Plasma Carboxyl-Metabolome Is Associated with Average Daily Gain Divergence in Beef Steers



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Feeding, and Growth Performance
2.2. CIL-LC/MS-Based Metabolomics
Metabolite Data Processing and Identification
2.3. Statistical and Data analysis
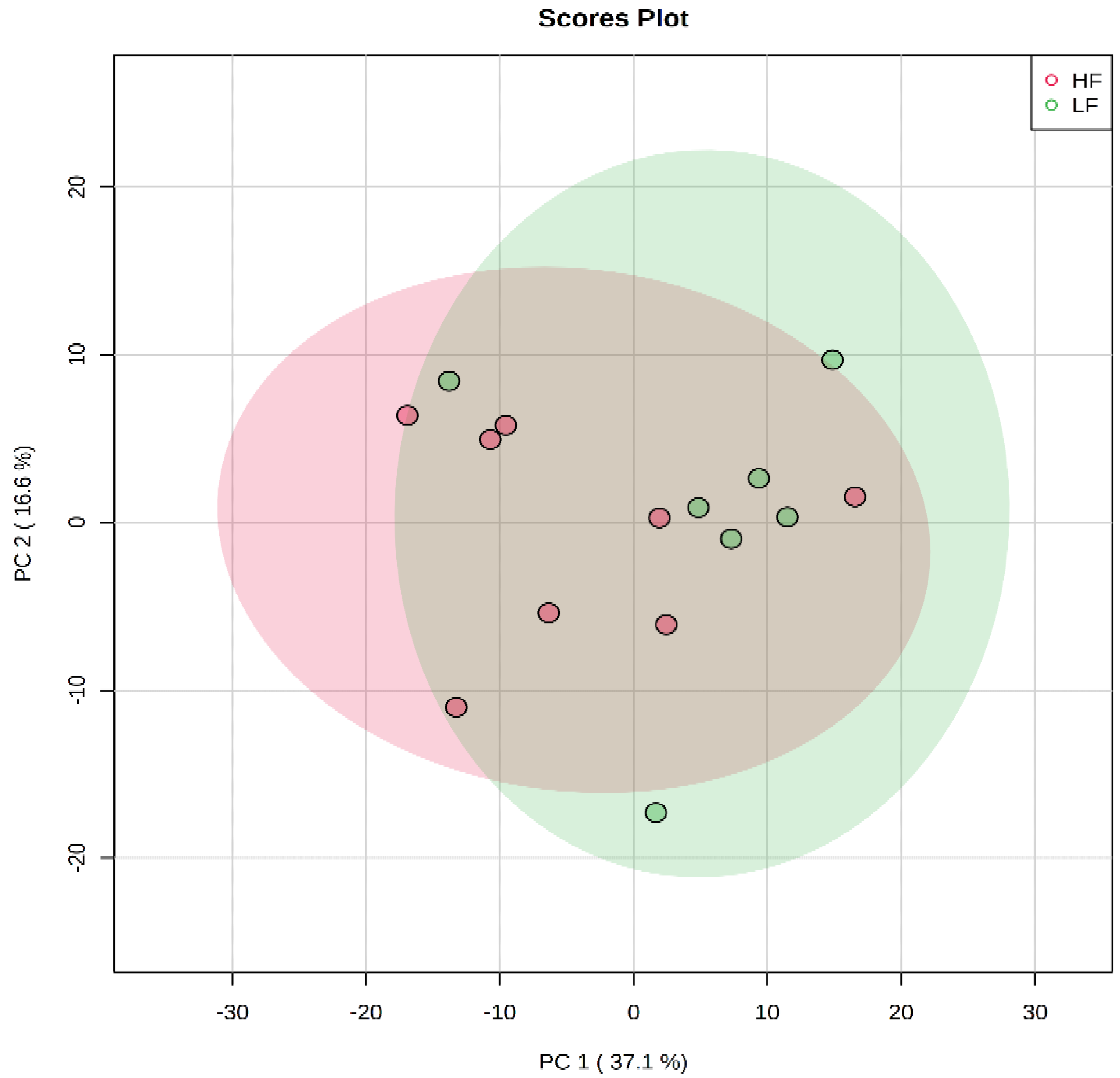
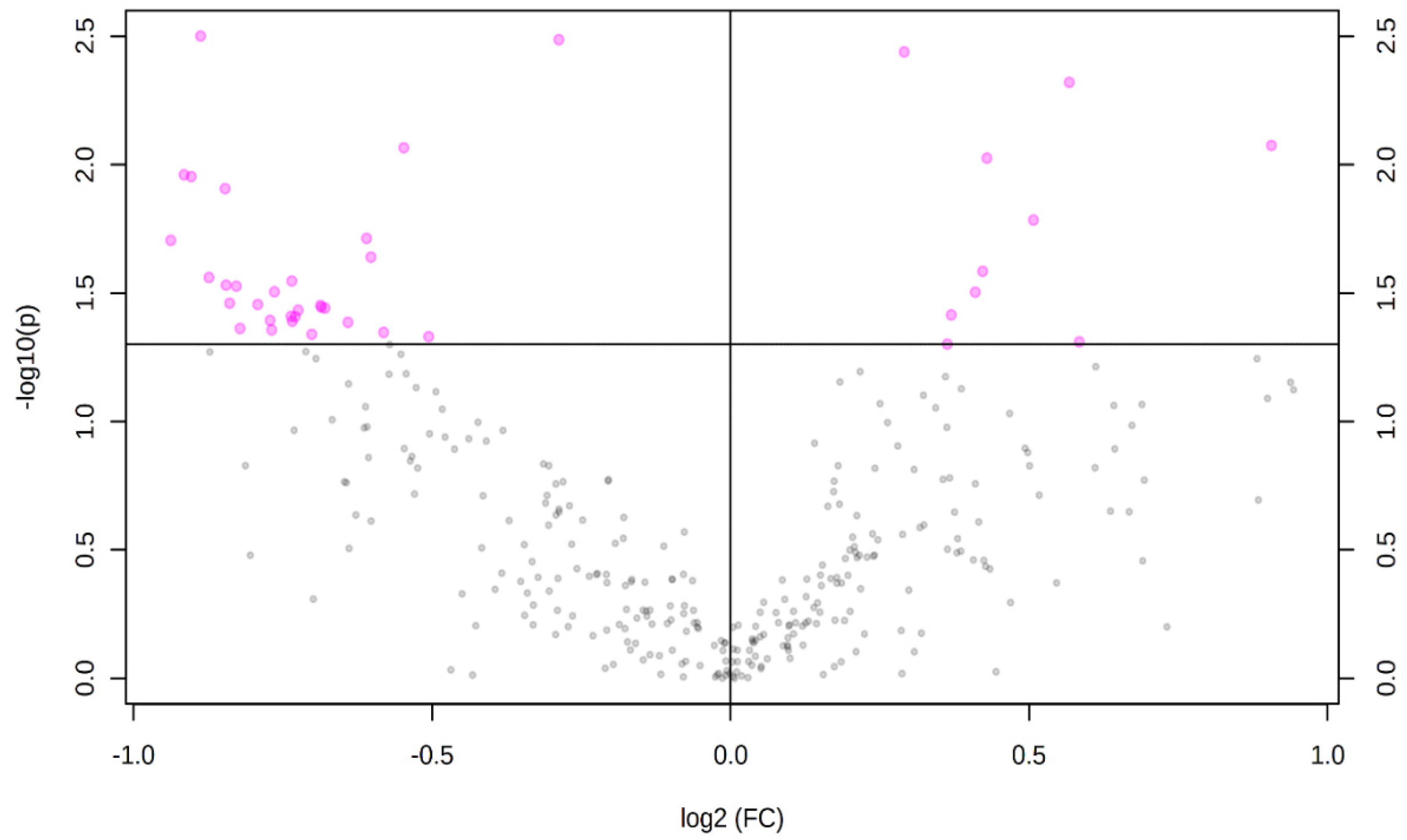
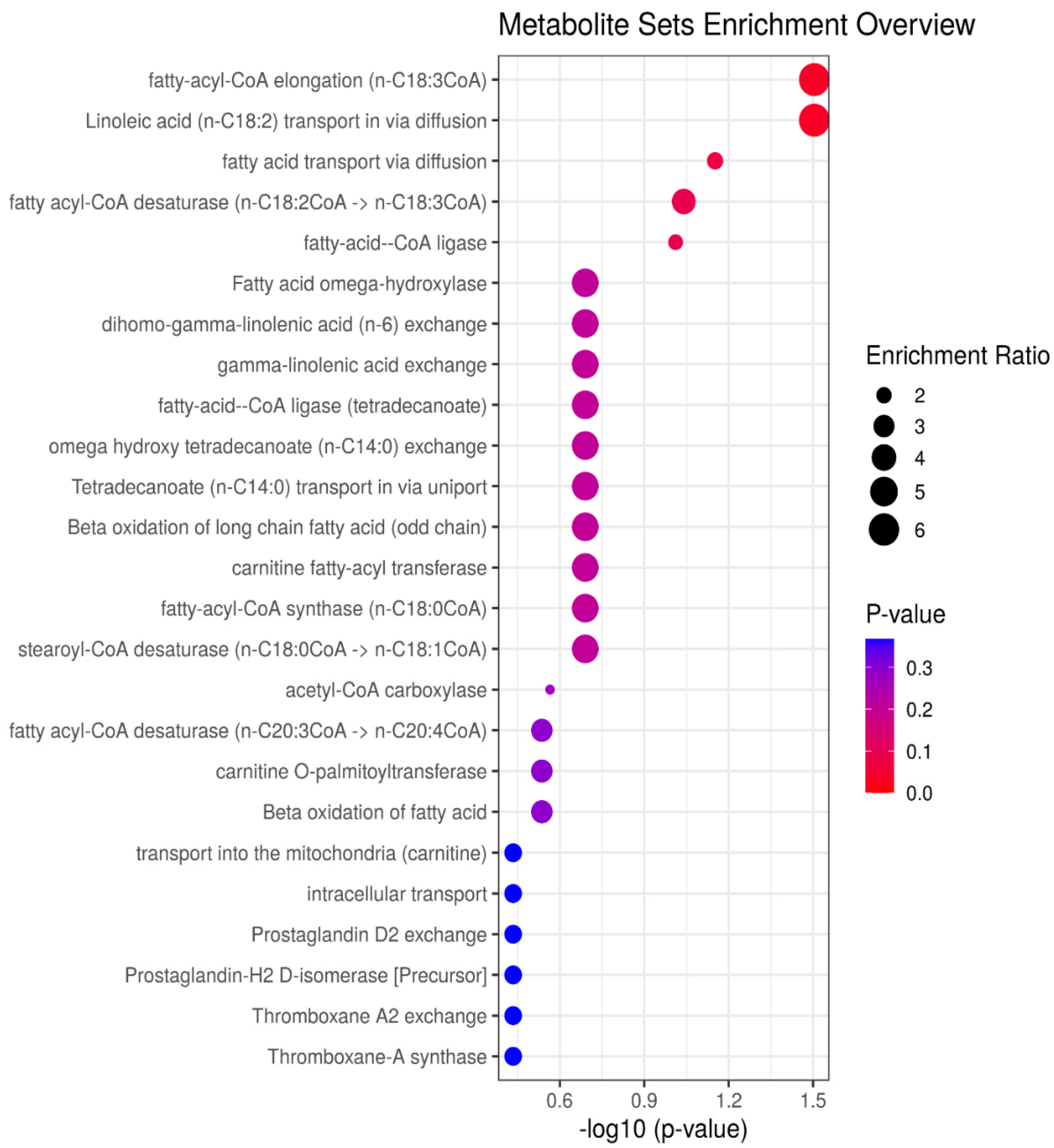
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonano, C.V.; Montanholi, Y.R.; Schenkel, F.S.; Smith, B.A.; Cant, J.P.; Miller, S.P. The relationship between feed efficiency and the circadian profile of blood plasma analytes measured in beef heifers at different physiological stages. Animal 2014, 8, 1684–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, P.A.; Kogelman, L.J.A.; Santana, M.H.A.; Passarelli, D.; Pulz, L.H.; Fantinato-Neto, P.; Silva, P.L.; Leme, P.R.; Strefezzi, R.F.; Coutinho, L.L.; et al. Liver transcriptomic networks reveal main biological processes associated with feed efficiency in beef cattle. BMC Genom. 2015, 16, 1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artegoitia, V.M.; Foote, A.P.; Lewis, R.M.; Freetly, H.C. Rumen Fluid Metabolomics Analysis Associated with Feed Efficiency on Crossbred Steers. Sci. Rep. 2017, 7, 2864. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, L.D.; Eler, J.P.; Pereira, M.A.; Rosa, A.F.; Alexandre, P.A.; Moncau, C.T.; Salvato, F.; Rosa-Fernandes, L.; Palmisano, G.; Ferraz, J.B.S.; et al. Liver proteomics unravel the metabolic pathways related to Feed Efficiency in beef cattle. Sci. Rep. 2019, 9, 5364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The Human Serum Metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, H.; Han, W.; Chan, W.; Li, L. Metabolomic Coverage of Chemical-Group-Submetabolome Analysis: Group Classification and Four-Channel Chemical Isotope Labeling LC-MS. Anal. Chem. 2019, 91, 12108–12115. [Google Scholar] [CrossRef]
- Ogunade, I.M.; McCoun, M. Average daily gain divergence in beef steers is associated with altered plasma metabolome and whole blood immune-related gene expression. Transl. Anim. Sci. 2020, 4, txaa074. [Google Scholar] [CrossRef]
- Mukiibi, R.; Vinsky, M.; Keogh, K.A.; Fitzsimmons, C.; Stothard, P.; Waters, S.M.; Li, C. Transcriptome analyses reveal reduced hepatic lipid synthesis and accumulation in more feed efficient beef cattle. Sci. Rep. 2018, 8, 7303. [Google Scholar] [CrossRef]
- Guo, K.; Li, L. High-Performance Isotope Labeling for Profiling Carboxylic Acid-Containing Metabolites in Biofluids by Mass Spectrometry. Anal. Chem. 2010, 82, 8789–8793. [Google Scholar] [CrossRef]
- Wu, Y.; Li, L. Determination of Total Concentration of Chemically Labeled Metabolites as a Means of Metabolome Sample Normalization and Sample Loading Optimization in Mass Spectrometry-Based Metabolomics. Anal. Chem. 2012, 84, 10723–10731. [Google Scholar] [CrossRef]
- Mung, D.; Li, L. Development of Chemical Isotope Labeling LC-MS for Milk Metabolomics: Comprehensive and Quantitative Profiling of the Amine/Phenol Submetabolome. Anal. Chem. 2017, 89, 4435–4443. [Google Scholar] [CrossRef] [PubMed]
