Investigating the Features of PDO Green Hams during Salting: Insights for New Markers and Genomic Regions in Commercial Hybrid Pigs

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Data Availability
2.2. Carcass and Ham Traits
2.3. Non-Invasive Magnetic Induction (MI) System Analysis
2.4. Statistical Analyses of Ham Traits
2.5. Genotyping and Association Study
3. Results
3.1. Ham Quality and Technological Traits
3.2. Association Study Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carni, I.M. Carne Suina e Salumi. Available online: http://www.ismeamercati.it/carni/carne-suina-salumi (accessed on 22 October 2020).
- Bosi, P.; Russo, V. The production of the heavy pig for high quality processed products. Ital. J. Anim. Sci. 2004, 3, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Candek-Potokar, M.; Skrlep, M. Factors in pig production that impact the quality of dry-cured ham: A review. Anim. Int. J. Anim. Biosci. 2012, 6, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Prosciutto di Parma (Parma Ham) Protected Designation of Origin (Specifications and Dossier Pursuant to Article 4 of Council Regulation EEC no. 2081/92 Dated 14 July 1992). Offic. J. L 1992, 208, 9–14.
- Schivazappa, C.; Virgili, R. Impact of salt levels on the sensory profile and consumer acceptance of Italian dry-cured ham. J. Sci. Food Agric. 2020, 100, 3370–3377. [Google Scholar] [CrossRef] [PubMed]
- Pinna, A.; Saccani, G.; Schivazappa, C.; Simoncini, N.; Virgili, R. Revision of the cold processing phases to obtain a targeted salt reduction in typical Italian dry-cured ham. Meat Sci. 2020, 161, 107994. [Google Scholar] [CrossRef]
- Fernández, A.; de Pedro, E.; Núñez, N.; Silió, L.; García-Casco, J.; Rodríguez, C. Genetic parameters for meat and fat quality and carcass composition traits in Iberian pigs. Meat Sci. 2003, 64, 405–410. [Google Scholar] [CrossRef]
- Davoli, R.; Catillo, G.; Serra, A.; Zappaterra, M.; Zambonelli, P.; Zilio, D.M.; Steri, R.; Mele, M.; Buttazzoni, L.; Russo, V. Genetic parameters of backfat fatty acids and carcass traits in Large White pigs. Anim. Int. J. Anim. Biosci. 2019, 13, 924–932. [Google Scholar] [CrossRef]
- Davoli, R.; Luise, D.; Mingazzini, V.; Zambonelli, P.; Braglia, S.; Serra, A.; Russo, V. Genome-wide study on intramuscular fat in Italian Large White pig breed using the PorcineSNP60 BeadChip. J. Anim. Breed. Genet. 2016, 133, 277–282. [Google Scholar] [CrossRef]
- Davoli, R.; Schivazappa, C.; Zambonelli, P.; Braglia, S.; Rossi, A.; Virgili, R. Association study between single nucleotide polymorphisms in porcine genes and pork quality traits for fresh consumption and processing into Italian dry-cured ham. Meat Sci. 2017, 126, 73–81. [Google Scholar] [CrossRef]
- Toldrá, F.; Flores, M.; Sanz, Y. Dry-cured ham flavour: Enzymatic generation and process influence. Food Chem. 1997, 59, 523–530. [Google Scholar] [CrossRef]
- Davoli, R.; Zappaterra, M.; Zambonelli, P. Genome-wide association study identifies markers associated with meat ultimate pH in Duroc pigs. Anim. Genet. 2019, 50, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Zappaterra, M.; Sami, D.; Davoli, R. Association between the splice mutation g.8283C>A of the PHKG1 gene and meat quality traits in Large White pigs. Meat Sci. 2019, 148, 38–40. [Google Scholar] [CrossRef]
- Renaville, B.; Piasentier, E.; Fan, B.; Vitale, M.; Prandi, A.; Rothschild, M.F. Candidate gene markers involved in San Daniele ham quality. Meat Sci. 2010, 85, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Schiavo, G.; Gallo, M.; Baiocco, C.; Galimberti, G.; Bovo, S.; Russo, V.; Buttazzoni, L. Genome-wide association study for ham weight loss at first salting in Italian Large White pigs: Towards the genetic dissection of a key trait for dry-cured ham production. Anim. Genet. 2017, 48, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulladosa, E.; Muñoz, I.; Serra, X.; Arnau, J.; Gou, P. X-ray absorptiometry for non-destructive monitoring of the salt uptake in bone-in raw hams during salting. Food Control 2015, 47, 37–42. [Google Scholar] [CrossRef]
- Simoncini, N.; Virgili, R.; Schivazappa, C.; Pinna, A.; Rossi, A.; Álvarez, J. Assessment of fat and lean content in Italian heavy green hams by means of on-line non-invasive techniques. In Proceedings of the 58th International Congress of Meat Science and Technology, Montreal, QC, Canada, 12–17 August 2012; p. 4. [Google Scholar]
- Schivazappa, C.; Virgili, R.; Simoncini, N.; Tiso, S.; Álvarez, J.; Rodríguez, J.M. Application of the magnetic induction technique for the non-destructive assessment of salt gain after the salting process of Parma ham. Food Control 2017, 80, 92–98. [Google Scholar] [CrossRef]
- Otto, G.; Roehe, R.; Looft, H.; Thoelking, L.; Knap, P.W.; Rothschild, M.F.; Plastow, G.S.; Kalm, E. Associations of DNA markers with meat quality traits in pigs with emphasis on drip loss. Meat Sci. 2007, 75, 185–195. [Google Scholar] [CrossRef]
- EUR-Lex—32009R1099—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2009/1099/oj (accessed on 28 October 2020).
