem. 2011, 286, 28235–28246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Shu, G.; Zhu, X.; Guo, J.; Cai, H.; Wang, S.; Wang, L.; Gao, P.; Xi, Q.; Zhang, Y.; et al. Effect of diacylglycerol acyltransferase 2 overexpression in 3T3-L1 is associated to an increase in mono-unsaturated fatty acid accumulation. J. Anim. Sci. Biotechnol. 2014, 5, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.C.; Monetti, M.; Watt, M.J.; Sajan, M.P.; Stevens, R.D.; Bain, J.R.; Newgard, C.B.; Farese, R.V. Increased lipid accumulation and insulin resistance in transgenic mice expressing DGAT2 in glycolytic (type II) muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1772–E1781. [Google Scholar] [CrossRef] [Green Version]
- Man, W.C.; Miyazaki, M.; Chu, K.; Ntambi, J. Colocalization of SCD1 and DGAT2: Implying preference for endogenous monounsaturated fatty acids in triglyceride synthesis. J. Lipid Res. 2006, 47, 1928–1939. [Google Scholar] [CrossRef] [Green Version]
- Solé, E.; Ros-Freixedes, R.; Tor, M.; Pena, R.N.; Estany, J. A sequence variant in the diacylglycerol O-acyltransferase 2 gene influences palmitoleic acid content in pig muscle. Sci. Rep. 2021, 11, 14797. [Google Scholar] [CrossRef]
- Hu, S.; Gao, S.; Zhu, J.; Gan, X.; Chen, X.; He, H.; Liang, L.; Hu, B.; Hu, J.; Liu, H.; et al. Differential actions of diacylglycerol acyltransferase (DGAT) 1 and 2 in regulating lipid metabolism and progesterone secretion of goose granulosa cells. J. Steroid Biochem. Mol. Biol. 2020, 202, 105721. [Google Scholar] [CrossRef]
- Smith, A.; Yu, X.; Yin, L. Diazinon exposure activated transcriptional factors CCAAT-enhancer-binding proteins α (C/EBPα) and peroxisome proliferator-activated receptor γ (PPARγ) and induced adipogenesis in 3T3-L1 preadipocytes. Pestic. Biochem. Physiol. 2018, 150, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Fan, X.; Miao, Y. LPIN1 promotes triglycerides synthesis and is transcriptionally regulated by PPARG in buffalo mammary epithelial cells. Sci. Rep. 2022, 12, 2390. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Rui, B.B.; Tang, L.Y.; Hu, C.M. Lipin family proteins—Key regulators in lipid metabolism. Ann. Nutr. Metab. 2015, 66, 10–18. [Google Scholar] [CrossRef]
- Guo, S.; Gong, L.; Shen, Q.; Xing, D. Photobiomodulation reduces hepatic lipogenesis and enhances insulin sensitivity through activation of CaMKKβ/AMPK signaling pathway. J. Photochem. Photobiol. B. 2020, 213, 112075. [Google Scholar] [CrossRef]
- Jung, E.J.; Kwon, S.W.; Jung, B.H.; Oh, S.H.; Lee, B.H. Role of the AMPK/SREBP-1 pathway in the development of orotic acid-induced fatty liver. J. Lipid Res. 2011, 52, 1617–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Kang, J.S.; Park, Y.H.; Je, B.I.; Lee, Y.J.; Kang, N.J.; Park, S.Y.; Hwang, D.Y.; Choi, Y.W. S-petasin inhibits lipid accumulation in oleic acid-induced HepG2 cells through activation of the AMPK signaling pathway. Food Funct. 2020, 11, 5664–5673. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Molina, A.; Lopez-Guadamillas, E.; Mattison, J.A.; Mitchell, S.J.; Muñoz-Martin, M.; Iglesias, G.; Gutierrez, V.M.; Vaughan, K.L.; Szarowicz, M.D.; González-García, I.; et al. Pharmacological inhibition of PI3K reduces adiposity and metabolic syndrome in obese mice and rhesus monkeys. Cell Metab. 2015, 21, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Schoettl, T.; Fischer, I.P.; Ussar, S. Heterogeneity of adipose tissue in development and metabolic function. J. Exp. Biol. 2018, 221 (Suppl. 1), jeb162958. [Google Scholar] [CrossRef] [Green Version]
- Carey, A.L.; Vorlander, C.; Reddy-Luthmoodoo, M.; Natoli, A.K.; Formosa, M.F.; Bertovic, D.A.; Anderson, M.J.; Duffy, S.J.; Kingwell, B.A. Reduced UCP-1 content in in vitro differentiated beige/brite adipocytes derived from preadipocytes of human subcutaneous white adipose tissues in obesity. PLoS ONE 2014, 9, e91997. [Google Scholar] [CrossRef] [Green Version]
