
Epidemiological Characterisation of blaNDM-Positive Enterobacterales from Food-Producing Animal Farms in Southwest China
 , and
, and 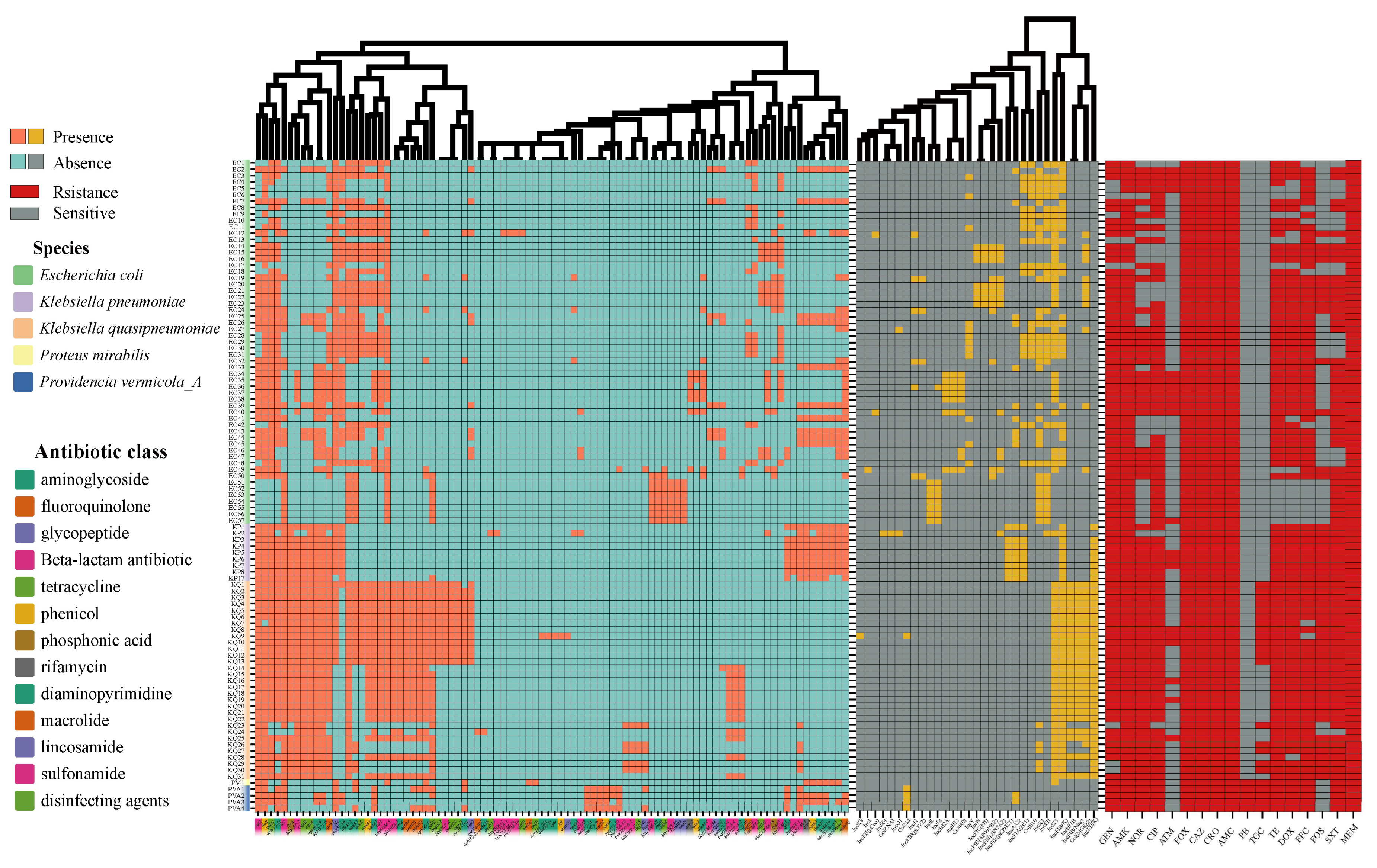{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Strain Isolation, and Identification
2.2. Antibiotic Susceptibility Testing
2.3. Whole Genome Sequencing and Assembly
2.4. Bioinformatics Analysis
2.5. Conjugation Experiment
2.6. Data Availability
3. Results
3.1. Prevalence of blaNDM-Positive Bacteria in Farm Animals
3.2. Antimicrobial Susceptibility, Antibiotic Resistance Genes, and Plasmids of NDM-Positive Strains
3.3. MLST and Phylogenetic Analysis
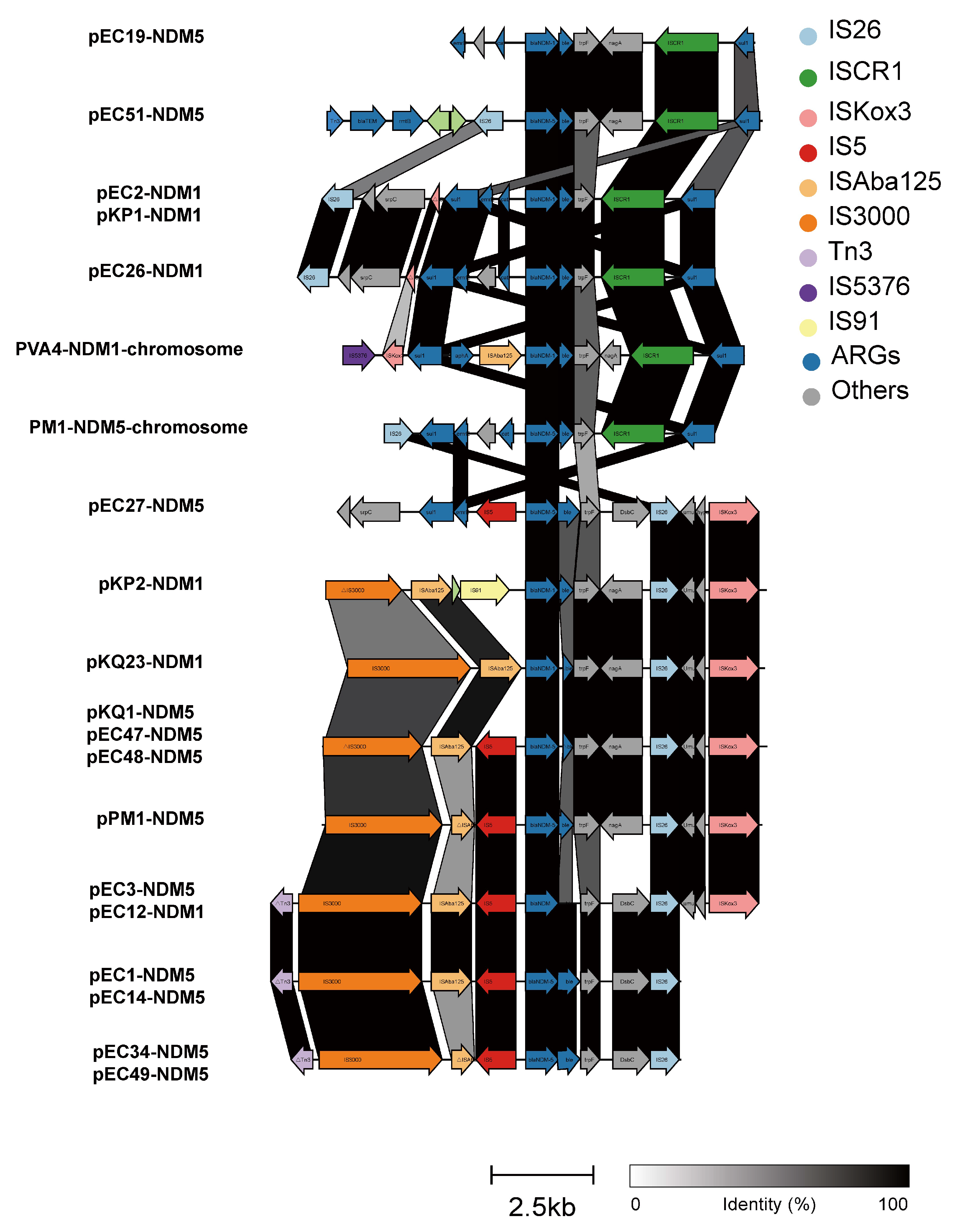
3.4. Genetic Environments and Transferability of blaNDM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bassetti, M.; Peghin, M.; Vena, A.; Giacobbe, D.R. Treatment of Infections Due to MDR Gram-Negative Bacteria. Front. Med. 2019, 6, 74. [Google Scholar] [CrossRef]
- Karaiskos, I.; Giamarellou, H. Multidrug-resistant and extensively drug-resistant Gram-negative pathogens: Current and emerging therapeutic approaches. Expert Opin. Pharmacother. 2014, 15, 1351–1370. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.W.; Harris, P.N.A.; Forde, B.M.; Ben Zakour, N.L.; Catchpoole, E.; Stanton-Cook, M.; Phan, M.D.; Sidjabat, H.E.; Bergh, H.; Heney, C.; et al. Integrating multiple genomic technologies to investigate an outbreak of carbapenemase-producing Enterobacter Hormaechei. Nat. Commun. 2020, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Chudejova, K.; Kraftova, L.; Mattioni Marchetti, V.; Hrabak, J.; Papagiannitsis, C.C.; Bitar, I. Genetic Plurality of OXA/NDM-Encoding Features Characterized from Enterobacterales Recovered from Czech Hospitals. Front. Microbiol. 2021, 12, 641415. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Sugawara, Y.; Akeda, Y.; Hagiya, H.; Sakamoto, N.; Takeuchi, D.; Shanmugakani Rathina, K.; Motooka, D.; Nishi, I.; Zin Khwar, N.; Aye Mya, M.; et al. Spreading Patterns of NDM-Producing Enterobacteriaceae in Clinical and Environmental Settings in Yangon, Myanmar. Antimicrob. Agents Chemother. 2019, 63, e01924-18. [Google Scholar] [CrossRef]
- Zhang, R.; Li, J.; Wang, Y.; Shen, J.; Shen, Z.; Wang, S. Presence of NDM in non-E. coli Enterobacteriaceae in the poultry production environment. J. Antimicrob. Chemother. 2019, 74, 2209–2213. [Google Scholar] [CrossRef]
