Validation of Suitable Reference Genes for Gene Expression Studies on Yak Testis Development

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. RNA Isolation and cDNA Synthesis
2.3. Selection of Candidate RGs and Primer Design
2.4. RT-qPCR Assay
2.5. Stability Analysis of RGs
2.6. Validation of Expression of RGs
3. Results
3.1. Primer Specificity, Amplification Efficiency, and Gene Expression Profiles
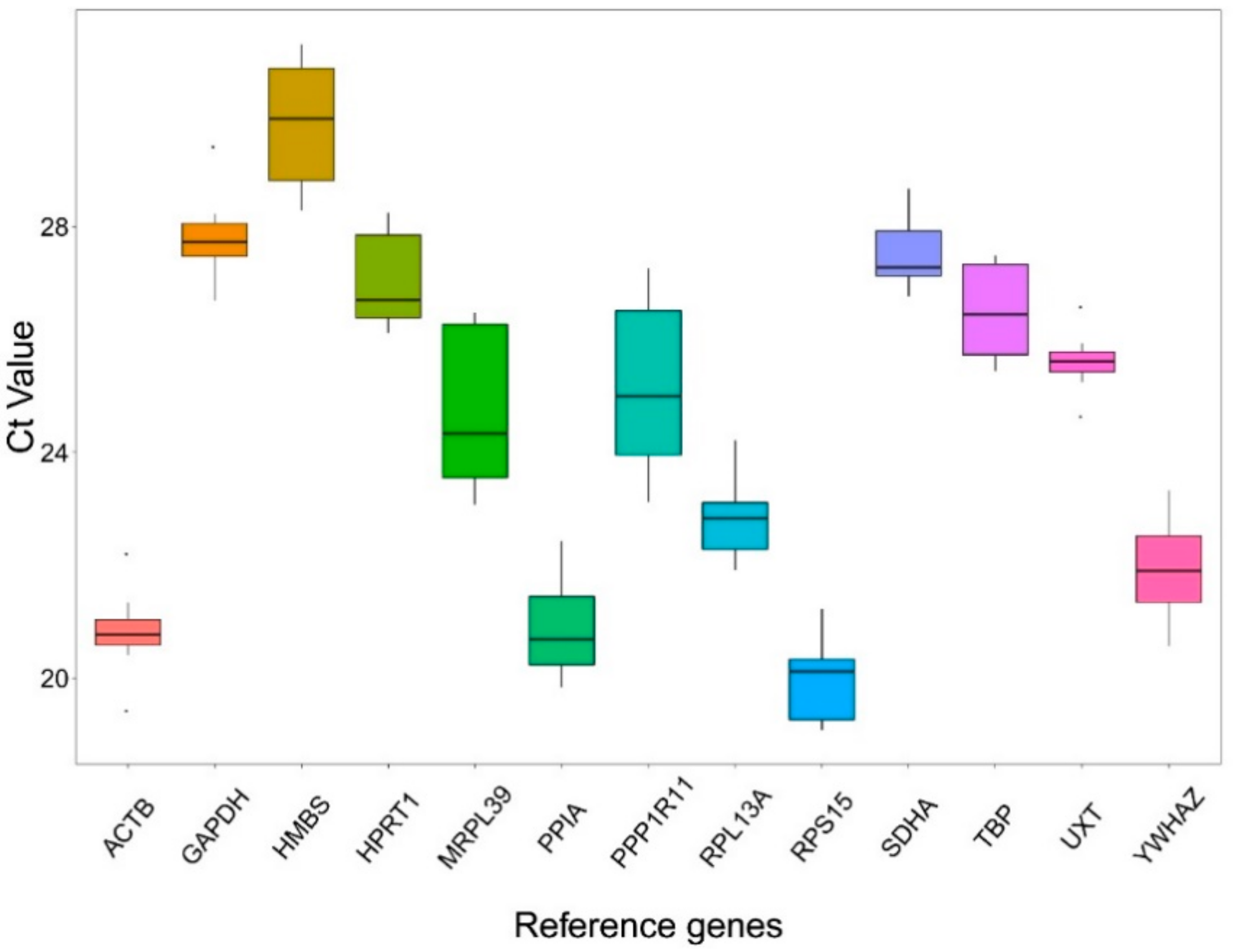
3.2. Ct Value
3.3. Expression Analysis of Candidate RGs in Yak Testis at Different Developmental Stages
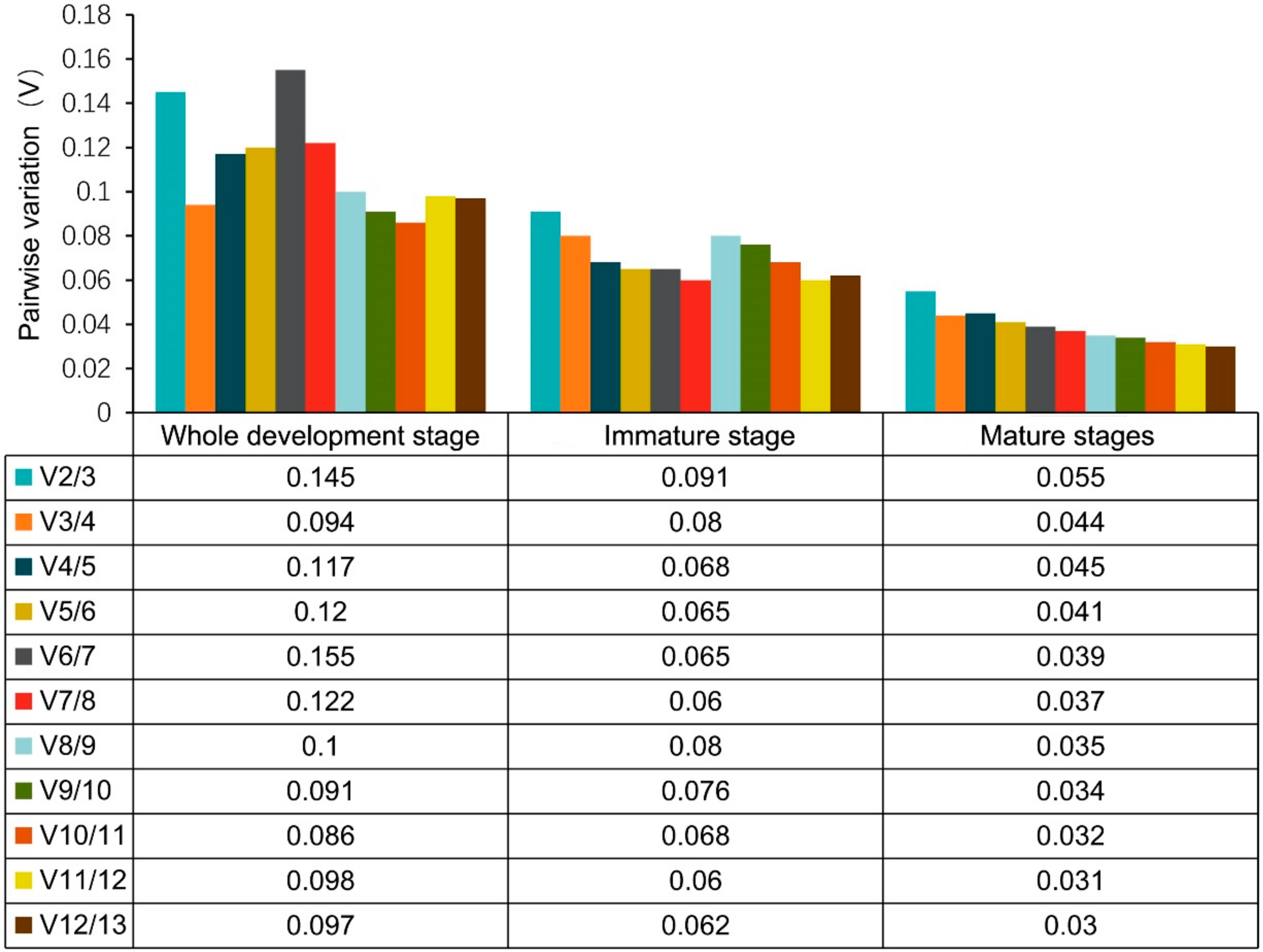
3.3.1. GeNorm Analysis of RGs
3.3.2. NormFinder Analysis of Expression Stability of RGs
3.3.3. BestKeeper Analysis of Expression Stability of RGs
3.3.4. ∆Ct Method Analysis of Expression Stability of RGs
3.3.5. RefFinder Ranking of the Most Stable Genes
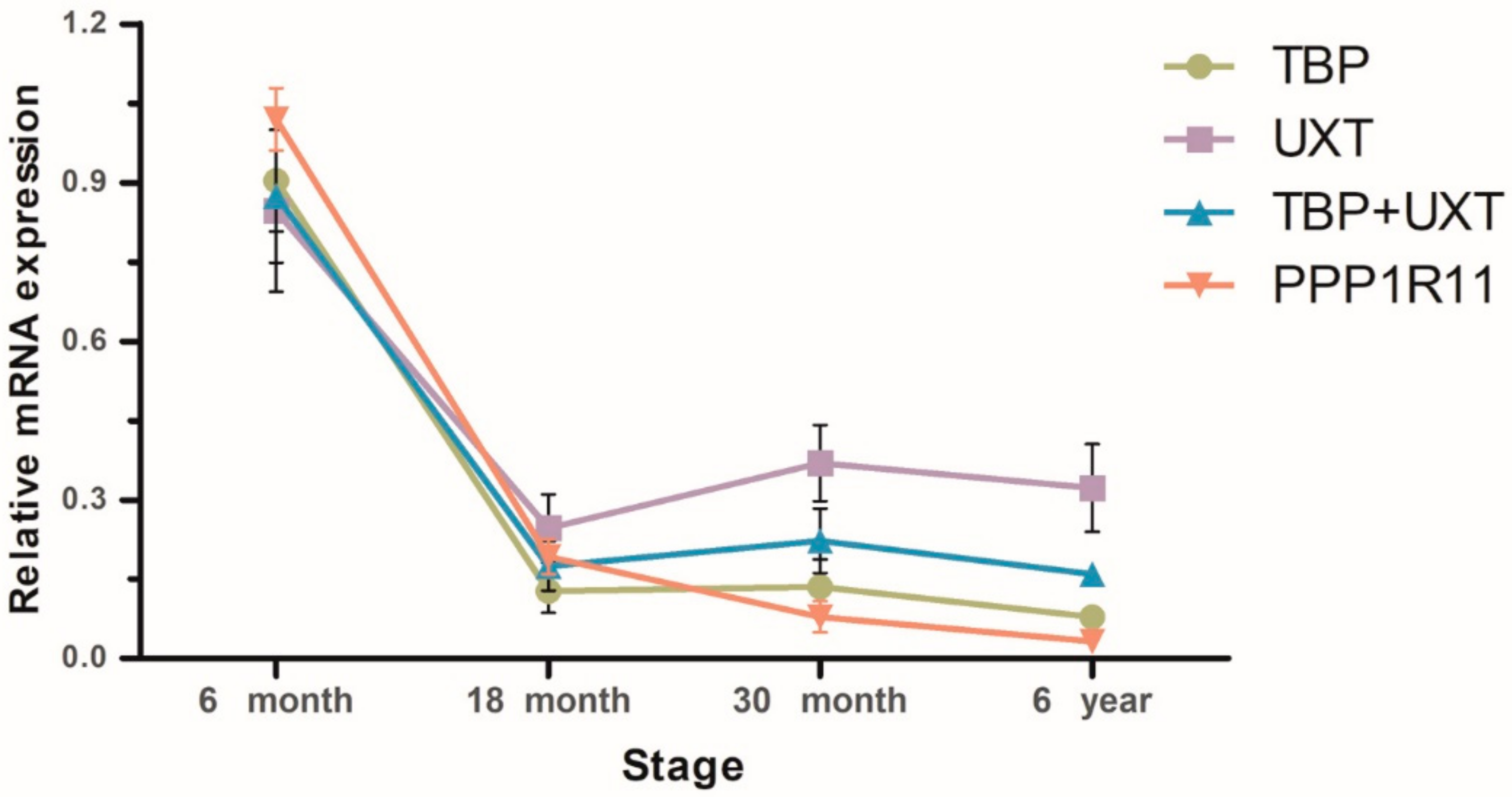
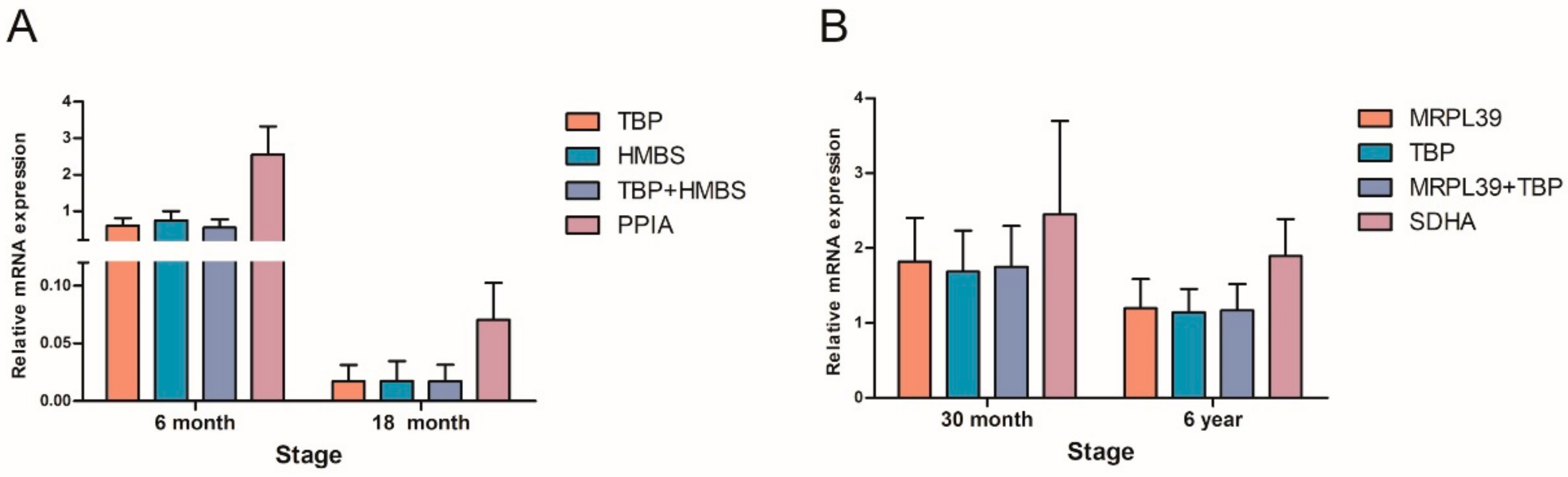
3.4. Validation of Candidate RGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wiener, G.; Han, J.; Long, R. The Yak, 2nd ed.; FAO Regional office for Asia and the Pacific: Bangkok, Thailand, 2003; pp. 1–2. [Google Scholar]
- Quan, Q.; Xing, Q. Environment, Nutrition and Reproduction of Yaks. Anim. Husb. Feed Sci. 2005, 26, 31–32. [Google Scholar]
- Handel, M.A.; Schimenti, J.C. Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nat. Rev. Genet. 2010, 11, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Pan, H.Q.; Zhi, D.J. Growth and development of testes in domesticated and hybrid (wild X domesticated) yak bulls. Anim. Sci. 2000, 70, 481–485. [Google Scholar]
- Zhang, Q.; Wang, Q.; Zhang, Y.; Cheng, S.; Hu, J.; Ma, Y.; Zhao, X. Comprehensive Analysis of MicroRNA–Messenger RNA from White Yak Testis Reveals the Differentially Expressed Molecules Involved in Development and Reproduction. Int. J. Mol. Sci. 2018, 19, 3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasimanickam, V.R.; Kasimanickam, R.K. Differential expression of microRNAs in sexually immature and mature canine testes. Theriogenology 2015, 83, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Johnen, H.; Gonzalez-Silva, L.; Carramolino, L.; Flores, J.M.; Torres, M.; Salvador, J.M. Gadd45g is essential for primary sex determination, male fertility and testis development. PLoS ONE 2013, 8, e58751. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Cui, Y.; Yu, S.; Zhang, Q.; Fan, J.; Abdul Rasheed, B.; Yang, K. The Expression of Epidermal Growth Factor (EGF) and its Receptor (EGFR) During Post-Natal Testes Development in the Yak. Reprod. Domest. Anim. 2014, 49, 970–976. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, X.; Chen, S.; Zheng, L.; He, X.; Liu, M.; Qiao, G.; Wang, Y.; Zhuo, R. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar] [CrossRef] [Green Version]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dheda, K.; Huggett, J.F.; Bustin, S.A.; Johnson, M.A.; Rook, G.; Zumla, A. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques 2004, 37, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Dheda, K.; Huggett, J.F.; Chang, J.S.; Kim, L.U.; Bustin, S.A.; Johnson, M.A.; Rook, G.A.; Zumla, A. The implications of using an inappropriate reference gene for real-time reverse transcription PCR data normalization. Anal. Biochem. 2005, 344, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Lee, J.N.; Bionaz, M.; Deng, X.Y.; Wang, Y. Evaluation of Suitable Internal Control Genes for RT-qPCR in Yak Mammary Tissue during the Lactation Cycle. PLoS ONE 2016, 11, e0147705. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wu, X.; Guo, X.; Bao, P.; Ding, X.; Chu, M.; Liang, C.; Yan, P. Identification of optimal reference genes for examination of gene expression in different tissues of fetal yaks. Czech J. Anim. Sci. 2017, 62, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Bai, W.L.; Yin, R.H.; Zhao, S.J.; Jiang, W.Q.; Yin, R.L.; Ma, Z.J.; Wang, Z.Y.; Zhu, Y.B.; Luo, G.B.; Yang, R.J.; et al. Technical note: Selection of suitable reference genes for studying gene expression in milk somatic cell of yak (Bos grunniens) during the lactation cycle. J. Dairy Sci. 2014, 97, 