Heat-Shock Proteins Gene Expression in Peripheral Blood Mononuclear Cells as an Indicator of Heat Stress in Beef Calves



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Experiment
2.1.1. Animals, Sampling and Isolation of PBMCs
2.1.2. Heat Stress Treatment
2.1.3. Measurement of Cell Viability (CCK-8 Assay)
2.2. In Vivo Experiment
2.2.1. Animals and Climatic Chamber and Management
2.2.2. Sampling and Isolation of Peripheral Blood Mononuclear Cells
2.3. Total RNA Extraction and Real-Time PCR Analysis
2.4. Statistical Analysis
3. Results & Discussion
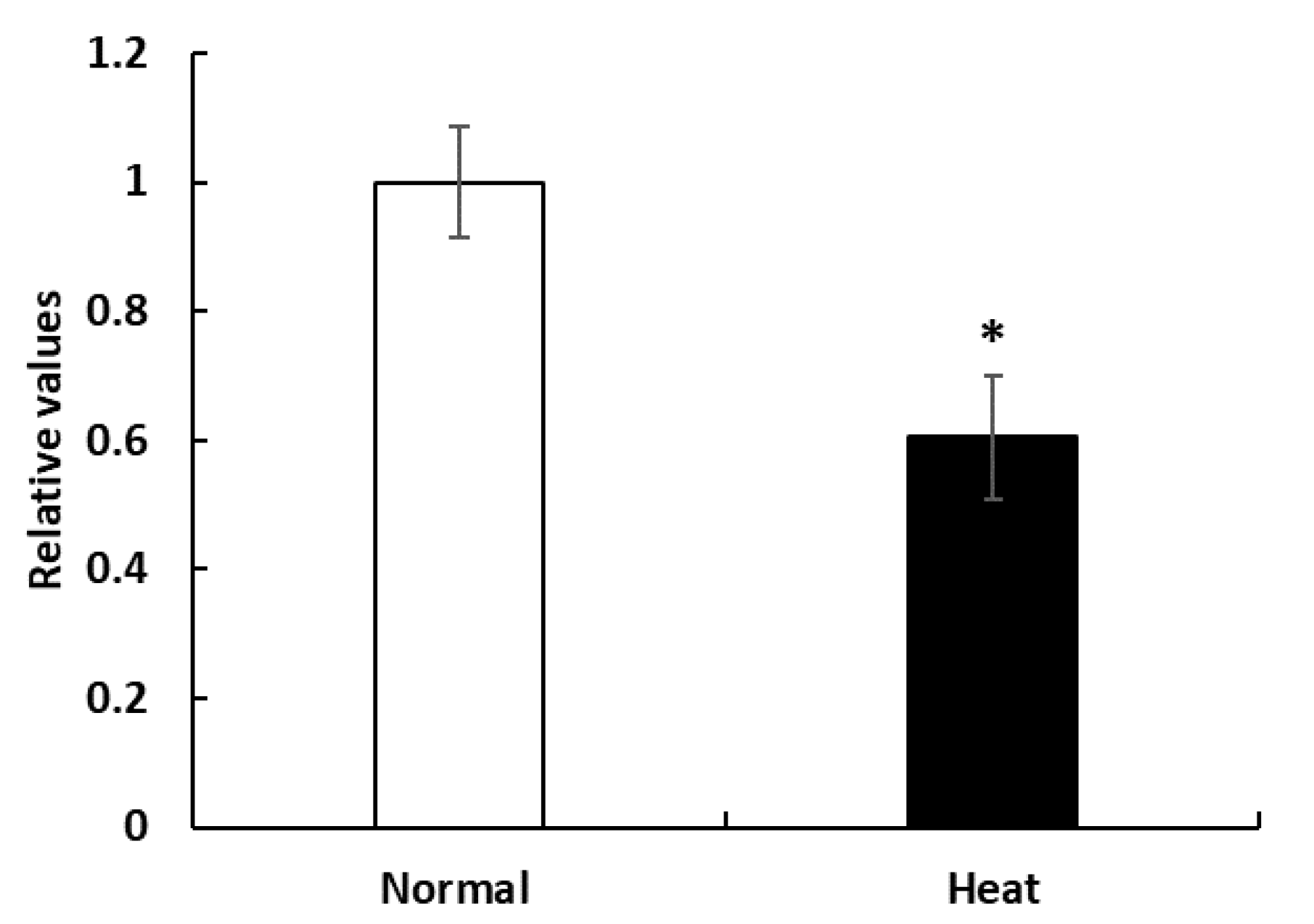
3.1. Proliferation of PBMCs in Cell Culture System
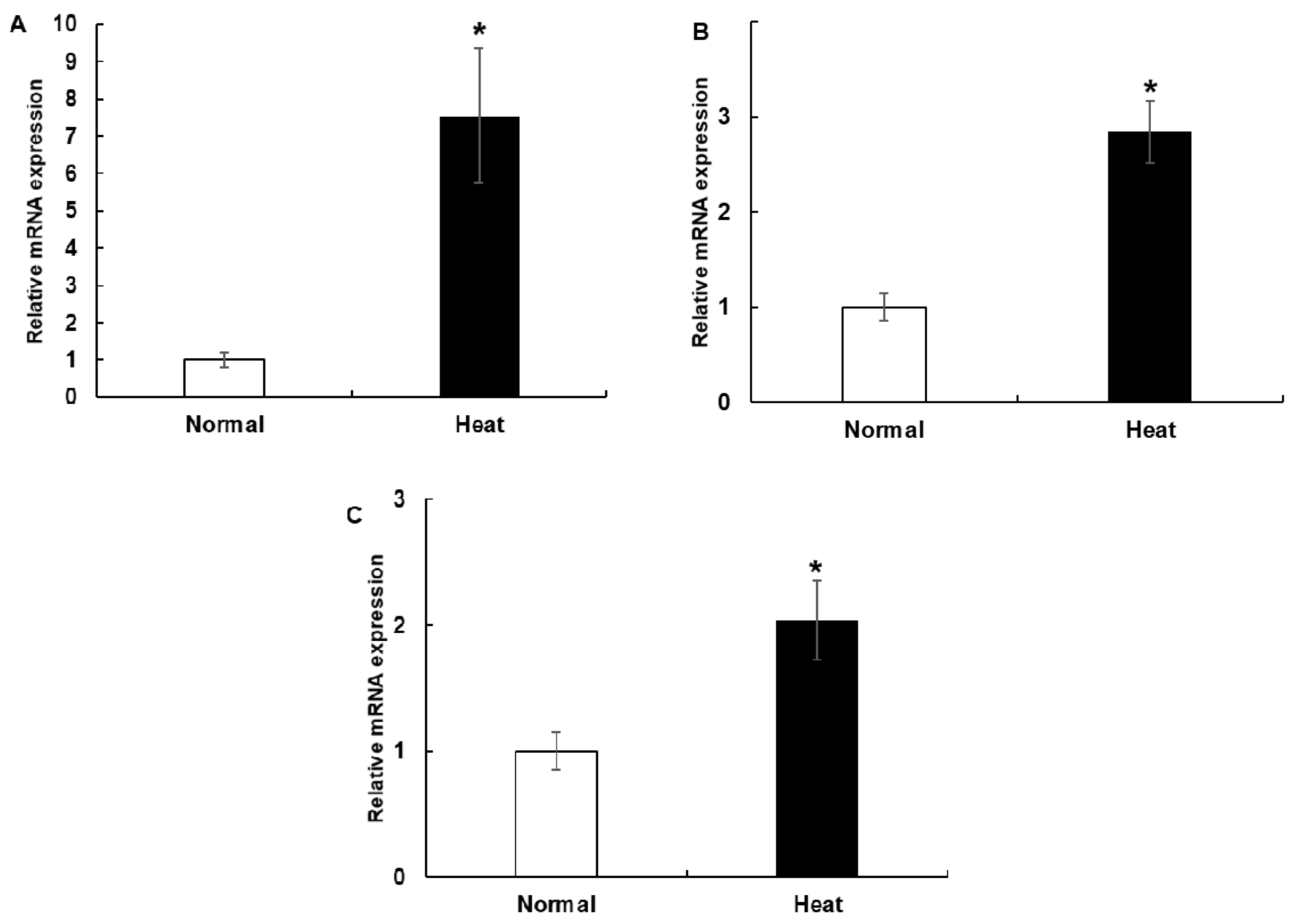
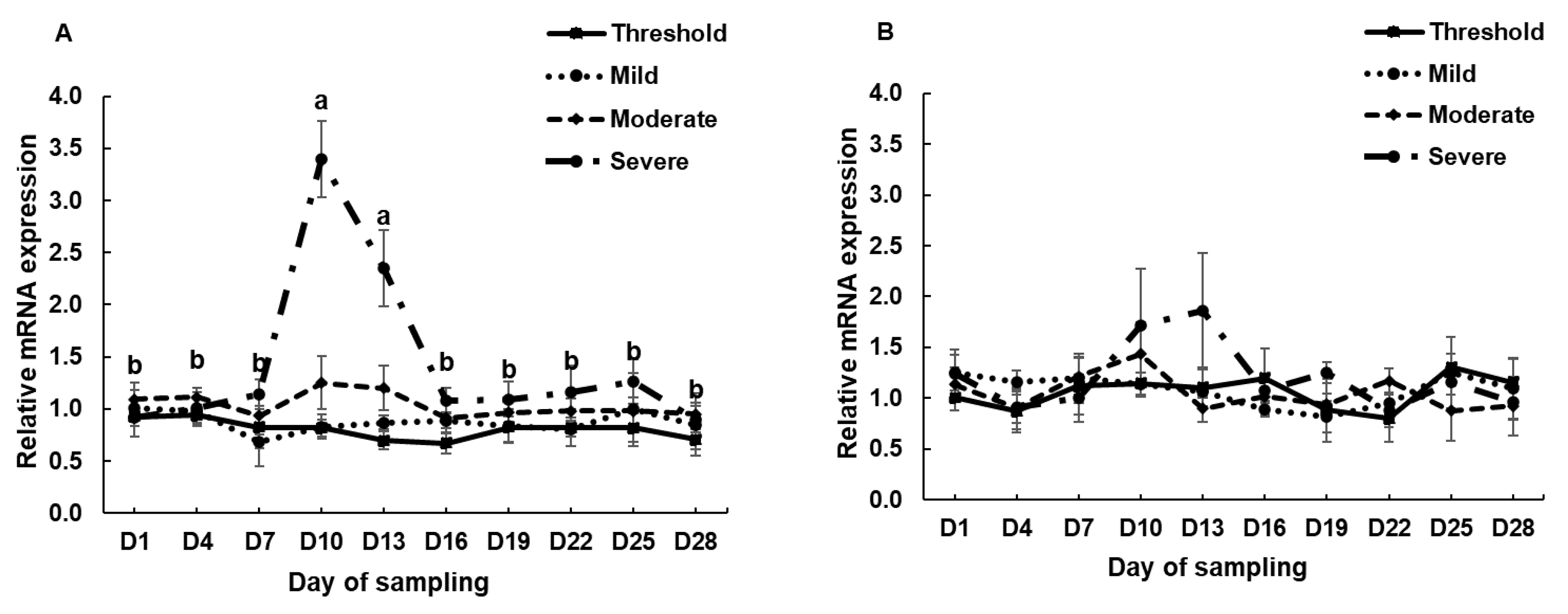
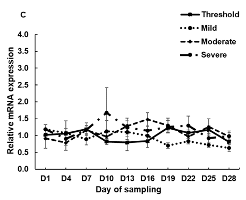
3.2. The mRNA Expression of HSP70, HSP90 and HSPB1 in PBMCs Cell Culture System and during the Experimental Period in the Climatic Chamber
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Kadzere, C.T.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- West, J.W. Effects of Heat-Stress on Production in Dairy Cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- O’Brien, M.D.; Rhoads, R.P.; Sanders, S.R.; Duff, G.C.; Baumgard, L.H. Metabolic adaptations to heat stress in growing cattle. Domest. Anim. Endocrinol. 2010, 38, 86–94. [Google Scholar] [CrossRef]
- Tao, S.; Monteiro, A.P.A.; Thompson, I.M.; Hayen, M.J.; Dahl, G.E. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef] [Green Version]
- Bhanuprakash, V.; Singh, U.; Sengar, G.; Sajjanar, B.; Bhusan, B.; Raja, T.V.; Alex, R.; Kumar, S.; Singh, R.; Ashish, K.; et al. Differential effect of thermal stress on HSP70 expression, nitric oxide production and cell proliferation among native and crossbred dairy cattle. J. Therm. Biol. 2016, 59, 18–25. [Google Scholar] [CrossRef]
