Use of the HRM Method in Quick Identification of FecXO Mutation in Highly Prolific Olkuska Sheep


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Material
2.2. Methods
2.2.1. DNA Extraction
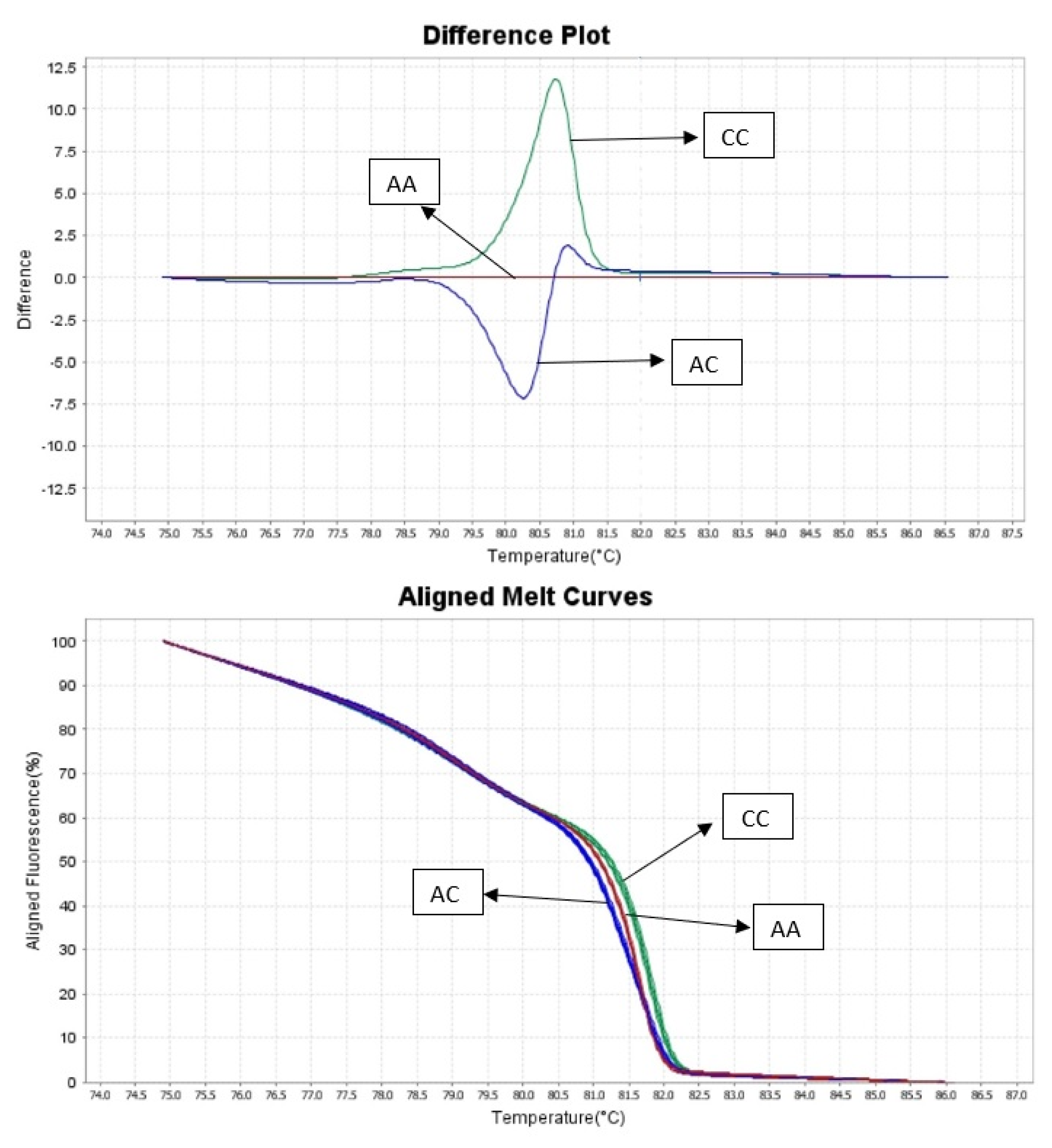
2.2.2. HRM Analysis
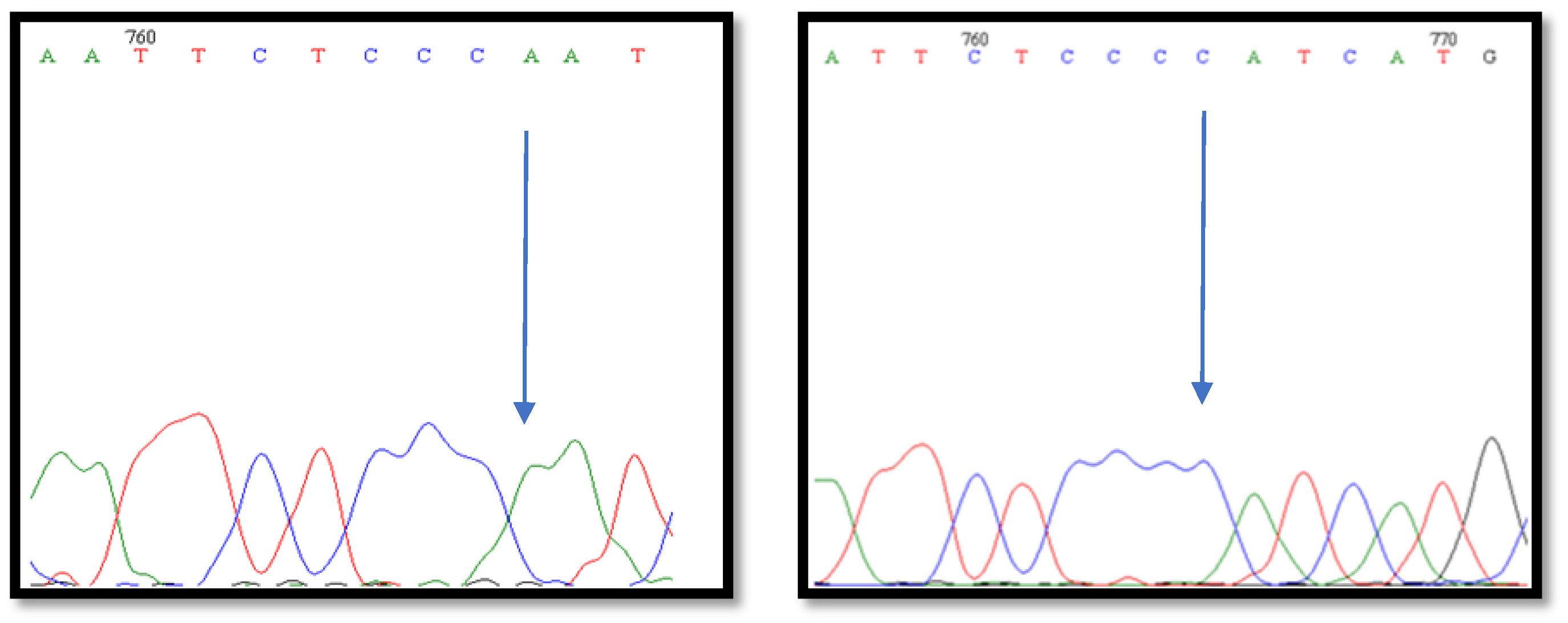
2.2.3. BMP-15 Fragment Gene Sequencing
2.2.4. Statistical Analysis
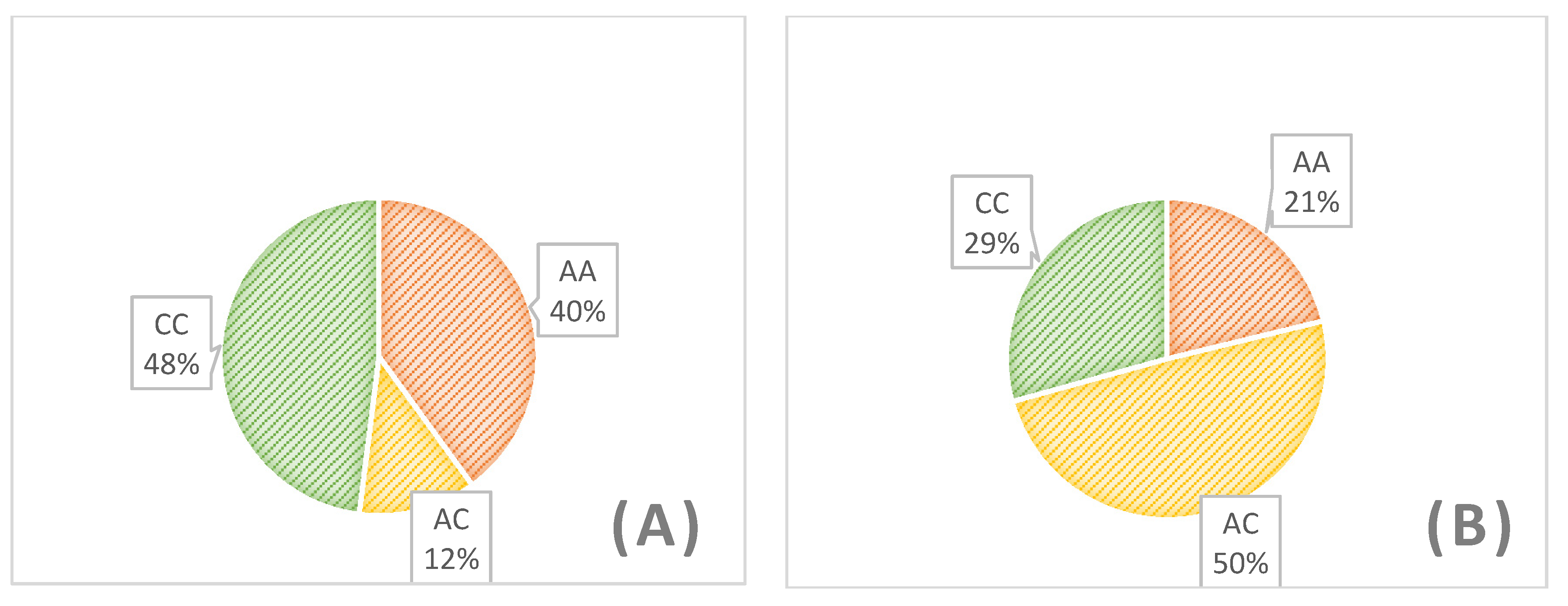
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Davis, G.H. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. GSE 2005, 37 (Suppl. 1), S11–S23. [Google Scholar] [CrossRef] [Green Version]
- Mandon-Pépin, B.; Oustry-Vaiman, A.; Vigier, B.; Piumi, F.; Cribiu, E.; Cotinot, C. Expression Profiles and Chromosomal Localization of Genes Controlling Meiosis and Follicular Development in the Sheep Ovary. Biol. Reprod. 2003, 68, 985–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safari, E.; Fogarty, N.M.; Gilmour, A.R. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest. Prod. Sci. 2005, 92, 271–289. [Google Scholar] [CrossRef]
- Gebreselassie, G.; Berihulay, H.; Jiang, L.; Ma, Y. Review on Genomic Regions and Candidate Genes Associated with Economically Important Production and Reproduction Traits in Sheep (Ovies aries). Animals 2020, 10, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snowder, G.D. Genetic Improvement of Overall Reproductive Successin Sheep: A Review. 2009. Available online: https://tspace.library.utoronto.ca/handle/1807/53119 (accessed on 26 January 2009).