902–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Wu, X.Y.; Zhou, X.L.; Ding, X.Z.; Chu, M.; Liang, C.N.; Pei, J.; Xiong, L.; Bao, P.J.; Guo, X.; Yan, P. The selection of reference genes for quantitative real-time pcr in the ashidan yak mammary gland during lactation and dry period. Animals 2019, 9, 943. [Google Scholar] [CrossRef] [Green Version]
- Datar, J.; Regassa, A.; Kim, W.K.; Taylor, C.G.; Zahradka, P.; Suh, M. Lipid metabolism is closely associated with normal testicular growth based on global transcriptome profiles in normal and underdeveloped testis of obese zucker (fa/fa) rats. Lipids 2017, 52, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Yazarloo, F.; Shirkoohi, R.; Mobasheri, M.B.; Emami, A.; Modarressi, M.H. Expression analysis of four testis-specific genes AURKC, OIP5, PIWIL2 and TAF7L in acute myeloid leukemia: A gender-dependent expression pattern. Med. Oncol. 2013, 30, 368. [Google Scholar] [CrossRef] [PubMed]
- Brieno-Enriquez, M.A.; Moak, S.L.; Abud-Flores, A.; Cohen, P.E. Characterization of telomeric repeat-containing RNA (TERRA) localization and protein interactions in primordial germ cells of the mouse. Biol. Reprod. 2019, 100, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Wang, S.; Huang, Y.; Zhao, R.; Zhu, Q.; Lin, W. Identification and validation of suitable reference genes for RT-qPCR analysis in mouse testis development. Mol. Genet. Genom. 2014, 289, 1157–1169. [Google Scholar] [CrossRef]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for qu antitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ramskold, D.; Wang, E.T.; Burge, C.B.; Sandberg, R. An abundance of ubiquitously expressed genes revealed by tissue transcriptome sequence data. PLoS Comput. Biol. 2009, 5, e1000598. [Google Scholar] [CrossRef]
- Lecluze, E.; Jégou, B.; Rolland, A.D.; Chalmel, F. New transcriptomic tools to understand testis development and functions. Mol. Cell. Endocrinol. 2018, 468, 47–59. [Google Scholar] [CrossRef]