- Huan, T.; Li, L. Quantitative Metabolome Analysis Based on Chromatographic Peak Reconstruction in Chemical Isotope Labeling Liquid Chromatography Mass Spectrometry. Anal. Chem. 2015, 87, 7011–7016. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, R.; Zhou, J.; Zuniga, A.; Stanislaus, A.E.; Wu, Y.; Huan, T.; Zheng, J.; Shi, Y.; Wishart, D.S.; et al. MyCompoundID: Using an Evidence-Based Metabolome Library for Metabolite Identification. Anal. Chem. 2013, 85, 3401–3408. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artegoitia, V.M.; Foote, A.P.; Lewis, R.M.; Freetly, H.C. Metabolomics Profile and Targeted Lipidomics in Multiple Tissues Associated with Feed Efficiency in Beef Steers. ACS Omega 2019, 4, 3973–3982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm-Perry, A.K.; Butler, A.R.; Kern, R.J.; Hill, R.; Kuehn, L.A.; Wells, J.E.; Oliver, W.T.; Hales, K.E.; Foote, A.P.; Freetly, H.C. Differential gene expression in the duodenum, jejunum and ileum among crossbred beef steers with divergent gain and feed intake phenotypes. Anim. Genet. 2016, 47, 408–427. [Google Scholar] [CrossRef]
- Foote, A.P.; Keel, B.N.; Zarek, C.M.; Lindholm-Perry, A.K. Beef steers with average dry matter intake and divergent average daily gain have altered gene expression in the jejunum. J. Anim. Sci. 2017, 95, 4430–4439. [Google Scholar] [CrossRef] [Green Version]
- Spector, A.A. Arachidonic acid cytochrome P450 epoxygenase pathway. J. Lipid Res. 2009, 50, S52–S56. [Google Scholar] [CrossRef] [Green Version]
- Bonet, M.L.; Ribot, J.; Felipe, F.; Palou, A. Vitamin A and the regulation of fat reserves. Cell. Mol. Life Sci. 2003, 60, 1311–1321. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Cunningham, H.C.; Kuehn, L.A.; Vallet, J.L.; Keele, J.W.; Foote, A.P.; Cammack, K.M.; Freetly, H.C. Relationships between the genes expressed in the mesenteric adipose tissue of beef cattle and feed intake and gain. Anim. Genet. 