- EUR-Lex—32005R0001—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32005R0001 (accessed on 28 October 2020).
- 2014/38/EU: Commission Implementing Decision of 24 January 2014 Authorising Methods for Grading Pig Carcases in Italy (Notified under Document C(2014) 279). Offic. J. Eur. Union 2014, 23, 35–40.
- Commission Implementing Regulation (EU) No 1208/2013 of 25 November 2013 Approving Minor Amendments to the Specification for a Name Entered in the Register of Protected Designations of Origin and Protected Geographical Indications (Prosciutto di Parma (PDO)); Publications Office of the European Union: Luxembourg, 2013; Volume 317.
- Council Regulation (EEC) No 2081/92 of 14 July 1992 on the Protection of Geographical Indications and Designations of Origin for Agricultural Products and Foodstuffs. Offic. J. L 1992, 208, 1–9.
- Parolari, G. Review: Achievements, needs and perspectives in dry-cured ham technology: The example of Parma ham/ Revisión: Avances, necesidades y perspectivas de la tecnología del jamón curado: El ejemplo del jamón de Parma. Food Sci. Technol. Int. 1996, 2, 69–78. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; van Duijn, C.M. GenABEL: An R library for genome-wide association analysis. Bioinform. Oxf. Engl. 2007, 23, 1294–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, G.H. Mendelian proportions in a mixed population. Science 1908, 28, 49–50. [Google Scholar] [CrossRef] [PubMed]
- Nicolazzi, E.L.; Biffani, S.; Biscarini, F.; Wengel, P.O.t.; Caprera, A.; Nazzicari, N.; Stella, A. Software solutions for the livestock genomics SNP array revolution. Anim. Genet. 2015, 46, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Burton, P.R.; Clayton, D.G.; Cardon, L.R.; Craddock, N.; Deloukas, P.; Duncanson, A.; Kwiatkowski, D.P.; McCarthy, M.I.; Ouwehand, W.H.; Samani, N.J.; et al. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 2007, 447, 661–678. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. {nlme}: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-150. 2020. Available online: https://cran.r-project.org/web/packages/nlme/nlme.pdf (accessed on 28 October 2020).
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; SAGE Publications, Inc: Thousand Oaks, CA, USA, 2011; ISBN 978-1-4129-7514-8. [Google Scholar]
- Baker, M. Quantitative data: Learning to share. Nat. Methods 2012, 9, 39–41. [Google Scholar] [CrossRef]
- Gupta, S.; Stamatoyannopoulos, J.A.; Bailey, T.L.; Noble, W.S. Quantifying similarity between motifs. Genome Biol. 2007, 8, R24. [Google Scholar] [CrossRef] [Green Version]
- Istituto Parma Qualità. Istituto Nord Est Qualità Impiego delle Nuove Equazioni e Criteri di Accettazione delle Carcasse Suine- Circolare Unificata N. 11/2014; 2014. Available online: http://www.parmaqualita.it/IPQ/datiute/6/112014CRITERIACCETTABILITACARCASSE.pdf (accessed on 28 October 2020).