- De Maio, A. Heat shock proteins: Facts, thoughts, and dreams. Shock 1999, 11, 1–12. [Google Scholar] [CrossRef]
- Zou, J.; Guo, Y.; Guettouche, T.; Smith, D.F.; Voellmy, R. Repression of Heat Shock Transcription Factor HSF1 Activation by HSP90 (HSP90 Complex) that Forms a Stress-Sensitive Complex with HSF1. Cell 1998, 94, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Ashraf, S.; Goud, T.S.; Grewal, A.; Singh, S.V.; Yadav, B.R.; Upadhyay, R.C. Expression profiling of major heat shock protein genes during different seasons in cattle (Bos indicus) and buffalo (Bubalus bubalis) under tropical climatic condition. J. Therm. Biol. 2015, 51, 55–64. [Google Scholar] [CrossRef]
- Givskov, S.J.; Nygaard, K.T.; Volker, L. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Maloyan, A.; Palmon, A.; Horowitz, M. Heat acclimation increases the basal HSP72 level and alters its production dynamics during heat stress. Am. J. Physiol. 1999, 276, R1506–R1515. [Google Scholar] [CrossRef] [PubMed]
- Lacetera, N.; Bernabucci, U.; Scalia, D.; Basirico, L.; Morera, P.; Nardone, A. Heat stress elicits different responses in peripheral blood mononuclear cells from Brown Swiss and Holstein cows. J. Dairy Sci. 2006, 89, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Kishore, A.; Sodhi, M.; Kumari, P.; Mohanty, A.K.; Sadana, D.K.; Kapila, N.; Khate, K.; Shandilya, U.; Kataria, R.S.; Mukesh, M. Peripheral blood mononuclear cells: A potential cellular system to understand differential heat shock response across native cattle (Bos indicus), exotic cattle (Bos taurus), and riverine buffaloes (Bubalus bubalis) of India. Cell Stress Chaperones 2014, 19, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Bharati, J.; Dangi, S.S.; Mishra, S.R.; Chouhan, V.S.; Verma, V.; Shankar, O.; Bharti, M.K.; Paul, A.; Mahato, D.K.; Rajesh, G.; et al. Expression analysis of Toll like receptors and interleukins in Tharparkar cattle during acclimation to heat stress exposure. J. Therm. Biol. 2017, 65, 48–56. [Google Scholar] [CrossRef]