- Shan, T.; Liu, W.; Kuang, S. Fatty acid binding protein 4 expression marks a population of adipocyte progenitors in white and brown adipose tissues. FASEB J. 2013, 27, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Schubert, M.F.; Noah, A.C.; Bedi, A.; Gumucio, J.P.; Mendias, C.L. Reduced Myogenic and Increased Adipogenic Differentiation Capacity of Rotator Cuff Muscle Stem Cells. J. Bone Jt. Surg. Am. 2019, 101, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Olguin, H.C.; Yang, Z.; Tapscott, S.J.; Olwin, B.B. Reciprocal inhibition between Pax7 and muscle regulatory factors modulates myogenic cell fate determination. J. Cell Biol. 2007, 177, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Chargé, S.B.; Seale, P.; Huh, M.; Rudnicki, M.A. Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis. J. Cell Biol. 2006, 172, 103–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| shRNA | Sense Strand | Anti-Sense Strand | Target Sequence |
|---|---|---|---|
| shRNA-108 | AATTCGGTAGAGAAGCAGCTCCAAGTTTCAAGAGAACTTGGAGCTGCTTCTCTACCTTTTTTG | GATCCAAAAAAGGTAGAGAAGCAGCTCCAAGTTCTCTTGAAACTTGGAGCTGCTTCTCTACCG | GGTAGAGAAGCAGCTCCAAGT |
| shRNA-320 | AATTCGCTACTTTCGAGACTACTTTCTTCAAGAGAGAAAGTAGTCTCGAAAGTAGCTTTTTTG | GATCCAAAAAAGCTACTTTCGAGACTACTTTCTCTCTTGAAGAAAGTAGTCTCGAAAGTAGCG | GCTACTTTCGAGACTACTTTC |
| shRNA-687 | AATTCGCGCAATCGCAAGGGCTTTGTTTCAAGAGAACAAAGCCCTTGCGATTGCGCTTTTTTG | GATCCAAAAAAGCGCAATCGCAAGGGCTTTGTTCTCTTGAAACAAAGCCCTTGCGATTGCGCG | GCGCAATCGCAAGGGCTTTGT |
| shRNA-NC | AATTCGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAACTTTTTTG | GATCCAAAAAAGTTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAACG | TTCTCCGAACGTGTCACGT |
| Gene Symbol | Gene ID | Primer Sequence (5′–3′) | Length (bp) |
|---|---|---|---|
| PPARγ | NM_181024 | F:ATCTGCTGCAAGCCTTGGA R:TGGAGCAGCTTGGCAAAGA | 138 |
| SREBF1 | NM_001113302 | F:CTGCTGACCGACATAGAAGACAT R:GTAGGGCGGGTCAAACAGG | 81 |
| C/EBPα | NM_176784 | F:CCAGAAGAAGGTGGAGCAACTG R:TCGGGCAGCGTCTTGAAC | 69 |
| C/EBPβ | NM_176788 | F:CAACCTGGAGACGCAGCACAAG R:CGGAGGAGGCGAGCAGAGG | 143 |
| FABP4 | NM_174314 | F:GTGGGCTTTGCTACCAGGAA R:GTGACCACACCCCCATTCAA | 77 |
| DGAT1 | NM_174693 | F:CTACACCATCCTCTTCCTCAAG R:AGTAGTAGAGATCGCGGTAGGTC | 176 |
| DGAT2 | NM_205793 | F:GACCCTCATAGCCTCCTACTCC R:GACCCATTGTAGCACCGAGATGAC | 145 |
| GPAT4 | NM_001083669 | F:AAGCAAGTTGCCCATCCTCA R:AAACTGTGGCTCCAATTTCGA | 101 |
| GPAM | NM_001012282 | F:GCGAACAACTGGGAAAACCC R:GGCAACAATGCTTGCTCCAA | 193 |
| LPIN1 | NM_001206156 | F:AGTCCTCGCCACACAAGATG R:AGATGCCCTGACCAGTGTTG | 137 |
| MGAT1 | NM_001015653 | F:AGCCGTGGTGGTAGAGGATGATC R:TGCTCCTTGCCATTGTCGTTCC | 132 |
| SCD | NM_173959 | F:TGCCCACCACAAGTTTTCAG R:GCCAACCCACGTGAGAGAAG | 80 |
| ACACA | NM_174224 | F: GCCAAACCTCTGGAGCTGAA R: CGAGCTTCACCAGGTTGCTA | 97 |
| FASN | NM_001012669 | F:CGCTTGCTGCTGGAGGTCAC R:GGTCTCAGGGTCTCGGCTCAG | 141 |
| MYOD1 | NM_001040478 | F:CCGACGGCATGATGGACTA R:CTCGCTGTAGTAAGTGCGGT | 80 |
| MYOG | NM_001111325 | F:CAGTGAATGCAGCTCCCATAG R:GCAGATGATCCCCTGGGTTG | 87 |
| MRF4 | NM_181811 | F:TGGACCCCTTCAGCTACAGA R:ATGCTTGTCCCTCCTTCCTTG | 139 |
| PAX7 | XM_015460690 | F:TGCCCTCAGTGAGTTCGATT R:CGGGTTCTGACTCCACATCT | 180 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-F.; Choi, S.-H.; Li, Q.; Wang, Y.; Sun, B.; Tang, L.; Wang, E.-Z.; Hua, H.; Li, X.-Z. Overexpression of DGAT2 Stimulates Lipid Droplet Formation and Triacylglycerol Accumulation in Bovine Satellite Cells. Animals 2022, 12, 1847. https://doi.org/10.3390/ani12141847
Zhang J-F, Choi S-H, Li Q, Wang Y, Sun B, Tang L, Wang E-Z, Hua H, Li X-Z. Overexpression of DGAT2 Stimulates Lipid Droplet Formation and Triacylglycerol Accumulation in Bovine Satellite Cells. Animals. 2022; 12(14):1847. https://doi.org/10.3390/ani12141847
Chicago/Turabian StyleZhang, Jun-Fang, Seong-Ho Choi, Qiang Li, Ying Wang, Bin Sun, Lin Tang, En-Ze Wang, Huan Hua, and Xiang-Zi Li. 2022. "Overexpression of DGAT2 Stimulates Lipid Droplet Formation and Triacylglycerol Accumulation in Bovine Satellite Cells" Animals 12, no. 14: 1847. https://doi.org/10.3390/ani12141847