- Harkouss, R.; Chevarin, C.; Daudin, J.-D.; Sicard, J.; Mirade, P.-S. Development of a multi-physical finite element-based model that predicts water and salt transfers, proteolysis and water activity during the salting and post-salting stages of the dry-cured ham process. J. Food Eng. 2018, 218, 69–79. [Google Scholar] [CrossRef]
- Bordbar, F.; Jensen, J.; Zhu, B.; Wang, Z.; Xu, L.; Chang, T.; Xu, L.; Du, M.; Zhang, L.; Gao, H.; et al. Identification of muscle-specific candidate genes in Simmental beef cattle using imputed next generation sequencing. PLoS ONE 2019, 14, e0223671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, S.; Luo, Y.; Ma, Y.; Deng, X.; He, Y.; Gao, N.; Zhang, H.; Li, J.; Chen, Z.; Zhang, Z. Genome-wide detection of selective signatures in a Duroc pig population. J. Integr. Agric. 2018, 17, 2528–2535. [Google Scholar] [CrossRef] [Green Version]
- Rooney, C.; White, G.; Nazgiewicz, A.; Woodcock, S.A.; Anderson, K.I.; Ballestrem, C.; Malliri, A. The Rac activator STEF (Tiam2) regulates cell migration by microtubule-mediated focal adhesion disassembly. EMBO Rep. 2010, 11, 292–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maartens, A.P.; Brown, N.H. The many faces of cell adhesion during Drosophila muscle development. Dev. Biol. 2015, 401, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ding, L.; Yang, W.; Wang, J.; Chen, L.; Chang, Y.; Geng, B.; Cui, Q.; Guan, Y.; Yang, J. Hepatic Activation of the FAM3C-HSF1-CaM Pathway Attenuates Hyperglycemia of Obese Diabetic Mice. Diabetes 2017, 66, 1185–1197. [Google Scholar] [CrossRef] [Green Version]
- Hamill, R.M.; McBryan, J.; McGee, C.; Mullen, A.M.; Sweeney, T.; Talbot, A.; Cairns, M.T.; Davey, G.C. Functional analysis of muscle gene expression profiles associated with tenderness and intramuscular fat content in pork. Meat Sci. 2012, 92, 440–450. [Google Scholar] [CrossRef]
- Zhong, J.; Baquiran, J.B.; Bonakdar, N.; Lees, J.; Ching, Y.W.; Pugacheva, E.; Fabry, B.; O’Neill, G.M. NEDD9 Stabilizes Focal Adhesions, Increases Binding to the Extra-Cellular Matrix and Differentially Effects 2D versus 3D Cell Migration. PLoS ONE 2012, 7, e35058. [Google Scholar] [CrossRef] [Green Version]
- Semelakova, M.; Grauzam, S.; Betadthunga, P.; Tiedeken, J.; Coaxum, S.; Neskey, D.M.; Rosenzweig, S.A. Vimentin and Non-Muscle Myosin IIA are Members of the Neural Precursor Cell Expressed Developmentally Down-Regulated 9 (NEDD9) Interactome in Head and Neck Squamous Cell Carcinoma Cells. Transl. Oncol. 2018, 12, 49–61. [Google Scholar] [CrossRef]