- Bharati, J.; Dangi, S.S.; Chouhan, V.S.; Mishra, S.R.; Bharti, M.K.; Verma, V.; Shankar, O.; Yadav, V.P.; Das, K.; Paul, A.; et al. Expression dynamics of HSP70 during chronic heat stress in Tharparkar cattle. Int. J. Biometeorol. 2017, 61, 1017–1027. [Google Scholar] [CrossRef]
- Mader, T.L.; Davis, M.S.; Brown-Brandl, T. Environmental factors influencing heat stress in feedlot cattle. J. Anim. Sci. 2006, 84, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Tolnai, S. A method for viable cell count. TCA Man. Tissue Cult. Assoc. 1975, 1, 37–38. [Google Scholar] [CrossRef]
- Kim, W.S.; Nejad, J.G.; Peng, D.Q.; Jung, U.S.; Kim, M.J.; Jo, Y.H.; Jo, J.H.; Lee, J.S.; Lee, H.G. Identification of heat shock protein gene expression in hair follicles as a novel indicator of heat stress in beef calves. Animal 2020, 1–8. [Google Scholar] [CrossRef]
- NRC. A Guide to Environmental Research on Animals; National Academy of Sciences: Washington, DC, USA, 1971. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Kishore, A.; Sodhi, M.; Khate, K.; Kapila, N.; Kumari, P.; Mukesh, M. Selection of stable reference genes in heat stressed peripheral blood mononuclear cells of tropically adapted Indian cattle and buffaloes. Mol. Cell. Probes 2013, 27, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mader, T.L. Environmental stress in confined beef cattle. J. Anim. Sci. 2003, 81, E110–E119. [Google Scholar]
- Kim, W.S.; Lee, J.S.; Jeon, S.W.; Peng, D.Q.; Kim, Y.S.; Bae, M.H.; Jo, Y.H.; Lee, H.G. Correlation between blood, physiological and behavioral parameters in beef calves under heat stress. Asian Australas. J. Anim. Sci. 2018, 31, 919–925. [Google Scholar] [CrossRef]