- Gomes, A.; Stuchi, L.P.; Siqueira, N.; Henrique, J.B.; Vicentini, R.; Ribeiro, M.L.; Darrieux, M.; Ferraz, L. Selection and validation of reference genes for gene expression studies in Klebsiella pneumoniae using Reverse Transcription Quantitative real-time PCR. Sci. Rep. 2018, 8, 9001. [Google Scholar] [CrossRef]
- Lou, Y.N.; Liu, W.J.; Wang, C.L.; Huang, L.; Jin, S.Y.; Lin, Y.Q.; Zheng, Y.C. Histological evaluation and Prdm9 expression level in the testis of sterile male cattle-yaks. Livest. Sci. 2014, 160, 208–213. [Google Scholar] [CrossRef]
- Huang, L.; Jin, S.; Xu, Y.; Li, Y.; Lin, Y.; Zheng, Y. Quantitation of alternative splicing variants of lactate dehydrogenase C gene in testes of adult yak, sexually immature yak calf and sterile male hybrid of yak. Can. J. Anim. Sci. 2012, 92, 291–296. [Google Scholar] [CrossRef]
- Glare, E.M.; Divjak, M.; Bailey, M.J.; Walters, E.H. beta-Actin and GAPDH housekeeping gene expression in asthmatic airways is variable and not suitable for normalising mRNA levels. Thorax 2002, 57, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Li, Y.; Luo, D.; Liao, D.J. Pseudogenes as weaknesses of ACTB (Actb) and GAPDH (Gapdh) used as reference genes in reverse transcription and polymerase chain reactions. PLoS ONE 2012, 7, e41659. [Google Scholar] [CrossRef] [PubMed]
- Neuvians, T.P.; Gashaw, I.; Sauer, C.G.; Ostau, C.V.; Kliesch, S.; Bergmann, M.; Häcker, A.; Grobholz, R. Standardization strategy for quantitative PCR in human seminoma and normal testis. J. Biotechnol. 2005, 117, 163–171. [Google Scholar] [CrossRef]
- Peters, I.R.; Peeters, D.; Helps, C.R.; Day, M.J. Development and application of multiple internal reference (housekeeper) gene assays for accurate normalisation of canine gene expression studies. Vet. Immunol. Immunopathol. 2007, 117, 55–66. [Google Scholar] [CrossRef]
- Hernandez, N. TBP, a universal eukaryotic transcription factor? Genes Dev. 1993, 7, 1291–1308. [Google Scholar] [CrossRef] [Green Version]
