
Eight Up-Coming Biotech Tools to Combat Climate Crisis
 , , , , and
, , , , and 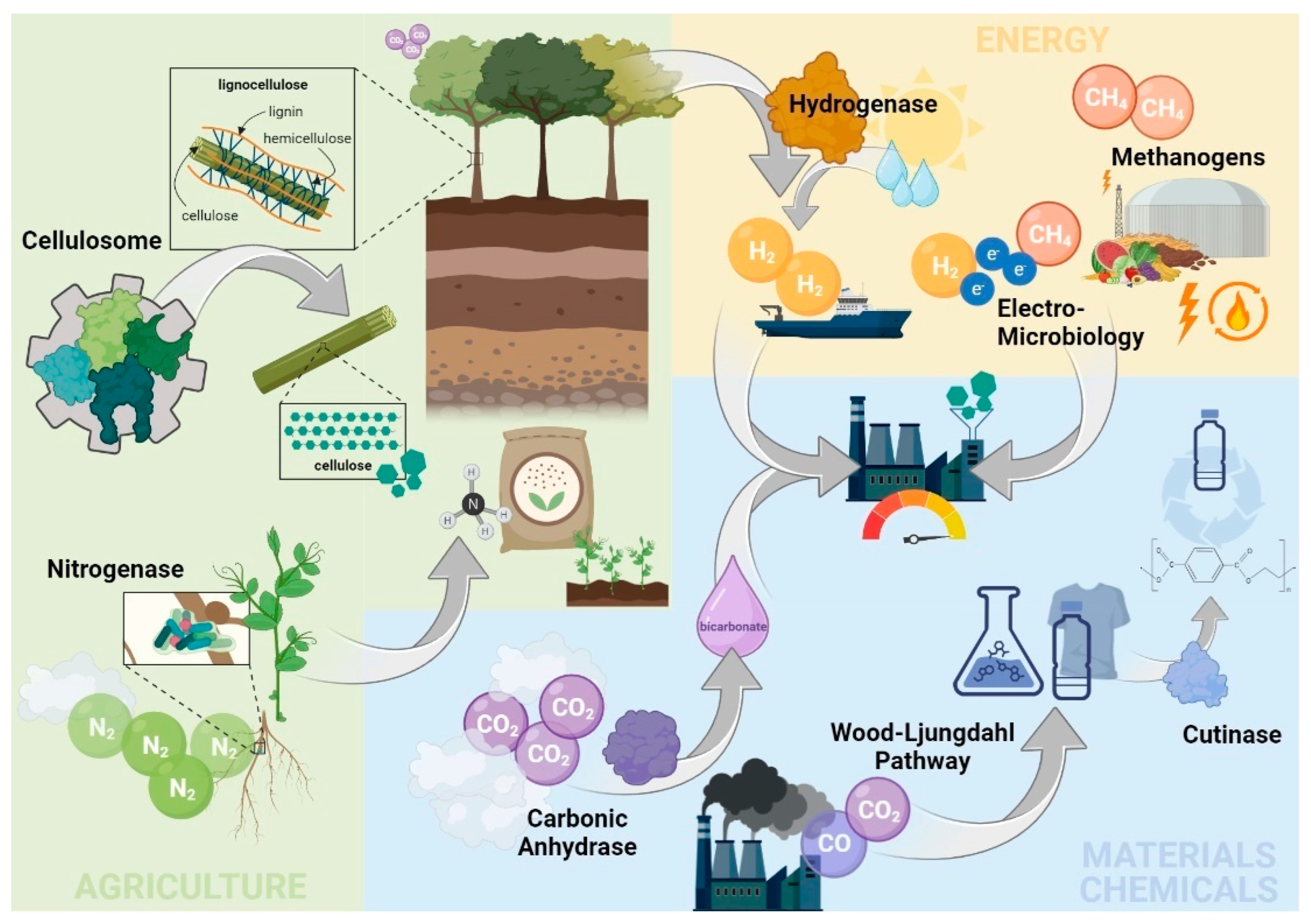{kind=link}
Abstract
:1. Introduction
2. Selected Biotech Tools to Mitigate GHG Emissions with Major Bioeconomy Sectors
2.1. Materials and Chemical Sector
2.1.1. The Wood–Ljungdahl Pathway High-Throughput Microbial CO2 Fixation
2.1.2. Carbonic Anhydrase: An Efficient Enzymatic Capture of CO2
2.1.3. Cutinases: Old Enzymes with a High Relevance for the Future
2.2. Energy Sector
2.2.1. Methanogens: Suppliers of Renewable Natural Gas
2.2.2. Electro Microbiology: Merging Electricity and Biology
2.2.3. Hydrogenases: A Key Enzyme behind the Green Hydrogen Economy
2.3. Agricultural Sector
2.3.1. Cellulosomes: Extracellular Nanomachines for Dismantling Plant Polysaccharides
2.3.2. Nitrogenase: A Bio-Based Approach to Substitute the Energy-Intensive Haber–Bosch Process
3. Potential Impact of Selected Biotechnological Tools
3.1. Wood–Ljungdahl Pathway
3.2. Carbonic Anhydrase
3.3. Cutinase
3.4. Methanogens
3.5. Electro-Microbiology
3.6. Hydrogenases
3.7. Cellulosome
3.8. Nitrogenase
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 6223. [Google Scholar] [CrossRef] [Green Version]
- UNEP. Emissions Gap Report 2022: The Closing Window—Climate Crisis Calls for Rapid Transformation of Societies. Available online: https://www.unep.org/emissions-gap-report-2022 (accessed on 21 December 2022).
- EurEau. Europe’s Water in Figures, An Overview of the European Drinking Water and Waste Water Sectors. Available online: https://www.eureau.org/resources/publications/eureau-publications/5824-europe-s-water-in-figures-2021/file (accessed on 21 December 2022).
- US EPA. EPA Facility Registry Service (FRS): Wastewater Treatment Plants. Available online: https://edg.epa.gov/data/PUBLIC/OEI/OIC/FRS_Wastewater.zip (accessed on 21 December 2022).
- Jain, S. Global Potential of Biogas. Available online: https://www.worldbiogasassociation.org/wp-content/uploads/2019/09/WBA-globalreport-56ppa4_digital-Sept-2019.pdf (accessed on 21 December 2022).
- Bajpai, P. Global Production of Bioethanol. In Developments in Bioethanol. Green Energy and Technology; Springer: Singapore, 2021; pp. 177–196. ISBN 978-981-15-8779-5. [Google Scholar]
- IEA. Implementation of Bioenergy in Brazil—2021 update. Available online: https://www.ieabioenergy.com/wp-content/uploads/2021/11/CountryReport2021_Brazil_final.pdf (accessed on 1 June 2023).