- Peli, A.; Scagliarini, L.; Bergamini, P.F.; Prosperi, A.; Bernardini, D.; Pietra, M. Influence of heat stress on the immunity in growing beef cattle. Large Anim. Rev. 2013, 19, 215–218. [Google Scholar]
- Lee, J.-S.; Kacem, N.; Kim, W.-S.; Peng, D.Q.; Kim, Y.-J.; Joung, Y.-G.; Lee, C.; Lee, H.-G. Effect of Saccharomyces boulardii Supplementation on Performance and Physiological Traits of Holstein Calves under Heat Stress Conditions. Animals 2019, 9, 510. [Google Scholar] [CrossRef] [Green Version]
- Li, C.M.; Yan, H.C.; Fu, H.L.; Xu, G.F.; Wang, X.Q. Molecular cloning, sequence analysis, and function of the intestinal epithelial stem cell marker Bmi1 in pig intestinal epithelial cells. J. Anim. Sci. 2014, 92, 85–94. [Google Scholar] [CrossRef]
- Costa, M.R.; Ortega, F.; Brill, M.S.; Beckervordersandforth, R.; Petrone, C.; Schroeder, T.; Gotz, M.; Berninger, B. Continuous live imaging of adult neural stem cell division and lineage progression in vitro. Development 2011, 138, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Pechan, P.M. Heat shock proteins and cell proliferation. FEBS Lett. 1991, 280, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Parsell, D.A.; Lindquist, S. The function of heat-shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 27, 437–496. [Google Scholar] [CrossRef]
- Guerriero, V., Jr.; Raynes, D.A. Synthesis of heat stress proteins in lymphocytes from livestock. J. Anim. Sci. 1990, 68, 2779–2783. [Google Scholar] [CrossRef]
- Agnew, L.L.; Colditz, I.G. Development of a method of measuring cellular stress in cattle and sheep. Vet. Immunol. Immunopathol. 2008, 123, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Dangi, S.S.; Dangi, S.K.; Chouhan, V.S.; Verma, M.R.; Kumar, P.; Singh, G.; Sarkar, M. Modulatory effect of betaine on expression dynamics of HSPs during heat stress acclimation in goat (Capra hircus). Gene 2016, 575, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, Y.; Negoro, S.; Kishimoto, S. Heat-shock protein synthesis by human peripheral mononuclear cells from sle patients. Biochem. Biophys. Res. Commun. 1987, 148, 1063–1068. [Google Scholar] [CrossRef]