- Vogel, T.; Ahrens, S.; Büttner, N.; Krieglstein, K. Transforming Growth Factor β Promotes Neuronal Cell Fate of Mouse Cortical and Hippocampal Progenitors In Vitro and In Vivo: Identification of Nedd9 as an Essential Signaling Component. Cereb. Cortex 2010, 20, 661–671. [Google Scholar] [CrossRef] [Green Version]
- Omata, Y.; Nakamura, S.; Koyama, T.; Yasui, T.; Hirose, J.; Izawa, N.; Matsumoto, T.; Imai, Y.; Seo, S.; Kurokawa, M.; et al. Identification of Nedd9 as a TGF-β-Smad2/3 Target Gene Involved in RANKL-Induced Osteoclastogenesis by Comprehensive Analysis. PLoS ONE 2016, 11, e0157992. [Google Scholar] [CrossRef] [PubMed]
- Ignotz, R.A.; Massagué, J. Type beta transforming growth factor controls the adipogenic differentiation of 3T3 fibroblasts. Proc. Natl. Acad. Sci. USA 1985, 82, 8530–8534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebbapragada, A.; Benchabane, H.; Wrana, J.L.; Celeste, A.J.; Attisano, L. Myostatin Signals through a Transforming Growth Factor β-Like Signaling Pathway To Block Adipogenesis. Mol. Cell. Biol. 2003, 23, 7230–7242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangs, F.K.; Schrode, N.; Hadjantonakis, A.-K.; Anderson, K.V. Lineage specificity of primary cilia in the mouse embryo. Nat. Cell Biol. 2015, 17, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrighi, N.; Lypovetska, K.; Moratal, C.; Giorgetti-Peraldi, S.; Dechesne, C.A.; Dani, C.; Peraldi, P. The primary cilium is necessary for the differentiation and the maintenance of human adipose progenitors into myofibroblasts. Sci. Rep. 2017, 7, 15248. [Google Scholar] [CrossRef] [PubMed]
- Zappaterra, M.; Gioiosa, S.; Chillemi, G.; Zambonelli, P.; Davoli, R. Muscle transcriptome analysis identifies genes involved in ciliogenesis and the molecular cascade associated with intramuscular fat content in Large White heavy pigs. PLoS ONE 2020, 15, e0233372. [Google Scholar] [CrossRef]
- Shagisultanova, E.; Gaponova, A.V.; Gabbasov, R.; Nicolas, E.; Golemis, E.A. Preclinical and clinical studies of the NEDD9 scaffold protein in cancer and other diseases. Gene 2015, 567, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Huang, Z. FoxO1: A novel insight into its molecular mechanisms in the regulation of skeletal muscle differentiation and fiber type specification. Oncotarget 2016, 8, 10662–10674. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, S.; Jemaa, S.B.; Ciani, E.; Sottile, G.; Moscarelli, A.; Boussaha, M.; Montedoro, M.; Pilla, F.; Cassandro, M. Genome-wide detection of signatures of selection in three Valdostana cattle populations. J. Anim. Breed. Genet. 2020, 137, 609–621. [Google Scholar] [CrossRef]