- García, J.L.; Galán, B. Integrating greenhouse gas capture and C1 biotechnology: A key challenge for circular economy. Microb. Biotechnol. 2022, 15, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Li, F.-L.; Wang, Y.; Wangikar, P.P.; Guarnieri, M.T.; Luan, G. Editorial: Bioconversion and Biorefinery of C1 Compounds. Front. Microbiol. 2021, 12, 778962. [Google Scholar] [CrossRef]
- Berg, I.A.; Kockelkorn, D.; Ramos-Vera, W.H.; Say, R.F.; Zarzycki, J.; Hügler, M.; Alber, B.E.; Fuchs, G. Autotrophic carbon fixation in archaea. Nat. Rev. Microbiol. 2010, 8, 447–460. [Google Scholar] [CrossRef]
- Fuchs, G. Alternative pathways of carbon dioxide fixation: Insights into the early evolution of life. Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Eden, G.; Fuchs, G. Total synthesis of acetyl coenzyme a involved in autotrophic CO2 fixation in Acetobacterium woodii. Arch. Microbiol. 1982, 133, 66–74. [Google Scholar] [CrossRef]
- Varma, S.J.; Muchowska, K.B.; Chatelain, P.; Moran, J. Native iron reduces CO2 to intermediates and end-products of the acetyl-CoA pathway. Nat. Ecol. Evol. 2018, 2, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Müller, V.; Chowdhury, N.; Basen, M. Electron bifurcation: A long-hidden energy-coupling mechanism. Annu. Rev. Microbiol. 2018, 72, 331–353. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Dürre, P. Gas fermentation for commodity chemicals and fuels. Microb. Biotechnol. 2017, 10, 1167–1170. [Google Scholar] [CrossRef]
- Katsyv, A.; Müller, V. Overcoming energetic barriers in acetogenic C1 conversion. Front. Bioeng. Biotechnol. 2020, 8, 621166. [Google Scholar] [CrossRef]
- Mock, J.; Zheng, Y.; Mueller, A.P.; Ly, S.; Tran, L.; Segovia, S.; Nagaraju, S.; Köpke, M.; Dürre, P.; Thauer, R.K. Energy conservation associated with ethanol formation from H2 and CO2 in Clostridium autoethanogenum involving electron bifurcation. J. Bacteriol. 2015, 197, 2965–2980. [Google Scholar] [CrossRef] [Green Version]
- Heffernan, J.K.; Valgepea, K.; de Souza Pinto Lemgruber, R.; Casini, I.; Plan, M.; Tappel, R.; Simpson, S.D.; Köpke, M.; Nielsen, L.K.; Marcellin, E. Enhancing CO2-Valorization using Clostridium autoethanogenum for sustainable fuel and chemicals production. Front. Bioeng. Biotechnol. 2020, 8, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demler, M.; Weuster-Botz, D. Reaction engineering analysis of hydrogenotrophic production of acetic acid by Acetobacterium woodii. Biotechnol. Bioeng. 2011, 108, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Kantzow, C.; Mayer, A.; Weuster-Botz, D. Continuous gas fermentation by Acetobacterium woodii in a submerged membrane reactor with full cell retention. J. Biotechnol. 2015, 212, 11–18. [Google Scholar] [CrossRef]