- Svingen, T.; Spiller, C.M.; Kashimada, K.; Harley, V.R.; Koopman, P. Identification of Suitable Normalizing Genes for Quantitative Real-Time RT-PCR Analysis of Gene Expression in Fetal Mouse Gonads. Sex. Dev. 2009, 3, 194–204. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Monteiro, A.; Fowler, P.A. Identification of stable endogenous reference genes for real-time PCR in the human fetal gonad using an external standard technique. Mol. Hum. Reprod. 2011, 17, 620–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Ran, M.; Weng, B.; Li, Z.; Dong, L.; Chen, B. Validation of reference genes for quantitative RT-PCR analysis in porcine testis tissues. Sci. Agric. Sin. 2017, 50, 3033–3041. [Google Scholar]
- Zhao, H.; Wang, Q.; Zhang, H.; Liu, Q.; Du, X.; Richter, M.; Greene, M.I. UXT is a novel centrosomal protein essential for cell viability. Mol. Biol. Cell 2005, 16, 5857–5865. [Google Scholar] [CrossRef] [Green Version]
- Tramontana, S.; Bionaz, M.; Sharma, A.; Graugnard, D.E.; Cutler, E.A.; Ajmone-Marsan, P.; Hurley, W.L.; Loor, J.J. Internal controls for quantitative polymerase chain reaction of swine mammary glands during pregnancy and lactation. J. Dairy Sci. 2008, 91, 3057–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashish, S.; Bhure, S.K.; Harikrishna, P.; Ramteke, S.S.; Muhammed Kutty, V.H.; Shruthi, N.; Ravi Kumar, G.V.; Manish, M.; Ghosh, S.K.; Mihir, S. Identification and evaluation of reference genes for accurate gene expression normalization of fresh and frozen-thawed spermatozoa of water buffalo (Bubalus bubalis). Theriogenology 2017, 92, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shen, L.; Gui, L.; Yang, F.; Li, J.; Cao, S.; Zuo, Z.; Ma, X.; Deng, J.; Ren, Z.; et al. Genome-wide DNA methylation profile of prepubertal porcine testis. Reprod. Fert. Dev. 2018, 30, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Wen, L.; Liao, S.; Lin, X.; Ma, T.; Liu, J.; Song, C.X.; Wang, M.; He, C.; Han, C.; et al. Dynamics of 5-hydroxymethylcytosine during mouse spermatogenesis. Nat. Commun. 2013, 4, 1995. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Dansranjavin, T.; Rogenhofer, N.; Oeztuerk, N.; Deuker, J.; Bergmann, M.; Schuppe, H.; Wagenlehner, F.; Weidner, W.; Steger, K.; et al. TET enzymes are successively expressed during human spermatogenesis and their expression level is pivotal for male fertility. Hum. Reprod. 2016, 31, 1411–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Accession No. | Primer Sequence (5′-3′) | Size (bp) |
