
10-Hydroxy-trans-2-decenoic Acid, a New Potential Feed Additive for Broiler Chickens to Improve Growth Performance

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
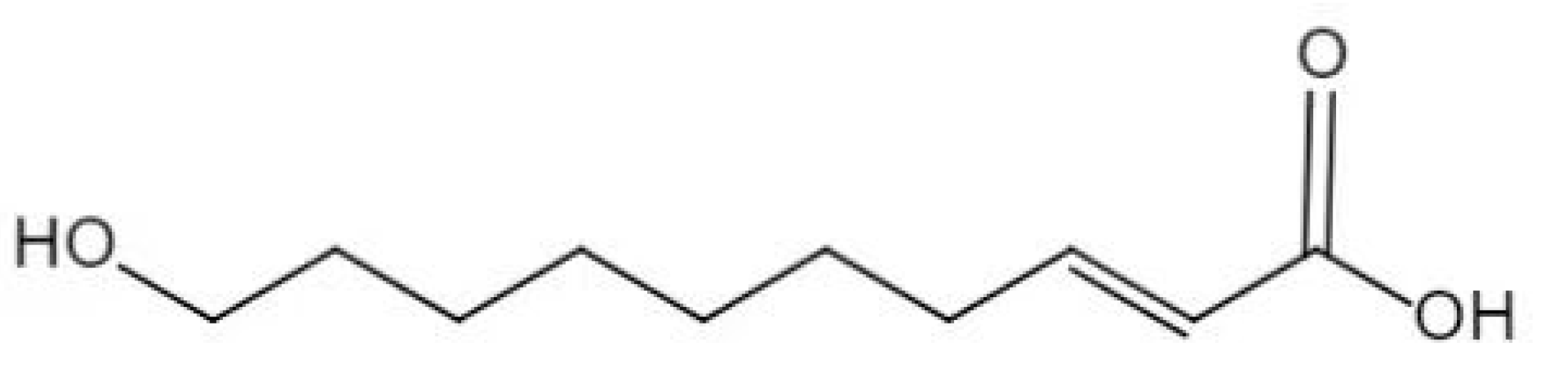
2.1. Experimental Material
2.2. Experimental Design, Diets and Management
2.3. Growth Performance
2.4. Sample Collection and Storage
2.5. Sample Analyses
2.6. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Immunity
3.3. Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butenandt, A.; Rembold, H. Royal jelly of the honeybee. I. Isolation, constitution analysis, and incidence of 10-hydroxy-delta 2-decenoi acid. Hoppe Seylers Z. Physiol. Chem. 1957, 308, 284–289. [Google Scholar] [CrossRef]
- Blum, M.S.; Novak, A.F.; Taber, S. 10-Hydroxy-Δ2-Decenoic Acid, an Antibiotic Found in Royal Jelly. Science 1959, 130, 452–453. [Google Scholar] [CrossRef] [PubMed]
- Šedivá, M.; Klaudiny, J. Antimikrobiálne látky Materskej kašičky. Chem. Listy 2015, 109, 755–761. Available online: http://chemicke-listy.cz/ojs3/index.php/chemicke-listy/article/view/298 (accessed on 6 July 2022).