- Abdoli, R.; Zamani, P.; Mirhoseini, S.Z.; Ghavi Hossein-Zadeh, N.; Nadri, S. A review on prolificacy genes in sheep. Reprod. Domest. Anim. Zuchthyg. 2016, 51, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Demars, J.; Fabre, S.; Sarry, J.; Rossetti, R.; Gilbert, H.; Persani, L.; Tosser-Klopp, G.; Mulsant, P.; Nowak, Z.; Drobik, W.; et al. Genome-Wide Association Studies Identify Two Novel BMP15 Mutations Responsible for an Atypical Hyperprolificacy Phenotype in Sheep. PLoS Genet. 2013, 9, e1003482. [Google Scholar] [CrossRef] [PubMed]
- Chantepie, L.; Bodin, L.; Sarry, J.; Woloszyn, F.; Plisson-Petit, F.; Ruesche, J.; Drouilhet, L.; Fabre, S. Genome-Wide identification of a regulatory mutation in BMP15 controlling prolificacy in sheep. bioRxiv 2019. [Google Scholar] [CrossRef]
- Juengel, J.L.; Davis, G.H.; McNatty, K.P. Using sheep lines with mutations in single genes to better understand ovarian function. Reproduction 2013, 146, R111–R123. [Google Scholar] [CrossRef] [PubMed]
- Knight, P.G.; Glister, C. TGF-β superfamily members and ovarian follicle development. Reproduction 2006, 132, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.H. Fecundity genes in sheep. Anim. Reprod. Sci. 2004, 82–83, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Pan, Z.; Wang, X.; Hu, W.; Di, R.; Yao, Y.; Chu, M. Progress on major genes for high fecundity in ewes. Front. Agric. Sci. Eng. 2014, 1, 282. [Google Scholar] [CrossRef] [Green Version]
- Kaczor, U. Genes Involved Litter Size in Olkuska Sheep. Genet. Polymorph. 2017. [Google Scholar] [CrossRef] [Green Version]