- Wang, M.G.; Zhang, R.M.; Wang, L.L.; Sun, R.Y.; Bai, S.C.; Han, L.; Fang, L.X.; Sun, J.; Liu, Y.H.; Liao, X.P. Molecular epidemiology of carbapenemase-producing Escherichia coli from duck farms in south-east coastal China. J. Antimicrob. Chemother. 2021, 76, 322–329. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, L.; Zhou, H.; Chan, E.W.; Li, J.; Fang, Y.; Li, Y.; Liao, K.; Chen, S. Nationwide Surveillance of Clinical Carbapenem-resistant Enterobacteriaceae (CRE) Strains in China. EBioMedicine 2017, 19, 98–106. [Google Scholar] [CrossRef]
- Li, J.; Bi, Z.; Ma, S.; Chen, B.; Cai, C.; He, J.; Schwarz, S.; Sun, C.; Zhou, Y.; Yin, J.; et al. Inter-host Transmission of Carbapenemase-Producing Escherichia coli among Humans and Backyard Animals. Environ. Health Perspect. 2019, 127, 107009. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L.; Carrër, A.; Toleman, M.A.; Walsh, T.R. How to detect NDM-1 producers. J. Clin. Microbiol. 2011, 49, 718–721. [Google Scholar] [CrossRef] [PubMed]
- M100-S20; Performance Standards for Antimicrobial Susceptibility Testing. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010.
- VET01; Performance Standards for Antimicrobial Disk and Dilution Susceptibility Test for Bacteria Isolated from Animals. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013.