2017, 48, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Hiragun, A.; Mitsui, H. Preadipocytes possess cellular retinoid binding proteins and their differentiation is inhibited by retinoids. Biochem. Biophys. Res. Commun. 1980, 95, 1839–1845. [Google Scholar] [CrossRef]
- Berry, D.C.; DeSantis, D.; Soltanian, H.; Croniger, C.M.; Noy, N. Retinoic Acid Upregulates Preadipocyte Genes to Block Adipogenesis and Suppress Diet-Induced Obesity. Diabetes 2012, 61, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, A.; Maruo, Y.; Miki, T.; Yamasaki, T.; Saito, T. Influence of vitamin A on the quality of beef from the Tajima strain of Japanese Black cattle. Meat Sci. 1998, 48, 159–167. [Google Scholar] [CrossRef]
- Allen, M.S. Drives and limits to feed intake in ruminants. Anim. Prod. Sci. 2014, 54, 1513–1524. [Google Scholar] [CrossRef]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of arachidonic acid metabolism: A review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef]
- Ohmori, K.; Yamada, H.; Yasuda, A.; Yamamoto, A.; Matsuura, N.; Kiniwa, M. Anti-hyperlipidemic action of a newly synthesized benzoic acid derivative, S-2E. Eur. J. Pharmacol. 2003, 471, 69–76. [Google Scholar] [CrossRef]
- Tong, L.; Harwood, H.J., Jr. Acetyl-coenzyme A carboxylases: Versatile targets for drug discovery. J. Cell. Biochem. 2006, 99, 1476–1488. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.W. Malonyl-CoA: The regulator of fatty acid synthesis and oxidation. J. Clin. Investig. 2012, 122, 1958–1959. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty Acid Desaturases, Polyunsaturated Fatty Acid Regulation, and Biotechnological Advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinti, D.L.; Cook, L.; Nagi, M.N.; Suneja, S.K. The fatty acid chain elongation system of mammalian endoplasmic reticulum. Prog. Lipid Res. 1992, 31, 1–51. [Google Scholar] [CrossRef]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Stahl, A. SLC27 fatty acid transport proteins. Mol. Asp. Med. 2013, 34, 516–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londesborough, J.C.; Webster, L.T. 14. Fatty Acyl-CoA Synthetases. In The Enzymes; Boyer, P.D., Ed.; Academic Press: New York, NY, USA, 1974; Volume 10, pp. 469–488. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Metabolite | FC | FDR | Identification Level |
|---|---|---|---|
| 6-amino-2-oxohexanoate | 0.52 | 0.02 | Tier 2 |
| 8-methyl-6-nonenoic acid | 0.53 | 0.01 | Tier 2 |
| Citronellate | 0.53 | 0.01 | Tier 2 |
| 12-oxo-9(Z)-dodecenoic acid | 0.54 | 0.01 | Tier 2 |
| Oleic acid | 0.55 | 0.03 | Tier 1 |
| Isomer 1 of oleic acid | 0.58 | 0.04 | Tier 1 |
| Retinoic acid | 0.56 | 0.01 | Tier 1 |
| Isomer of retinoic acid | 0.57 | 0.04 | Tier 2 |
| Isomer 2 of oleic acid | 0.56 | 0.03 | Tier 1 |
| Linoleate | 0.56 | 0.03 | Tier 1 |
| 9-cis,11-trans-octadecadienoate | 0.56 | 0.03 | Tier 2 |
| Octadecatrienoic acid | 0.58 | 0.04 | Tier 2 |
| Hexadecenoic acid | 0.59 | 0.04 | Tier 2 |
| 9Z-hexadecenoic acid | 0.59 | 0.04 | Tier 2 |
| Pentadecylic acid | 0.59 | 0.03 | Tier 1 |
| Myristic acid | 0.60 | 0.04 | Tier 1 |
| Arachidonic acid | 0.60 | 0.03 | Tier 1 |
| 9,10,18-trihydroxystearate | 0.60 | 0.04 | Tier 2 |
| Isomer of pentadecylic acid | 0.60 | 0.04 | Tier 1 |
| 9,10,18-trihydroxystearate | 0.61 | 0.04 | Tier 2 |
| 9,10-epoxy-18-hydroxystearate | 0.61 | 0.05 | Tier 2 |
| 9Z-hexadecenoic acid | 0.62 | 0.04 | Tier 2 |