- Kucharski, M.; Kaczor, U.; Kaczor, A. Identification of FecXO mutation in the BMP15 gene in prolific Olkuska sheep. Gene 2015, 14, 93–100. [Google Scholar]
- Vossen, R.H.; Aten, E.; Roos, A.; den Dunnen, J.T. High-Resolution Melting Analysis (HRMA)—More than just sequence variant screening. Human Mutation 2009, 30, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Notter, D.R. Genetic improvement of reproductive efficiency of sheep and goats. Anim. Reprod. Sci. 2012, 130, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Słomka, M.; Sobalska-Kwapis, M.; Wachulec, M.; Bartosz, G.; Strapagiel, D. High Resolution Melting (HRM) for High-Throughput Genotyping—Limitations and Caveats in Practical Case Studies. Int. J. Mol. Sci. 2017, 18, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, E.J.; Huynh, B.L. Genotyping by High-Resolution Melting Analysis. In Crop Breeding; Fleury, D., Whitford, R., Eds.; Springer: New York, NY, USA, 2014; Volume 1145, pp. 59–66. ISBN 978-1-4939-0445-7. [Google Scholar]
- Montgomery, J.L.; Sanford, L.N.; Wittwer, C.T. High-Resolution DNA melting analysis in clinical research and diagnostics. Expert Rev. Mol. Diagn. 2010, 10, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Galloway, S.M.; Gregan, S.M.; Wilson, T.; McNatty, K.P.; Juengel, J.L.; Ritvos, O.; Davis, G.H. Bmp15 mutations and ovarian function. Mol. Cell. Endocrinol. 2002, 191, 15–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence [5′–3′] | Analysis Type | PCR Product Length (bp) |
|---|---|---|---|
| BMP-15F | CAGAAGACCAAACCTCTCCCTA | Sanger sequencing | 498 |