- Steger, F.; Ergal, İ.; Daubek, A.; Loibl, N.; Rachbauer, L.; Fuchs, W.; Rittmann, S.K.-M.R.; Bochmann, G. Trickle-bed bioreactors for acetogenic H2/CO2 conversion. Front. Energy Res. 2022, 10, 842284. [Google Scholar] [CrossRef]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Production of acids and alcohols from syngas in a two-stage continuous fermentation process. Bioresour. Technol. 2018, 253, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Jing, Y.; Lin, Y.; Zhang, S.; An, D. A novel concept for syngas biomethanation by two-stage process: Focusing on the selective conversion of syngas to acetate. Sci. Total Environ. 2018, 645, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.V.; Mohan, S.V.; Chang, Y.-C. Medium-chain fatty acids (MCFA) production through anaerobic fermentation using Clostridium kluyveri: Effect of ethanol and acetate. Appl. Biochem. Biotechnol. 2018, 185, 594–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roghair, M.; Liu, Y.; Strik, D.P.B.T.B.; Weusthuis, R.A.; Bruins, M.E.; Buisman, C.J.N. Development of an effective chain elongation process from acidified food waste and ethanol into n-Caproate. Front. Bioeng. Biotechnol. 2018, 6, 50. [Google Scholar] [CrossRef]
- Stamatopoulou, P.; Malkowski, J.; Conrado, L.; Brown, K.; Scarborough, M. Fermentation of organic residues to beneficial chemicals: A review of medium-chain fatty acid production. Processes 2020, 8, 1571. [Google Scholar] [CrossRef]
- Venkateswar Reddy, M.; Kumar, G.; Mohanakrishna, G.; Shobana, S.; Al-Raoush, R.I. Review on the production of medium and small chain fatty acids through waste valorization and CO2 fixation. Bioresour. Technol. 2020, 309, 123400. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, G.; Farooq, R. Bio-electrochemical synthesis of commodity chemicals by autotrophic acetogens utilizing CO2 for environmental remediation. J. Biosci. 2016, 41, 367–380. [Google Scholar] [CrossRef]
- Candry, P.; Huang, S.; Carvajal-Arroyo, J.M.; Rabaey, K.; Ganigue, R. Enrichment and characterisation of ethanol chain elongating communities from natural and engineered environments. Sci. Rep. 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedorf, H.; Fricke, W.F.; Veith, B.; Brüggemann, H.; Liesegang, H.; Strittmatter, A.; Miethke, M.; Buckel, W.; Hinderberger, J.; Li, F.; et al. The genome of Clostridium kluyveri, a strict anaerobe with unique metabolic features. Proc. Natl. Acad. Sci. USA 2008, 105, 2128–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; Kim, J.-Y.; Park, B.; Choi, I.-G.; Chang, I.S. Genetic engineering system for syngas-utilizing acetogen, Eubacterium limosum KIST612. Bioresour. Technol. Rep. 2020, 11, 100452. [Google Scholar] [CrossRef]