|---|---|---|---|---|
| ACTB | β-actin | XM_005887322.2 | F: ATTGCCGATGGTGATGAC R: ACGGAGCGTGGCTACAG | 177 |
| GAPDH | glyceraldehyde 3 phosphate dehydrogenase | XM_014482068.1 | F: TCACCAGGGCTGCTTTTA R: CTGTGCCGTTGAACTTGC | 126 |
| UXT | Ubiquitously Expressed Prefoldin Like Chaperone | XM_005899362.2 | F: AGGTGGATTTGGGCTGTAAC R: CTTGGTGAGGTTGTCGCTGA | 170 |
| TBP | TATA box-binding protein | NW_005395834.1 | F: GTCCAATGATGCCTTACGG R: TGCTGCTCCTCCAGAATAGA | 82 |
| YWHAZ | Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein Zeta | XM_005887010.2 | F: AATGTTGTAGGAGCCCGTAG R: CTGCTTGTGAAGCGTTGG | 190 |
| RPL13A | Ribosomal Protein L13a | XM_014481217.1 | F: CAAGCGGATGAACACCAA R: GCAGCAGGAACCACCATT | 192 |
| SDHA | Succinate Dehydrogenase Complex Flavoprotein Subunit A | XM_005894659.2 | F: GGGAACATGGAGGAGGACA R: CCAAAGGCACGCTGGTAGA | 188 |
| RPS15 | Ribosomal Protein S15 | XM_005890466.2 | F: GACCTTCCGCAAGTTCACCT R: ACCACCTCGGGCTTCTCCAT | 198 |
| HPRT1 | hypoxanthine guanine phosphoribosyl transferase 1 | XM_005911180.2 | F: GTGATGAAGGAGATGGG R: ACAGGTCGGCAAAGAAC | 79 |
| PPIA | peptidylprolyl isomerase A | XM_005891872.2 | F: TTTTGAAGCATACAGGTCC R: CCACTCAGTCTTGGCAGT | 98 |
| HMBS | Hydroxymethylbilane Synthase | XM_005897126.2 | F: GAACAAAGGAGCCAAGAAC R: CAGAGGGCTGGGATGTAG | 121 |
| MRPL39 | Mitochondrial Ribosomal Protein L39 | XM_005898618.2 | F: AAACCTTTGACCAAGTCCTGT R: TTCCTCTTTGAATGCCCTCTC | 135 |
| PPP1R11 | Protein Phosphatase 1 Regulatory Inhibitor Subunit 11 | XM_005911410.2 | F: CAGAAAAGACAGAAGGGTGC R: TTCCGAAGTTTGATGGTTAG | 164 |
| TET2 | Tet methyl cytosine dioxygenase 2 | XM_005890479.1 | F: ATGAAAGGAAGCCAAAAGAG R: ATGGAGCCCAGAGAGAGAAG | 126 |
| Rank | GeNorm | NormFinder | Best Keeper | ΔCt | RefFinder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | RPS15 | 0.153 | TBP | 0.345 | UXT | 0.31 | TBP | 0.82 | TBP | 2.74 |