- rs343048625 (SNP)—Explore This Variant—Sus_scrofa—Ensembl Genome Browser 101. Available online: http://uswest.ensembl.org/Sus_scrofa/Variation/Explore?v=rs343048625;vdb=variation (accessed on 12 October 2020).
- England, S.J.; Cerda, G.A.; Kowalchuk, A.; Sorice, T.; Grieb, G.; Lewis, K.E. Hmx3a does not require its homeodomain for its essential functions in spinal cord, ear and lateral line development. bioRxiv 2020, 68. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, J.; Li, S.; Fairall, L.; Pfisterer, S.G.; Gurnett, J.E.; Xiao, X.; Weston, T.A.; Vashi, D.; Ferrari, A.; Orozco, J.L.; et al. Aster Proteins Facilitate Nonvesicular Plasma Membrane to ER Cholesterol Transport in Mammalian Cells. Cell 2018, 175, 514–529.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutyavin, V.I.; Chawla, A. Aster: A New Star in Cholesterol Trafficking. Cell 2018, 175, 307–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farber, C.; Aten, J.; Farber, E.; Vera, V.; Gularte-Mérida, R.; Islas-Trejo, A.; Wen, P.; Horvath, S.; Lucero, M.; Lusis, A.; et al. Genetic dissection of a major mouse obesity QTL (Carfhg2): Integration of gene expression and causality modeling. Physiol. Genomics 2009, 37, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Serão, N.V.; González-Peña, D.; Beever, J.E.; Faulkner, D.B.; Southey, B.R.; Rodriguez-Zas, S.L. Single nucleotide polymorphisms and haplotypes associated with feed efficiency in beef cattle. BMC Genet. 2013, 14, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]


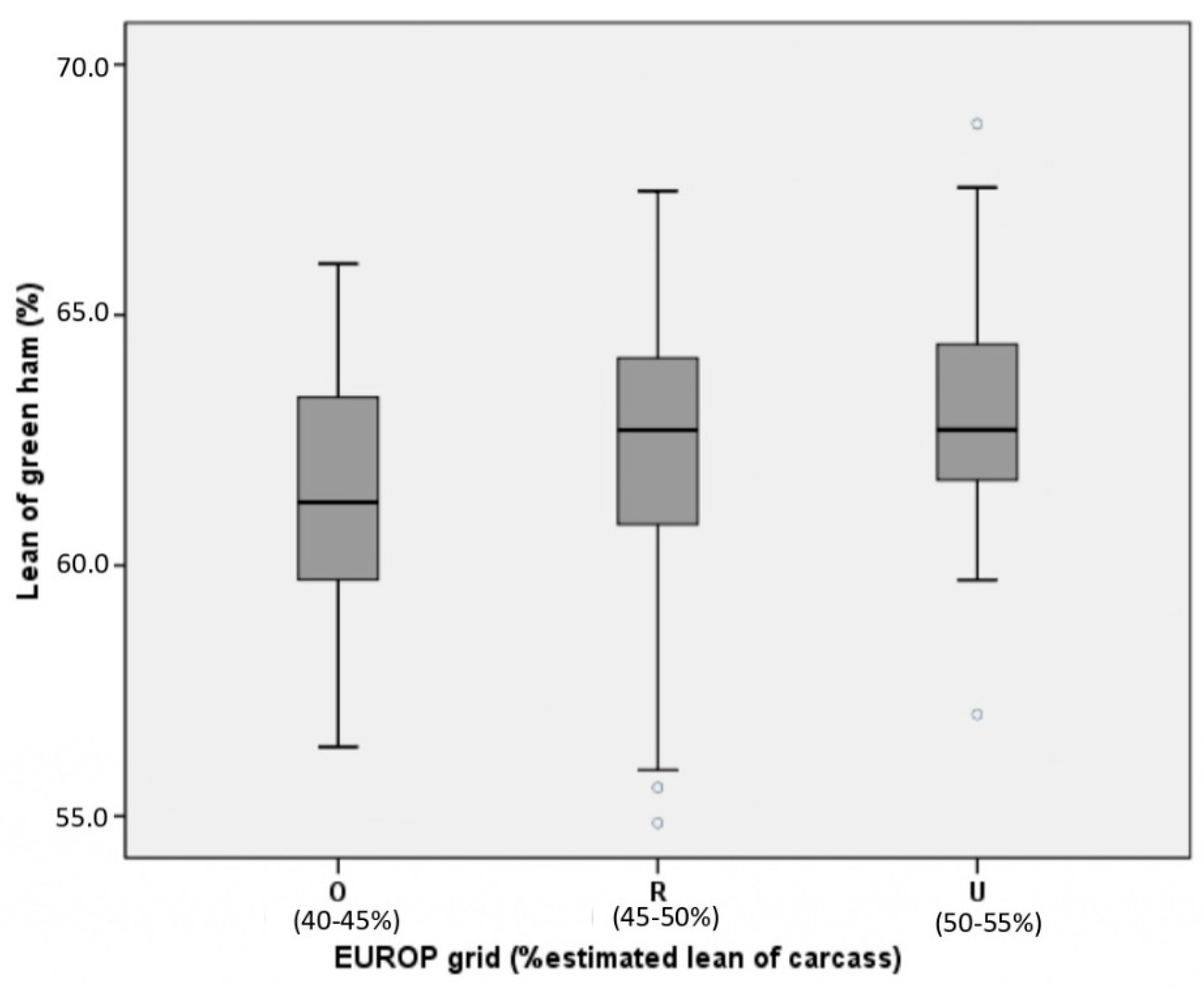{kind=link}
| Ham Traits | Processing Plant (PP) | Sex (S) | PPxS | |||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | p-Value | Barrow | Female | p-Value | ||
| n. 60 | n.120 | n.50 | n. 105 | n. 125 | p-Value | |||
| pHu | 5.63c ± 0.02 | 5.69 b ± 0.01 | 5.79 a ± 0.02 | <0.001 | 5.71 ± 0.01 | 5.70 ± 0.01 | n.s. | n.s. |
| Weight GH, kg 1 | 15.10a ± 0.10 | 13.90 b ± 0.10 | 12.60 c ± 0.20 | <0.001 | 13.90 ± 0.11 | 13.80 ± 0.10 | n.s. | n.s. |
| Lean GH, % 2 | 62.70a ± 0.30 | 62.90 a ± 0.20 | 60.60 b ± 0.30 | <0.001 | 61.10 ± 0.20 | 62.90 ± 0.20 | <0.001 | n.s. |
| Weight 1S, kg 3 | 14.90a ± 0.10 | 13.70 b ± 0.10 | 12.50 c ± 0.20 | <0.001 | 13.70 ± 0.10 | 13.60 ± 0.10 | n.s. | n.s. |
| Weight Loss 1S, % 4 | 1.31 ± 0.03 | 1.26 ± 0.02 | 1.36 ± 0.04 | n.s. | 1.28 ± 0.03 | 1.35 ± 0.02 | <0.05 | n.s. |
| Salt 1S, % 5 | 1.06 c ± 0.01 | 1.21 b ± 0.01 | 1.46 a ± 0.02 | <0.001 | 1.22 ± 0.01 | 1.27 ± 0.01 | <0.001 | n.s. |
| Weight ES, kg 6 | 14.60 a ± 0.10 | 13.50 b ± 0.10 | 12.20 c ± 0.20 | <0.001 | 13.70 ± 0.10 | 13.40 ± 0.10 | n.s. | n.s. |