- Jin, S.; Bae, J.; Song, Y.; Pearcy, N.; Shin, J.; Kang, S.; Minton, N.P.; Soucaille, P.; Cho, B.-K. Synthetic biology on acetogenic bacteria for highly efficient conversion of C1 gases to biochemicals. Int. J. Mol. Sci. 2020, 21, 7639. [Google Scholar] [CrossRef]
- Jia, D.; He, M.; Tian, Y.; Shen, S.; Zhu, X.; Wang, Y.; Zhuang, Y.; Jiang, W.; Gu, Y. Metabolic engineering of gas-fermenting Clostridium ljungdahlii for efficient co-production of isopropanol, 3-hydroxybutyrate, and ethanol. ACS Synth. Biol. 2021, 10, 2628–2638. [Google Scholar] [CrossRef]
- Liew, F.E.; Nogle, R.; Abdalla, T.; Rasor, B.J.; Canter, C.; Jensen, R.O.; Wang, L.; Strutz, J.; Chirania, P.; de Tissera, S.; et al. Carbon-negative production of acetone and isopropanol by gas fermentation at industrial pilot scale. Nat. Biotechnol. 2022, 40, 335–344. [Google Scholar] [CrossRef]
- Heijstra, B.D.; Leang, C.; Juminaga, A. Gas fermentation: Cellular engineering possibilities and scale up. Microb. Cell Fact. 2017, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- Benevenuti, C.; Amaral, P.; Ferreira, T.; Seidl, P. Impacts of syngas composition on anaerobic fermentation. Reactions 2021, 2, 391–407. [Google Scholar] [CrossRef]
- US Patent & Trademark Office. Available online: www.uspto.gov/ (accessed on 19 February 2023).
- EEA. Total Greenhous Gas Emission Trends and Projections in Europe. Available online: https://www.eea.europa.eu/ims/total-greenhouse-gas-emission-trends (accessed on 27 September 2022).
- Walsh, B.; Ciais, P.; Janssens, I.A.; Peñuelas, J.; Riahi, K.; Rydzak, F.; van Vuuren, D.P.; Obersteiner, M. Pathways for balancing CO2 emissions and sinks. Nat. Commun. 2017, 8, 14856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skea, J.; Shukla, P.R.; Reisinger, A.; Slade, R.; Pathak, M.; Khourdajie, A.A.; van Diemen, R.; Abdulla, A.; Akimoto, K.; Babiker, M.; et al. Working Group III Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2022—Mitigation of Climate Change. Available online: https://www.ipcc.ch/report/ar6/wg3/downloads/report/IPCC_AR6_WGIII_FullReport.pdf (accessed on 25 February 2023).
- Brickett, L. Carbon Dioxide Capture Handbook. Available online: https://netl.doe.gov/sites/default/files/netl-file/Carbon-Dioxide-Capture-Handbook-2015.pdf (accessed on 25 February 2023).