| 2 | RPL13A | 0.153 | YWHAZ | 0.460 | ACTB | 0.43 | YWHAZ | 0.87 | UXT | 3.46 |
| 3 | ACTB | 0.343 | HPRT1 | 0.479 | GAPDH | 0.48 | HPRT1 | 0.88 | YWHAZ | 3.87 |
| 4 | UXT | 0.386 | SDHA | 0.509 | RPL13A | 0.51 | SDHA | 0.94 | RPL13A | 4.47 |
| 5 | GAPDH | 0.485 | PPIA | 0.671 | SDHA | 0.52 | HMBS | 0.96 | SDHA | 4.68 |
| 6 | SDHA | 0.581 | UXT | 0.681 | RPS15 | 0.54 | UXT | 0.97 | ACTB | 4.70 |
| 7 | TBP | 0.739 | HMBS | 0.684 | YWHAZ | 0.69 | PPIA | 1.01 | RPS15 | 5.19 |
| 8 | YWHAZ | 0.816 | GAPDH | 0.855 | TBP | 0.72 | GAPDH | 1.08 | HPRT1 | 5.20 |
| 9 | HPRT1 | 0.857 | ACTB | 0.903 | HPRT1 | 0.76 | ACTB | 1.09 | GAPDH | 5.57 |
| 10 | PPIA | 0.898 | RPL13A | 1.008 | PPIA | 0.77 | RPL13A | 1.13 | PPIA | 7.69 |
| 11 | HMBS | 0.923 | RPS15 | 1.014 | HMBS | 0.94 | RPS15 | 1.14 | HMBS | 8.07 |
| 12 | MRPL39 | 0.975 | MRPL39 | 1.044 | MRPL39 | 1.19 | MRPL39 | 1.17 | MRPL39 | 12.00 |
| 13 | PPP1R11 | 1.033 | PPP1R11 | 1.259 | PPP1R11 | 1.36 | PPP1R11 | 1.35 | PPP1R11 | 13.00 |
| Rank | GeNorm | NormFinder | BestKeeper | ΔCt | RefFinder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | TBP | 0.191 | TBP | 0.258 | UXT | 0.30 | TBP | 0.55 | TBP | 1.57 |
| 2 | HMBS | 0.191 | HMBS | 0.287 | ACTB | 0.32 | HMBS | 0.56 | HMBS | 2.45 |
| 3 | YWHAZ | 0.254 | YWHAZ | 0.395 | RPL13A | 0.34 | YWHAZ | 0.60 | YWHAZ | 3.41 |
| 4 | GAPDH | 0.300 | GAPDH | 0.42 | RPS15 | 0.36 | GAPDH | 0.64 | UXT | 4.05 |
| 5 | PPP1R11 | 0.346 | UXT | 0.446 | YWHAZ | 0.37 | HPRT1 | 0.67 | GAPDH | 4.76 |
| 6 | SDHA | 0.379 | HPRT1 | 0.455 | PPP1R11 | 0.42 | UXT | 0.67 | PPP1R11 | 6.44 |
| 7 | MRPL39 | 0.410 | PPPA1R11 | 0.510 | TBP | 0.42 | PPPA1R11 | 0.69 | ACTB | 6.69 |
| 8 | HPRT1 | 0.442 | MRPL39 | 0.523 | GAPDH | 0.44 | MRPL39 | 0.69 | HPRT1 | 7.00 |
| 9 | UXT | 0.518 | SDHA | 0.528 | HMBS | 0.44 | SDHA | 0.70 | RPL13A | 7.95 |
| 10 | ACTB | 0.577 | ACTB | 0.587 | HPRT1 | 0.48 | ACTB | 0.75 | SDHA | 8.55 |
| 11 | RPL13A | 0.624 | RPL13A | 0.656 | SDHA | 0.49 | RPL13A | 0.79 | MRPL39 | 8.56 |
| 12 | RPS15 | 0.655 | RPS15 | 0.704 | MRPL39 | 0.67 | RPS15 | 0.83 | RPS15 | 9.12 |