- Sheikh, A.A.; Aggarwal, A.; Indu, B.; Aarif, O. Inorganic zinc supplementation modulates heat shock and immune response in heat stressed peripheral blood mononuclear cells of periparturient dairy cows. Theriogenology 2017, 95, 75–82. [Google Scholar] [CrossRef]
- Kim, W.; Lee, J.; Peng, D.; Conejos, J.; Jo, Y.; Jo, J.; Seo, J.; Choe, W.; Kim, J.; Kim, T.; et al. Responses of growth performance, physiological and blood parameter to long-term heat stress in beef calves. J. Anim. Sci. 2018, 96, 384–385. [Google Scholar] [CrossRef]
- Voellmy, R. Sensing stress and responding to stress. In Stress-Inducible Cellular Responses; Feige, U., Yahara, I., Morimoto, R.I., Polla, B.S., Eds.; Birkhäuser Basel: Basel, Switzerland, 1996; pp. 121–137. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, M.; Xu, J.; Song, E.; Lv, Y.; Tang, S.; Zhang, X.; Kemper, N.; Hartung, J.; Bao, E. In vitro evaluation of aspirin-induced HspB1 against heat stress damage in chicken myocardial cells. Cell Stress Chaperones 2016, 21, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Urushido, M.; Ide, H.; Ishihara, M.; Hamada-Ode, K.; Shimamura, Y.; Ogata, K.; Inoue, K.; Taniguchi, Y.; Taguchi, T.; et al. Small Heat Shock Protein Beta-1 (HSPB1) Is Upregulated and Regulates Autophagy and Apoptosis of Renal Tubular Cells in Acute Kidney Injury. PLoS ONE 2015, 10, e0126229. [Google Scholar] [CrossRef] [Green Version]
- Concannon, C.G.; Gorman, A.M.; Samali, A. On the role of Hsp27 in regulating apoptosis. Apoptosis Int. J. Program. Cell Death 2003, 8, 61–70. [Google Scholar] [CrossRef]