- Hepburn, C.; Adlen, E.; Beddington, J.; Carter, E.A.; Fuss, S.; Mac Dowell, N.; Minx, J.C.; Smith, P.; Williams, C.K. The technological and economic prospects for CO2 utilization and removal. Nature 2019, 575, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Bui, M.; Adjiman, C.S.; Bardow, A.; Anthony, E.J.; Boston, A.; Brown, S.; Fennell, P.S.; Fuss, S.; Galindo, A.; Hackett, L.A.; et al. Carbon capture and storage (CCS): The way forward. Energy Environ. Sci. 2018, 11, 1062–1176. [Google Scholar] [CrossRef] [Green Version]
- Orlov, A.A.; Valtz, A.; Coquelet, C.; Rozanska, X.; Wimmer, E.; Marcou, G.; Horvath, D.; Poulain, B.; Varnek, A.; de Meyer, F. Computational screening methodology identifies effective solvents for CO2 capture . Commun. Chem. 2022, 5, 1. [Google Scholar] [CrossRef]
- Gervasi, J.; Dubois, L.; Thomas, D. Simulation of the post-combustion CO2 capture with Aspen HysysTM software: Study of different configurations of an absorptionregeneration process for the application to cement flue gases. Energy Procedia 2014, 63, 1018–1028. [Google Scholar] [CrossRef] [Green Version]
- Øi, L.E. Aspen HYSYS simulation of CO2 removal by amine absorption from a gas based power plant. In Proceedings of the 48th Scandinavian Conference on Simulation and Modeling (SIMS 2007), Göteborg (Särö), Sweden, 30–31 October 2007; pp. 73–81. [Google Scholar]
- House, K.Z.; Harvey, C.F.; Aziz, M.J.; Schrag, D.P. The energy penalty of post-combustion CO2 capture & storage and its implications for retrofitting the U.S. installed base. Energy Environ. Sci. 2009, 2, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Grant, T.; Anderson, C.; Hooper, B. Comparative life cycle assessment of potassium carbonate and monoethanolamine solvents for CO2 capture from post combustion flue gases. Int. J. Greenh. Gas Control 2014, 28, 35–44. [Google Scholar] [CrossRef]
- Mazari, S.A.; Si Ali, B.; Jan, B.M.; Saeed, I.M.; Nizamuddin, S. An overview of solvent management and emissions of amine-based CO2 capture technology. Int. J. Greenh. Gas Control 2015, 34, 129–140. [Google Scholar] [CrossRef]
- Shi, J.; Jiang, Y.; Jiang, Z.; Wang, X.; Wang, X.; Zhang, S.; Han, P.; Yang, C. Enzymatic conversion of carbon dioxide. Chem. Soc. Rev. 2015, 44, 5981–6000. [Google Scholar] [CrossRef]
- Obert, R.; Dave, B.C. Enzymatic conversion of carbon dioxide to methanol: Enhanced methanol production in silica sol-gel matrices. J. Am. Chem. Soc. 1999, 121, 12192–12193. [Google Scholar] [CrossRef]
- Xu, S.W.; Lu, Y.; Li, J.; Jiang, Z.-Y.; Wu, H. Efficient conversion of CO2 to methanol catalyzed by three dehydrogenases co-encapsulated in an alginate-silica (ALG-SiO2) hybrid gel. Ind. Eng. Chem. Res. 2006, 45, 4567–4573. [Google Scholar] [CrossRef]
- Jain, S.; Dietrich, H.M.; Müller, V.; Basen, M. Formate is required for growth of the thermophilic acetogenic bacterium Thermoanaerobacter kivui lacking hydrogen-dependent carbon dioxide reductase (HDCR). Front. Microbiol. 2020, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, F.M.; Müller, V. Whole-cell biocatalysis for hydrogen storage and syngas conversion to formate using a thermophilic acetogen. Biotechnol. Biofuels 2020, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.K.; Bhatia, R.K.; Jeon, J.M.; Kumar, G.; Yang, Y.H. Carbon dioxide capture and bioenergy production using biological system—A review. Renew. Sustain. Energy Rev. 2019, 110, 143–158. [Google Scholar] [CrossRef]