| 13 | PPIA | 0.697 | PPIA | 0.807 | PPIA | 0.78 | PPIA | 0.93 | PPIA | 13.00 |
| Rank | GeNorm | NormFinder | BestKeeper | ΔCt | RefFinder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | PPIA | 0.139 | TBP | 0.102 | HPRT1 | 0.16 | TBP | 0.29 | MRPL39 | 2.11 |
| 2 | MRPL39 | 0.139 | MRPL39 | 0.129 | PPIA | 0.28 | MRPL39 | 0.29 | TBP | 2.63 |
| 3 | RPS15 | 0.167 | RPL13A | 0.144 | UXT | 0.28 | RPL13A | 0.30 | PPIA | 3.08 |
| 4 | RPL13A | 0.174 | RPS15 | 0.167 | RPS15 | 0.31 | RPS15 | 0.30 | RPS15 | 3.72 |
| 5 | HPRT1 | 0.204 | ACTB | 0.230 | MRPL39 | 0.32 | PPIA | 0.34 | HPRT1 | 3.96 |
| 6 | TBP | 0.227 | PPIA | 0.248 | ACTB | 0.35 | ACTB | 0.35 | RPL13A | 3.98 |
| 7 | UXT | 0.250 | HPRT1 | 0.252 | RPL13A | 0.36 | HPRT1 | 0.35 | UXT | 6.12 |
| 8 | ACTB | 0.271 | HMBS | 0.286 | TBP | 0.38 | HMBS | 0.38 | ACTB | 6.16 |
| 9 | HMBS | 0.290 | YWHAZ | 0.300 | HMBS | 0.40 | YWHAZ | 0.38 | HMBS | 8.49 |
| 10 | YWHAZ | 0.309 | UXT | 0.325 | PPPA1R11 | 0.42 | UXT | 0.40 | YWHAZ | 9.93 |
| 11 | GAPDH | 0.327 | GAPDH | 0.331 | GAPDH | 0.44 | GAPDH | 0.41 | GAPDH | 11.00 |
| 12 | PPP1R11 | 0.343 | PPP1R11 | 0.374 | YWHAZ | 0.49 | PPP1R11 | 0.44 | PPP1R11 | 11.47 |
| 13 | SDHA | 0.360 | SDHA | 0.383 | SDHA | 0.56 | SDHA | 0.45 | SDHA | 13.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Wu, X.; Chu, M.; Liang, C.; Ding, X.; Pei, J.; Xiong, L.; Bao, P.; Guo, X.; Yan, P. Validation of Suitable Reference Genes for Gene Expression Studies on Yak Testis Development. Animals 2020, 10, 182. https://doi.org/10.3390/ani10020182
Zhou X, Wu X, Chu M, Liang C, Ding X, Pei J, Xiong L, Bao P, Guo X, Yan P. Validation of Suitable Reference Genes for Gene Expression Studies on Yak Testis Development. Animals. 2020; 10(2):182. https://doi.org/10.3390/ani10020182
Chicago/Turabian StyleZhou, Xuelan, Xiaoyun Wu, Min Chu, Chunnian Liang, Xuezhi Ding, Jie Pei, Lin Xiong, Pengjia Bao, Xian Guo, and Ping Yan. 2020. "Validation of Suitable Reference Genes for Gene Expression Studies on Yak Testis Development" Animals 10, no. 2: 182. https://doi.org/10.3390/ani10020182