| Weight Loss ES, % 7 | 3.13 a ± 0.06 | 2.64 b ± 0.04 | 2.72 b ± 0.07 | <0.001 | 2.73 ± 0.05 | 2.93 ± 0.04 | <0.01 | n.s. |
| Salt ES, % 8 | 2.37 c ± 0.02 | 2.80 a ± 0.02 | 2.52 b ± 0.03 | <0.001 | 2.53 ± 0.02 | 2.60 ± 0.02 | <0.01 | <0.05 |
| Salt 1S /Salt ES, % 9 | 44.70 b ± 0.30 | 43.00 c ± 0.20 | 57.50 a ± 0.40 | <0.001 | 48.50 ± 0.30 | 48.30 ± 0.20 | n.s. | n.s. |
| Trait | Marker | Marker Rs Code | Position 1 | MAF 2 | Pc1df 3 | Type of Variant | Candidate Genes in the Region 4 |
|---|---|---|---|---|---|---|---|
| pHu | WU_10.2_18_17949287 | rs321317414 | 18:17,103,785 | 0.22 | 2.54 × 10−6 | intergenic variant | PLXNA4, MKLN1 |
| WU_10.2_4_91195648 | rs342976952 | 4:83,519,869 | 0.13 | 3.94 × 10−5 | intron variant of the gene CD247 | DCAF6, MPC2, ADCY10, MPZL1, RCSD1, CREG1, CD247, POU2F1, DUSP27, GPA33, MAEL | |
| Lean GH, % 5 | ASGA0016987 | rs80994554 | 4:2,020,990 | 0.25 | 1.42 × 10−5 | exon variant of a non-coding transcript | ADGRB1, MROH5, PTP4A3, GPR20, SLC45A4 |
| ALGA0002237 | rs80883186 | 1:30,193,313 | 0.16 | 3.07 × 10−5 | intergenic variant | SGK1, SLC2A12, TBPL1, TCF21, EYA4 | |
| H3GA0000815 | rs80848905 | 1:11,662,820 | 0.35 | 4.72 × 10−5 | intron variant of the gene TIAM2 | NOX3, TFB1M, TIAM2, SCAF8 | |
| ASGA0001055 | rs80923830 | 1:11,684,450 | 0.33 | 4.72 × 10−5 | intron variant of the gene TIAM2 | NOX3, TFB1M, TIAM2, SCAF8 | |
| Weight GH, kg 6 | CASI0010463 | rs335635913 | 15:12,997,362 | 0.13 | 5.48 × 10−6 | intron variant of the gene NXPH2 | NXPH2, SPOPL |
| WU_10.2_14_144250775 | rs343048625 | 14:132,664,262 | 0.40 | 1.33 × 10−5 | intergenic variant | AWN, PSP-II, SPMI, PSTK, IKZF5, ACADSB, HMX2, HMX3, BUB3 | |
| Weight 1S, kg 7 | CASI0010463 | rs335635913 | 15:12,997,362 | 0.13 | 4.13 × 10−6 | intron variant of the gene NXPH2 | NXPH2, SPOPL |
| WU_10.2_14_144250775 | rs343048625 | 14:132,664,262 | 0.40 | 1.26 × 10−5 | intergenic variant | AWN, PSP-II, SPMI, PSTK, IKZF5, ACADSB, HMX2, HMX3, BUB3 | |
| ASGA0026341 | rs80859829 | 5: 74,704,457 | 0.33 | 4.57 × 10−5 | intergenic variant | ADAMTS20, PUS7L, IRAK4, TWF1, U6, TMEM117 | |
| Weight ES, kg 8 | CASI0010463 | rs335635913 | 15:12,997,362 | 0.13 | 2.15 × 10−6 | intron variant of the gene NXPH2 | NXPH2, SPOPL |
| WU_10.2_14_144250775 | rs343048625 | 14:132,664,262 | 0.40 | 4.27 × 10−6 | intergenic variant | AWN, PSP-II, SPMI, PSTK, IKZF5, ACADSB, HMX2, HMX3, BUB3 | |
| ASGA0026341 | rs80859829 | 5: 74,704,457 | 0.33 | 2.23 × 10−5 | intergenic variant | ADAMTS20, PUS7L, IRAK4, TWF1, U6, TMEM117 | |
| WU_10.2_7_118557013 | rs325887861 | 7:111,991,773 | 0.12 | 4.25 × 10−5 | intergenic variant | EFCAB11, TDP1, KCNK13, PSMC1, NRDE2, CALM1 | |