- Boucif, N.; Roizard, D.; Favre, E. The Carbonic Anhydrase Promoted Carbon Dioxide Capture. In Membranes for Environmental Applications. Environmental Chemistry for a Sustainable World; no. 42; Zhang, Z., Zhang, W., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–44. ISBN 978-3-030-33978-4. [Google Scholar]
- Jensen, E.L.; Maberly, S.C.; Gontero, B. Insights on the functions and ecophysiological relevance of the diverse carbonic anhydrases in microalgae. Int. J. Mol. Sci. 2020, 21, 2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Capasso, C. A highlight on the inhibition of fungal carbonic anhydrases as drug targets for the antifungal armamentarium. Int. J. Mol. Sci. 2021, 22, 4324. [Google Scholar] [CrossRef]
- Angeli, A.; Carta, F.; Supuran, C.T. Carbonic anhydrases: Versatile and useful biocatalysts in chemistry and biochemistry. Catalysts 2020, 10, 1008. [Google Scholar] [CrossRef]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzym. Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Supuran, C.T. A simple yet multifaceted enzyme. Rev. Chim. 2020, 71, 1–16. [Google Scholar] [CrossRef]
- Lin, J.Y.; Sri Wahyu Effendi, S.; Ng, I.-S. Enhanced carbon capture and utilization (CCU) using heterologous carbonic anhydrase in Chlamydomonas reinhardtii for lutein and lipid production. Bioresour. Technol. 2022, 351, 127009. [Google Scholar] [CrossRef] [PubMed]
- Lindskog, S. Structure and mechanism of carbonic anhydrase. Pharmacol. Ther. 1997, 74, 1–20. [Google Scholar] [CrossRef]
- de Oliveira Maciel, A.; Christakopoulos, P.; Rova, U.; Antonopoulou, I. Carbonic anhydrase to boost CO2 sequestration: Improving carbon capture utilization and storage (CCUS). Chemosphere 2022, 299, 134419. [Google Scholar] [CrossRef]
- di Fiore, A.; Alterio, V.; Monti, S.M.; de Simone, G.; D’Ambrosio, K. Thermostable carbonic anhydrases in biotechnological applications. Int. J. Mol. Sci. 2015, 16, 15456–15480. [Google Scholar] [CrossRef] [Green Version]
- Feron, P.H.M. Exploring the potential for improvement of the energy performance of coal fired power plants with post-combustion capture of carbon dioxide. Int. J. Greenh. 2010, 4, 152–160. [Google Scholar] [CrossRef]
- US 2013/0171718A1; Highly Stable Beta-Class Carbonic Anhydrases Useful in Carbon Capture Systems. Codexis, Inc.: Redwood City, CA, USA, 2013.
- WO 2012/025577 A1; Heat-Stable Persephonella Carbonic Anhydrases and Their Use. WIPO: Geneva, Switzerland, 2012.
- US7892814B2; Heat-Stable Carbonic Anhydrases and Their Use. Codexis, Inc.: Redwood City, CA, USA, 2011.
- WO 201406699 Al; Techniques for CO2 Capture Using Sulfurihydrogenibium sp. Carbonic Anhydrase. WIPO: Geneva, Switzerland, 2014.
- WO2013064195Al; A New Heat-Stable Carbonic Anhydrase and Uses Thereof. WIPO: Geneva, Switzerland, 2013.