- Gao, K.; Su, B.; Dai, J.; Li, P.; Wang, R.; Yang, X. Anti-Biofilm and Anti-Hemolysis Activities of 10-Hydroxy-2-decenoic Acid against Staphylococcus aureus. Molecules 2022, 27, 1485. [Google Scholar] [CrossRef]
- Sugiyama, T.; Takahashi, K.; Mori, H. Royal jelly acid, 10-hydroxy-trans-2-decenoic acid, as a modulator of the innate immune responses. Endocr. Metab. Immune Disord. Drug Targets 2012, 12, 368–376. [Google Scholar] [CrossRef]
- Sugiyama, T.; Takahashi, K.; Tokoro, S.; Gotou, T.; Neri, P.; Mori, H. Inhibitory effect of 10-hydroxy-trans-2-decenoic acid on LPS-induced IL-6 production via reducing IκB-ζ expression. Innate Immun. 2012, 18, 429–437. [Google Scholar] [CrossRef]
- Fan, P.; Han, B.; Hu, H.; Wei, Q.; Zhang, X.; Meng, L.; Nie, J.; Tang, X.; Tian, X.; Zhang, L.; et al. Proteome of thymus and spleen reveals that 10-hydroxydec-2-enoic acid could enhance immunity in mice. Expert Opin. Ther. Targets 2020, 24, 267–279. [Google Scholar] [CrossRef]
- Gu, H.; Song, I.B.; Han, H.J.; Lee, N.Y.; Cha, J.Y.; Son, Y.K.; Kwon, J. Antioxidant Activity of Royal Jelly Hydrolysates Obtained by Enzymatic Treatment. Korean J. Food Sci. Anim. Resour. 2018, 38, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Watadani, R.; Kotoh, J.; Sasaki, D.; Someya, A.; Matsumoto, K.; Maeda, A. 10-Hydroxy-2-decenoic acid, a natural product, improves hyperglycemia and insulin resistance in obese/diabetic KK-Ay mice, but does not prevent obesity. J. Vet. Med. Sci. 2017, 79, 1596–1602. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.C.; Sun, H.T.; Lin, I.P.; Kuo, P.C.; Li, J.C. The functional property of royal jelly 10-hydroxy-2-decenoic acid as a melanogenesis inhibitor. BMC Complement. Altern. Med. 2017, 17, 392. [Google Scholar] [CrossRef]
- Li, S.; Jiang, Z.; Thamm, L.; Zhou, G. 10-Hydroxy-2-decenoic Acid as an Antimicrobial Agent in Draft Keg-conditioned Wheat Beer. J. Am. Soc. Brew. Chem. 2010, 68, 114–118. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I. Vitagenes in poultry production: Part 1. Technological and environmental stresses. Worlds Poult. Sci. J. 2016, 72, 721–734. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994; pp. 57–63. [Google Scholar]
- AOAC International. Official Methods of the AOAC International, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Weiser, M.J.; Grimshaw, V.; Wynalda, K.M.; Mohajeri, M.H.; Butt, C.M. Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition. Nutrients 2018, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Roura, E.; Homedes, J.; Klasing, K.C. Prevention of Immunologic Stress Contributes to the Growth-Permitting Ability of Dietary Antibiotics in Chicks. J. Nutr. 1992, 122, 2383–2390. [Google Scholar] [CrossRef]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Lai, J.; Khalique, A.; Li, G.; Pan, K.; Jing, B.; Zeng, D. Probiotic Enhanced Intestinal Immunity in Broilers against Subclinical Necrotic Enteritis. Front. Immunol. 2017, 8, 1592. [Google Scholar] [CrossRef] [Green Version]
- Zanotti, S.; Kumar, A.; Kumar, A. Cytokine modulation in sepsis and septic shock. Expert Opin. Investig. Drugs 2002, 11, 1061–1075. [Google Scholar] [CrossRef]