| ALGA0044906 | rs80886909 | 7:112,364,405 | 0.08 | 4.66 × 10−5 | intron variant of the gene TTC7B | PSMC1, NRDE2, CALM1, TTC7B, RPS6K, A5TTC7B | |
| Weight Loss 1S, % 9 | ASGA0031014 | rs80963318 | 7:8,081,734 | 0.47 | 1.00 × 10−5 | intron variant of the gene NEDD9 | GCM2, ELOVL2, SMIM13, NEDD9, TMEM170B, ADTRP, HIVEP1 |
| WU_10.2_10_74620421 | - | 10:67,919,711 | 0.25 | 1.57 × 10−5 | sequence variant of a non-coding gene | ADARB2 | |
| Weight Loss ES, % 10 | ALGA0022599 | rs80917191 | 4:6,665,856 | 0.27 | 2.90 × 10−5 | intergenic variant | KHDRBS3 |
| Salt 1S, % 11 | WU_10.2_14_36295226 | rs323879154 | 14:34,218,184 | 0.44 | 1.71 × 10−5 | intron variant of the genes SUDS3 and SRRM4 | SUDS3, TAOK3, VSIG10, WSB2, RFC5, KSR2SUDS3, SRRM4 |
| ASGA0000817 | rs80921216 | 1:8,287,008 | 0.42 | 2.22 × 10−5 | intergenic variant | FNDC1, TAGAP, RSPH3, EZR, SYTL3, DYNLT1, TMEM181 | |
| Salt ES, % 12 | INRA0000796 | rs332490862 | 1:13,370,639 | 0.08 | 4.36 × 10−6 | intron variant of the gene RGS17 | RGS17,MTRFL1, FBXO5, VIP, MYCT1 |
| ASGA0102337 | rs81323631 | 9:50,457,899 | 0.35 | 1.46 × 10−5 | intron variant of the gene GRAMD1B | HSPA8, CLMP, GRAMD1B, SCN3B, ZNF202, OR6X1, OR6M1, OR4D5, OR6T1, |
| Trait | Marker | Allele | LSM ± S.E (N) | Additive Effect | Dominance Effect | |||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 11 | 12 | 22 | ||||
| pHu | WU_10.2_18_17949287 | G | A | 5.71 a ± 0.04 | 5.73 a ± 0.02 | 5.70 a ± 0.01 | n.s. | n.s. |
| (10) | (55) | (104) | ||||||
| WU_10.2_4_91195648 | G | A | - | 5.69 a ± 0.02 | 5.72 a ± 0.01 | - | - | |
| (0) | (43) | (126) | ||||||
| Lean GH, % 1 | ASGA0016987 | G | A | 63.91 a ± 0.81 | 61.91 b ± 0.27 | 62.61 ab ± 0.23 | n.s. | 0.042 |
| (8) | (64) | (93) | ||||||
| ALGA0002237 | C | A | 62.15 a ± 0.21 | 63.04 a ± 0.33 | 62.47 a ± 1.10 | n.s. | n.s. | |
| (115) | (46) | (4) | ||||||
| H3GA0000815 | G | A | 63.57 a ± 0.48 | 62.71 a ± 0.25 | 61.75 b ± 0.26 | 0.0005 | n.s. | |
| (20) | (75) | (70) | ||||||
| ASGA0001055 | G | A | 61.72 a ± 0.51 | 62.61 a ± 0.26 | 62.40 a ± 0.25 | n.s. | n.s. | |
| (19) | (70) | (76) | ||||||
| Weight GH, kg 2 | CASI0010463 | C | A | 13.58 ab ± 0.68 | 14.05 a ± 0.16 | 13.66 b ± 0.09 | n.s. | n.s. |
| (2) | (38) | (125) | ||||||
| WU_10.2_14_144250775 | G | A | 13.93 ab ± 0.20 | 13.88 a ± 0.10 | 13.48 b ± 0.13 | n.s. | n.s. | |
| (24) | (85) | (56) | ||||||
| Weight 1S, kg 3 | CASI0010463 | C | A | 13.42 ab ± 0.69 | 13.85 a ± 0.16 | 13.48 b ± 0.09 | n.s. | n.s. |
| (2) | (37) | (123) | ||||||
| WU_10.2_14_144250775 | G | A | 13.74 ab ± 0.20 | 13.69 a ± 0.11 | 13.29 b ± 0.13 | n.s. | n.s. | |