| Octadecatrienoic acid | 0.62 | 0.04 | Tier 2 |
| Arachidonic acid | 0.62 | 0.04 | Tier 2 |
| Tetradecanoic acid | 0.64 | 0.04 | Tier 2 |
| 8-methyl-6-nonenoic acid | 0.66 | 0.02 | Tier 2 |
| syn-Stemoden-19-oate | 0.66 | 0.02 | Tier 2 |
| Eicosatrienoic acid | 0.67 | 0.04 | Tier 2 |
| Isomer eicosadienoic acid | 0.67 | 0.04 | Tier 2 |
| 16-oxopalmitate | 0.68 | <0.01 | Tier 2 |
| Isomer of eicosatrienoic acid | 0.69 | 0.04 | Tier 2 |
| 18-oxooleate | 0.71 | 0.05 | Tier 2 |
| 6-hydroxy-5-isopropenyl-2-methylhexanoate | 0.82 | 0.01 | Tier 2 |
| 3-isohydroxybutyrate | 1.22 | <0.01 | Tier 2 |
| Butanoic acid | 1.29 | 0.05 | Tier 2 |
| Hydroxyisovalerate | 1.29 | 0.04 | Tier 2 |
| Leukotriene B4 | 1.33 | 0.03 | Tier 2 |
| Acetate | 1.34 | 0.03 | Tier 1 |
| 3-hydroxybutyrate | 1.35 | <0.01 | Tier 1 |
| Isomer of acetate | 1.42 | 0.02 | Tier 2 |
| 11H-14,15-EETA | 1.48 | 0.01 | Tier 2 |
| 4-ethylbenzoic acid | 1.50 | 0.05 | Tier 1 |
| L-threo-3-methylaspartate | 1.87 | 0.01 | Tier 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogunade, I.; Oyebade, A.; Osa-Andrews, B.; Peters, S. Plasma Carboxyl-Metabolome Is Associated with Average Daily Gain Divergence in Beef Steers. Animals 2021, 11, 67. https://doi.org/10.3390/ani11010067
Ogunade I, Oyebade A, Osa-Andrews B, Peters S. Plasma Carboxyl-Metabolome Is Associated with Average Daily Gain Divergence in Beef Steers. Animals. 2021; 11(1):67. https://doi.org/10.3390/ani11010067
Chicago/Turabian StyleOgunade, Ibukun, Adeoye Oyebade, Bremansu Osa-Andrews, and Sunday Peters. 2021. "Plasma Carboxyl-Metabolome Is Associated with Average Daily Gain Divergence in Beef Steers" Animals 11, no. 1: 67. https://doi.org/10.3390/ani11010067