- Park, E.J.; Lee, J.H.; Yu, G.-Y.; He, G.; Ali, S.R.; Holzer, R.G.; Österreicher, C.H.; Takahashi, H.; Karin, M. Dietary and Genetic Obesity Promote Liver Inflammation and Tumorigenesis by Enhancing IL-6 and TNF Expression. Cell 2010, 140, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; You, M.M.; Liu, Y.C.; Shi, Y.Z.; Wang, K.; Lu, Y.Y.; Hu, F.L. Potential protective effect of trans-10-hydroxy-2-decenoic acid on the inflammation induced by Lipoteichoic acid. J. Funct. Foods 2018, 45, 491–498. [Google Scholar] [CrossRef]
- Huang, S.; Tao, R.; Zhou, J.; Qian, L.; Wu, J. Trans-10-hydroxy-2-decenoic acid alleviates dextran sulfate sodium-induced colitis in mice via regulating the inflammasome-mediated pyroptotic pathway and enhancing colonic barrier function. Mol. Nutr. Food Res. 2022, 2100821. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem.-Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Hadj Ahmed, S.; Koubaa, N.; Kharroubi, W.; Zarrouk, A.; Mnari, A.; Batbout, F.; Gamra, H.; Hammami, S.; Lizard, G.; Hammami, M. Identification of long and very long chain fatty acids, plasmalogen-C16:0 and phytanic acid as new lipid biomarkers in Tunisian coronary artery disease patients. Prostaglandins Other Lipid Mediat. 2017, 131, 49–58. [Google Scholar] [CrossRef]
- Albalawi, A.E.; Althobaiti, N.A.; Alrdahe, S.S.; Alhasani, R.H.; Alaryani, F.S.; BinMowyna, M.N. Anti-Tumor Effects of Queen Bee Acid (10-Hydroxy-2-Decenoic Acid) Alone and in Combination with Cyclophosphamide and Its Cellular Mechanisms against Ehrlich Solid Tumor in Mice. Molecules 2021, 26, 7021. [Google Scholar] [CrossRef]
- Almeer, R.S.; Alarifi, S.; Alkahtani, S.; Ibrahim, S.R.; Ali, D.; Moneim, A. The potential hepatoprotective effect of royal jelly against cadmium chloride-induced hepatotoxicity in mice is mediated by suppression of oxidative stress and upregulation of Nrf2 expression. Biomed. Pharmacother. 2018, 106, 1490–1498. [Google Scholar] [CrossRef]
- Guo, Y.; Yu, S.; Zhang, C.; Kong, A.-N.T. Epigenetic regulation of Keap1-Nrf2 signaling. Free Radical Biol. Med. 2015, 88, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Makino, J.; Ogasawara, R.; Kamiya, T.; Hara, H.; Mitsugi, Y.; Yamaguchi, E.; Itoh, A.; Adachi, T. Royal Jelly Constituents Increase the Expression of Extracellular Superoxide Dismutase through Histone Acetylation in Monocytic THP-1 Cells. J. Nat. Prod. 2016, 79, 1137–1143. [Google Scholar] [CrossRef]
- Wang, B.; Zhu, X.; Kim, Y.; Li, J.; Huang, S.; Saleem, S.; Li, R.-C.; Xu, Y.; Dore, S.; Cao, W. Histone deacetylase inhibition activates transcription factor Nrf2 and protects against cerebral ischemic damage. Free Radical Biol. Med. 2012, 52, 928–936. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radical Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Ingredients | 0~21 d | 22~42 d |
|---|---|---|
| Corn | 60.13 | 61.53 |
| Soybean meal | 32.50 | 31.70 |
| Fish meal | 2.00 | 0.00 |
| Soybean oil | 1.50 | 3.00 |
| Dicalcium phosphate | 1.50 | 1.70 |
| Limestone | 1.34 | 1.15 |
| DL-methionine, 98% | 0.23 | 0.12 |
| Sodium Chloride | 0.30 | 0.30 |
| Premix 1 | 0.50 | 0.50 |
| Total | 100.00 | 100.00 |