- Fradette, L.; Lefebvre, S.; Carley, J. Demonstration results of enzyme-accelerated CO2 capture. Energy Procedia 2017, 114, 1100–1109. [Google Scholar] [CrossRef]
- Alvizo, O.; Nguyen, L.J.; Savile, C.K.; Bresson, J.A.; Lakhapatri, S.L.; Solis, E.O.P.; Fox, R.J.; Broering, J.M.; Benoit, M.R.; Zimmerman, S.A.; et al. Directed evolution of an ultrastable carbonic anhydrase for highly efficient carbon capture from flue gas. Proc. Natl. Acad. Sci. USA 2014, 111, 16436–16441. [Google Scholar] [CrossRef] [Green Version]
- Littlechild, J.A. Enzymes from extreme environments and their industrial applications. Front. Bioeng. Biotechnol. 2015, 3, 161. [Google Scholar] [CrossRef] [Green Version]
- Steger, F.; Reich, J.; Fuchs, W.; Rittmann, S.K.-M.R.; Gübitz, G.M.; Ribitsch, D.; Bochmann, G. Comparison of carbonic anhydrases for CO2 sequestration. Int. J. Mol. Sci. 2022, 23, 957. [Google Scholar] [CrossRef]
- Fabbricino, S.; del Prete, S.; Russo, M.E.; Capasso, C.; Marzocchella, A.; Salatino, P. In vivo immobilized carbonic anhydrase and its effect on the enhancement of CO2 absorption rate. J. Biotechnol. 2021, 336, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-J.; Cheng, J.-C.; Hu, C.-J.; Yu, C.-Y. Entrapment of the fastest known carbonic anhydrase with biomimetic silica and its application for CO2 sequestration. Polym. J. 2021, 13, 2452. [Google Scholar] [CrossRef]
- Rasouli, H.; Iliuta, I.; Bougie, F.; Garnier, A.; Iliuta, M.C. Enhanced CO2 capture in packed-bed column bioreactors with immobilized carbonic anhydrase. J. Chem. Eng. 2022, 432, 134029. [Google Scholar] [CrossRef]
- Rasouli, H.; Nguyen, K.; Iliuta, M.C. Recent advancements in carbonic anhydrase immobilization and its implementation in CO2 capture technologies: A review. Sep. Purif. Technol. 2022, 296, 121299. [Google Scholar] [CrossRef]
- Russo, M.E.; Capasso, C.; Marzocchella, A.; Salatino, P. Immobilization of carbonic anhydrase for CO2 capture and utilization. Appl. Microbiol. Biotechnol. 2022, 106, 3419–3430. [Google Scholar] [CrossRef]
- Saipem. 2020. Available online: https://www.saipem.com/en/media/press-releases/2020-01-22/press-note-saipem-acquired-co2-capture-technology (accessed on 21 December 2022).
- Zaks, A.; Reardon, J. Advanced Low Energy Enzyme Catalyzed Solvent for CO2 Capture. 2013. Available online: https://www.osti.gov/biblio/1121752 (accessed on 21 December 2022).
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant. Sci. 2021, 12, 663165. [Google Scholar] [CrossRef]
- Arya, G.C.; Cohen, H. The Multifaceted Roles of Fungal Cutinases during Infection. J. Fungi 2022, 8, 199. [Google Scholar] [CrossRef]
- Liang, X.; Zou, H. Biotechnological application of cutinase: A powerful tool in synthetic biology. SynBio 2022, 1, 54–64. [Google Scholar] [CrossRef]
- Nikolaivits, E.; Kanelli, M.; Dimarogona, M.; Topakas, E. A middle-aged enzyme still in its prime: Recent advances in the field of cutinases. Catalysts 2018, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Kalantzi, S.; Kekos, D.; Mamma, D. Bioscouring of cotton fabrics by multienzyme combinations: Application of Box–Behnken design and desirability function. Cellulose 2019, 26, 2771–2790. [Google Scholar] [CrossRef]
- Wakelyn, P.J.; Bertoniere, N.R.; French, A.D.; Thibodeaux, D.P.; Triplett, B.A.; Rousselle, M.-A.; Goynes, J.; Edwards, J.V.; Hunter, L.; McAlister, D.D. Cotton Fiber Chemistry and Technology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-042-914-126-3. [Google Scholar]
- Agrawal, P.B.; Nierstrasz, V.A.; Bouwhuis, G.H.; Warmoeskerken, M.M.C.G. Cutinase and pectinase in cotton bioscouring: An innovative and fast bioscouring process. Biocatal. Biotransform. 2008, 26, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Degani, O. Synergism between cutinase and pectinase in the hydrolysis of cotton fibers’ cuticle. Catalysts 2021, 11, 84. [Google Scholar] [CrossRef]
- Lee, S.H.; Song, W.S.; Kim, H.R. Cutinase treatment of cotton fabrics. Fibers Polym. 2009, 10, 802–806. [Google Scholar] [CrossRef]
- Liu, Z.Z. Application of Cutinase in Textile Industry. In Cutinase: Preparation and Application; BP International, Inc.: Arcadia, CA, USA, 2021; pp. 100–121. [Google Scholar] [CrossRef]
- Wilson, A. Big Benefits with Biotech. Innovation in Textiles. 7 November 2022. Available online: https://www.innovationintextiles.com/big-benefits-with-biotech/ (accessed on 21 December 2022).