- Demond, H.; Hanna, C.W.; Castillo-Fernandez, J.; Santos, F.; Papachristou, E.K.; Segonds-Pichon, A.; Kishore, K.; Andrews, S.; D’Santos, C.S.; Kelsey, G. Multi-omics analyses demonstrate a critical role for EHMT1 methyltransferase in transcriptional repression during oogenesis. Genome Res. 2023, 33, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Maiden, M.C.J. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; Van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization by One Table (tvBOT): A web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res. 2023, 51, 587–592. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023, 51, 484–492. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.H. clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Fu, B.; Yin, D.; Sun, C.; Shen, Y.; Liu, D.; Bai, R.; Zhang, R.; Shen, J.; Hu, F.; Wang, Y. Clonal and Horizontal Transmission of blaNDM among Klebsiella pneumoniae in Children’s Intensive Care Units. Microbiol. Spectr. 2022, 10, e0157421. [Google Scholar] [CrossRef]
- Dong, H.; Li, Y.; Cheng, J.; Xia, Z.; Liu, W.; Yan, T.; Chen, F.; Wang, Z.; Li, R.; Shi, J.; et al. Genomic Epidemiology Insights on NDM-Producing Pathogens Revealed the Pivotal Role of Plasmids on blaNDM Transmission. Microbiol. Spectr. 2022, 10, e0215621. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.C.; Kong, L.H.; Wang, Y.L.; Huang, Z.R.; Yang, X.; Zhou, C.Y.; Li, C.; Ma, B.H.; Li, C.; Lei, C.W.; et al. Characterization and Public Health Insights of the New Delhi Metallo-β-Lactamase-Producing Enterobacterales from Laying Hens in China. Microorganisms 2022, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Gao, M.; Jiang, W.; Shi, W.; Li, A.; Liu, S.; Zhang, L.; Zhang, X.; Li, Q.; Lin, H.; et al. Identification of a novel aminoglycoside O-nucleotidyltransferase AadA33 in Providencia vermicola. Front. Microbiol. 2022, 13, 990739. [Google Scholar] [CrossRef]
- Kong, L.H.; Xiang, R.; Wang, Y.L.; Wu, S.K.; Lei, C.W.; Kang, Z.Z.; Chen, Y.P.; Ye, X.L.; Lai, Y.; Wang, H.N. Integration of the blaNDM-1 carbapenemase gene into a novel SXT/R391 integrative and conjugative element in Proteus vulgaris. J. Antimicrob. Chemother. 2020, 75, 1439–1442. [Google Scholar] [CrossRef]
- He, J.; Sun, L.; Zhang, L.; Leptihn, S.; Yu, Y.; Hua, X. A Novel SXT/R391 Integrative and Conjugative Element Carries Two Copies of the blaNDM-1 Gene in Proteus mirabilis. MSphere 2021, 6, e0058821. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, R.; Wei, H.; Zhang, T.; Ma, P.; Wang, Q.; Li, C.; Li, Z.; Lei, C.; Wang, H. Epidemiological Characterisation of blaNDM-Positive Enterobacterales from Food-Producing Animal Farms in Southwest China. Microorganisms 2023, 11, 2304. https://doi.org/10.3390/microorganisms11092304
Wen R, Wei H, Zhang T, Ma P, Wang Q, Li C, Li Z, Lei C, Wang H. Epidemiological Characterisation of blaNDM-Positive Enterobacterales from Food-Producing Animal Farms in Southwest China. Microorganisms. 2023; 11(9):2304. https://doi.org/10.3390/microorganisms11092304
Chicago/Turabian StyleWen, Renqiao, Hongcheng Wei, Tiejun Zhang, Peng Ma, Qin Wang, Chao Li, Zhonghan Li, Changwei Lei, and Hongning Wang. 2023. "Epidemiological Characterisation of blaNDM-Positive Enterobacterales from Food-Producing Animal Farms in Southwest China" Microorganisms 11, no. 9: 2304. https://doi.org/10.3390/microorganisms11092304