| Nutrient Concentrations (%) 1 | 10-Hydroxy-trans-2-decenoic Acid (mg/kg) | |
|---|---|---|
| 0 | 40 | |
| 0~21 d | ||
| Metabolizable energy (MJ/kg) | 12.59 | 12.59 |
| Crude protein | 21.51 | 21.54 |
| Calcium | 1.06 | 1.09 |
| Total phosphorus | 0.69 | 0.70 |
| Lysine | 1.20 | 1.18 |
| Methionine + Cysteine | 0.92 | 0.93 |
| 22~42 d | ||
| Metabolizable energy (MJ/kg) | 13.22 | 13.22 |
| Crude protein | 20.31 | 20.21 |
| Calcium | 0.92 | 0.91 |
| Total phosphorus | 0.66 | 0.67 |
| Lysine | 1.14 | 1.13 |
| Methionine + Cysteine | 0.85 | 0.85 |
| Item 2 | 10-Hydroxy-trans-2-decenoic Acid (mg/kg) | SEM | p Value | |
|---|---|---|---|---|
| 0 | 40 | |||
| BW at d 42 (kg) | 2.24 | 2.35 | 0.028 | 0.014 |
| 0~21 d | ||||
| ADG (g) | 31.5 | 32.2 | 0.293 | 0.111 |
| ADFI (g) | 42.3 | 42.6 | 0.285 | 0.395 |
| F:G | 1.34 | 1.32 | 0.016 | 0.443 |
| 22~42 d | ||||
| ADG (g) | 73.1 | 77.9 | 1.03 | 0.008 |
| ADFI (g) | 131 | 135 | 1.30 | 0.080 |
| F:G | 1.79 | 1.73 | 0.030 | 0.190 |
| 0~42 d | ||||
| ADG (g) | 51.3 | 53.9 | 0.621 | 0.013 |
| ADFI (g) | 84.7 | 86.6 | 0.658 | 0.076 |
| F:G | 1.65 | 1.61 | 0.023 | 0.165 |
| Item 2 | 10-Hydroxy-trans-2-decenoic Acid (mg/kg) | SEM | p Value | |
|---|---|---|---|---|
| 0 | 40 | |||
| 21 d | ||||
| IgG (g/L) | 5.64 | 6.82 | 0.370 | 0.048 |
| IgA (g/L) | 0.915 | 0.883 | 0.041 | 0.589 |
| IgM (g/L) | 0.781 | 0.878 | 0.035 | 0.079 |
| IL-1β (pg/mL) | 43.2 | 39.1 | 2.57 | 0.287 |
| TNF-α (pg/mL) | 87 | 72.6 | 2.80 | 0.005 |
| IL-6 (pg/mL) | 72.4 | 69.2 | 1.71 | 0.206 |
| IL-10 (pg/mL) | 39.9 | 41.6 | 1.77 | 0.521 |
| 42 d | ||||
| IgG (g/L) | 7.13 | 8.09 | 0.439 | 0.151 |
| IgA (g/L) | 1.24 | 1.19 | 0.060 | 0.557 |
| IgM (g/L) | 0.729 | 0.897 | 0.030 | 0.003 |
| IL-1β (pg/mL) | 42 | 28.5 | 1.16 | <0.001 |
| TNF-α (pg/mL) | 83.1 | 65.8 | 2.09 | <0.001 |
| IL-6 (pg/mL) | 75.1 | 62.8 | 1.41 | <0.001 |
| IL-10 (pg/mL) | 35.9 | 43.7 | 1.42 | 0.003 |
| Item 2 | 10-Hydroxy-trans-2-decenoic Acid (mg/kg) | SEM | p Value | |
|---|---|---|---|---|
| 0 | 40 | |||
| 21 d | ||||
| T-SOD (U/mL) | 114 | 129 | 6.66 | 0.164 |
| GSH-PX (U/mL) | 352 | 358 | 9.16 | 0.632 |
| T-AOC (mmol/L) | 0.311 | 0.318 | 0.010 | 0.601 |
| MDA (nmol/mL) | 4.19 | 3.17 | 0.257 | 0.019 |
| 42 d | ||||
| T-SOD (U/mL) | 134 | 153 | 1.62 | < 0.001 |
| GSH-PX (U/mL) | 370 | 392 | 12.1 | 0.222 |
| T-AOC (mmol/L) | 0.323 | 0.318 | 0.008 | 0.649 |
| MDA (nmol/mL) | 3.39 | 2.32 | 0.154 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Geng, S.; Di, Y.; Sun, Y.; Liu, Y.; Li, J.; Zhang, L. 10-Hydroxy-trans-2-decenoic Acid, a New Potential Feed Additive for Broiler Chickens to Improve Growth Performance. Animals 2022, 12, 1846. https://doi.org/10.3390/ani12141846
Zhang Y, Geng S, Di Y, Sun Y, Liu Y, Li J, Zhang L. 10-Hydroxy-trans-2-decenoic Acid, a New Potential Feed Additive for Broiler Chickens to Improve Growth Performance. Animals. 2022; 12(14):1846. https://doi.org/10.3390/ani12141846
Chicago/Turabian StyleZhang, Yuxin, Shixia Geng, Yuting Di, Yongbo Sun, Ying Liu, Juntao Li, and Liying Zhang. 2022. "10-Hydroxy-trans-2-decenoic Acid, a New Potential Feed Additive for Broiler Chickens to Improve Growth Performance" Animals 12, no. 14: 1846. https://doi.org/10.3390/ani12141846