| BMP-15R | CTGATTACGCCAGTTTGCAC | ||
| FecXO-F | TCCACCCTTTTCAAGTCAGC | HRM-PCR | 248 |
| FecXO-R | ACTCCCATTTGCCTCAATCA |
| Olkuska Breed | Genotype | Allele | HWE p-Value | |||
|---|---|---|---|---|---|---|
| AA | AC | CC | A | C | ||
| Flock 1 (n = 20) | 0.2 | 0.3 | 0.5 | 0.35 | 0.65 | 0.12764 |
| Flock 2 (n = 20) | 0.5 | 0.05 | 0.45 | 0.525 | 0.475 | 0.000057 |
| Flock 3 (n = 20) | 0.3 | 0.1 | 0.6 | 0.35 | 0.65 | 0.000484 |
| Flock 4(n = 20) | 0.3 | 0.05 | 0.65 | 0.325 | 0.675 | 0.000074 |
| Flock 5 (n = 20) | 0.7 | 0.1 | 0.2 | 0.75 | 0.25 | 0.001040 |
| Total (n = 100) | 0.4 | 0.12 | 0.48 | 0.46 | 0.54 | 0.0000001 |
| Flocks | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| 1 | * | 0.04 | ns | ns | 0.006 |
| 2 | * | ns | ns | ns | |
| 3 | * | ns | 0.01 | ||
| 4 | * | 0.01 | |||
| 5 | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smołucha, G.; Piórkowska, K.; Ropka-Molik, K.; Sikora, J. Use of the HRM Method in Quick Identification of FecXO Mutation in Highly Prolific Olkuska Sheep. Animals 2020, 10, 844. https://doi.org/10.3390/ani10050844
Smołucha G, Piórkowska K, Ropka-Molik K, Sikora J. Use of the HRM Method in Quick Identification of FecXO Mutation in Highly Prolific Olkuska Sheep. Animals. 2020; 10(5):844. https://doi.org/10.3390/ani10050844
Chicago/Turabian StyleSmołucha, Grzegorz, Katarzyna Piórkowska, Katarzyna Ropka-Molik, and Jacek Sikora. 2020. "Use of the HRM Method in Quick Identification of FecXO Mutation in Highly Prolific Olkuska Sheep" Animals 10, no. 5: 844. https://doi.org/10.3390/ani10050844