| (24) | (83) | (55) | ||||||
| ASGA0026341 | G | A | 14.01 a ± 0.24 | 13.71 a ± 0.11 | 13.29 b ± 0.12 | 0.009 | n.s. | |
| (16) | (78) | (68) | ||||||
| Weight ES, kg 4 | CASI0010463 | C | A | 13.12 ab ± 0.69 | 13.63 a ± 0.17 | 13.24 b ± 0.09 | n.s. | n.s. |
| (2) | (37) | (116) | ||||||
| WU_10.2_14_144250775 | G | A | 13.48 ab ± 0.21 | 13.48 a ± 0.11 | 13.04 b ± 0.13 | n.s. | n.s. | |
| (23) | (79) | (52) | ||||||
| ASGA0026341 | G | A | 13.70 a ± 0.25 | 13.50 a ± 0.11 | 13.05 b ± 0.12 | 0.022 | n.s. | |
| (16) | (78) | (68) | ||||||
| WU_10.2_7_118557013 | G | A | 13.18 c ± 0.09 | 13.74 b ± 0.17 | 15.19 a ± 0.67 | 0.003 | n.s. | |
| (120) | (33) | (2) | ||||||
| ALGA0044906 | G | A | 13.24 b ± 0.08 | 13.81 a ± 0.20 | - | - | - | |
| (131) | (24) | (0) | ||||||
| Weight Loss 1S, % 5 | ASGA0031014 | G | A | 1.23 b ± 0.04 (42) | n.s. | |||
| WU_10.2_10_74620421 | G | A | 1.24 b ± 0.02 | 1.37 a ± 0.03 | 1.35 ab ± 0.08 | n.s. | n.s. | |
| (91) | (61) | (10) | ||||||
| Weight Loss ES, % 6 | ALGA0022599 | G | A | 2.82 a ± 0.04 | 2.81 a ± 0.05 | 2.90 a ± 0.11 | n.s. | n.s. |
| (86) | (57) | (12) | ||||||
| Salt 1S, % 7 | WU_10.2_14_36295226 | G | A | 1.26 a ± 0.02 | 1.25 ab ± 0.01 | 1.21 b ± 0.01 | 0.036 | n.s. |
| (28) | (85) | (49) | ||||||
| ASGA0000817 | G | A | 1.25 a ± 0.01 | 1.23 a ± 0.01 | 1.24 a ± 0.02 | n.s. | n.s. | |
| (51) | (87) | (24) | ||||||
| Salt ES, % 8 | INRA0000796 | C | A | - | 2.62 a ± 0.03 | 2.63 a ± 0.01 | - | - |
| (0) | (21) | (134) | ||||||
| ASGA0102337 | G | A | 2.62 a ± 0.04 | 2.62 a ± 0.02 | 2.64 a ± 0.02 | n.s. | n.s. | |
| (17) | (73) | (65) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zappaterra, M.; Zambonelli, P.; Schivazappa, C.; Simoncini, N.; Virgili, R.; Stefanon, B.; Davoli, R. Investigating the Features of PDO Green Hams during Salting: Insights for New Markers and Genomic Regions in Commercial Hybrid Pigs. Animals 2021, 11, 68. https://doi.org/10.3390/ani11010068
Zappaterra M, Zambonelli P, Schivazappa C, Simoncini N, Virgili R, Stefanon B, Davoli R. Investigating the Features of PDO Green Hams during Salting: Insights for New Markers and Genomic Regions in Commercial Hybrid Pigs. Animals. 2021; 11(1):68. https://doi.org/10.3390/ani11010068
Chicago/Turabian StyleZappaterra, Martina, Paolo Zambonelli, Cristina Schivazappa, Nicoletta Simoncini, Roberta Virgili, Bruno Stefanon, and Roberta Davoli. 2021. "Investigating the Features of PDO Green Hams during Salting: Insights for New Markers and Genomic Regions in Commercial Hybrid Pigs" Animals 11, no. 1: 68. https://doi.org/10.3390/ani11010068