- Thériault, J.R.; Lambert, H.; Chávez-Zobel, A.T.; Charest, G.; Lavigne, P.; Landry, J. Essential role of the NH2-terminal WD/EPF motif in the phosphorylation-activated protective function of mammalian Hsp27. J. Biol. Chem. 2004, 279, 23463–23471. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Basal Diets | |
|---|---|---|
| Concentrate (60%) | Roughage (Timothy Grass) (40%) | |
| % of dry matter | ||
| Crude protein | 16.70 | 6.16 |
| Ether extract | 3.13 | 1.17 |
| Crude fiber | 8.84 | 36.57 |
| Crude ash | 6.94 | 7.49 |
| ADF | 10.08 | 38.69 |
| NDF | 22.90 | 67.13 |
| Calcium | 1.43 | 0.35 |
| Phosphorus | 0.50 | 0.20 |
| Gene | Accession Number | Sequence | Length (bp) |
|---|---|---|---|
| HSP70 | U09861 | F: TACGTGGCCTTCACCGATACR: GTCGTTGATGACGCGGAAAG | 171 |
| HSP90 | NM_001012670 | F: GGAGGATCACTTGGCTGTCAR: GGGATTAGCTCCTCGCAGTT | 177 |
| HSPB1 | NM_001025569 | F: CCTGGACGTCAACCATTCR: GCTTGCCAGTGATCTCCAC | 77 |
| GAPDH | NM_001034034.2 | F: GGCAAGGTCATCCCTGAGR: GCAGGTCAGATCCACAACAG | 166 |
| RPS15A | NM_001037443.2 | F: CCGTGCTCCAAAGTCATCGTR: GGGAGCAGGTTATTCTGCCA | 200 |
| B2 M | NM_173893.3 | F: GACACCCACCAGAAGATGGAR: CAGGTCTGACTGCTCCGATT | 125 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.-S.; Ghassemi Nejad, J.; Roh, S.-G.; Lee, H.-G. Heat-Shock Proteins Gene Expression in Peripheral Blood Mononuclear Cells as an Indicator of Heat Stress in Beef Calves. Animals 2020, 10, 895. https://doi.org/10.3390/ani10050895
Kim W-S, Ghassemi Nejad J, Roh S-G, Lee H-G. Heat-Shock Proteins Gene Expression in Peripheral Blood Mononuclear Cells as an Indicator of Heat Stress in Beef Calves. Animals. 2020; 10(5):895. https://doi.org/10.3390/ani10050895
Chicago/Turabian StyleKim, Won-Seob, Jalil Ghassemi Nejad, Sang-Gun Roh, and Hong-Gu Lee. 2020. "Heat-Shock Proteins Gene Expression in Peripheral Blood Mononuclear Cells as an Indicator of Heat Stress in Beef Calves" Animals 10, no. 5: 895. https://doi.org/10.3390/ani10050895