- Ito, S.; Kobayashi, T.; Hatada, Y.; Horikoshi, K. Enzymes in Modern Detergents. In Microbial Enzymes and Biotransformations; Barredo, J.L., Ed.; Humana Press: Totova, NJ, USA, 2005; Volume 17, pp. 151–161. ISBN 978-1-59259-846-5. [Google Scholar]
- Novozymes. Enzymes—No Compromise on Sustainability. Available online: https://biosolutions.novozymes.com/en/laundry/insights/sustainability (accessed on 21 December 2022).
- BlueWeave. Polyethylene Terephthalate (PET) Resin Market Size, Share & Growth. 2022. Available online: https://www.blueweaveconsulting.com/report/polyethylene-terephthalate-pet-resin-market (accessed on 21 December 2022).
- Eurostat. EU Recycled 41% of Plastic Packaging Waste in 2019. 27 October 2021. Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news/-/ddn-20211027-2 (accessed on 21 December 2022).
- Ronkvist, A.M.; Xie, W.; Lu, W.; Gross, R.A. Cutinase-catalyzed hydrolysis of poly(ethylene terephthalate). Macromolecules 2009, 42, 5128–5138. [Google Scholar] [CrossRef]
- Herrero Acero, E.; Ribitsch, D.; Steinkellner, G.; Gruber, K.; Greimel, K.; Eiteljoerg, I.; Trotscha, E.; Wei, R.; Zimmermann, W.; Zinn, M.; et al. Enzymatic surface hydrolysis of PET: Effect of structural diversity on kinetic properties of cutinases from Thermobifida. Macromolecules 2011, 44, 4632–4640. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, S.; Yamato, S.; Kanaya, E.; Kim, J.J.; Koga, Y.; Takano, K.; Kanaya, S. Isolation of a novel cutinase homolog with polyethylene terephthalate-degrading activity from leaf-branch compost by using a metagenomic approach. Appl. Environ. Microbiol. 2012, 78, 1556–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, F.; Kawabata, T.; Oda, M. Current knowledge on enzymatic PET degradation and its possible application to waste stream management and other fields. Appl. Microbiol. Biotechnol. 2019, 103, 4253–4268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tournier, V.; Topham, C.M.; Gilles, A.; David, B.; Folgoas, C.; Moya-Leclair, E.; Kamionka, E.; Desrousseaux, M.-L.; Texier, H.; Gavalda, S.; et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 2020, 580, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Carbios. Carbios Publishes First Sustainability Report and Underscores Commitment to Circularity and Environmental Responsibilities. 15 December 2022. Available online: https://www.carbios.com/en/carbios-publishes-first-sustainability-report-and-underscores-commitment-to-circularity-and-environmental-responsibilities/ (accessed on 21 December 2022).
- Ramanathan, V.; Cicerone, R.J.; Singh, H.B.; Kiehl, J.T. Trace gas trends and their potential role in climate change. J. Geophys. Res. 1985, 90, 5547–5566. [Google Scholar] [CrossRef] [Green Version]
- Monaco, A.; Ross, K.; Waskow, D.; Ge, M. How Methane Emissions Contribute to Climate Change. Available online: https://www.wri.org/insights/methane-gas-emissions-climate-change (accessed on 19 February 2023).
- Kirschke, S.; Bousquet, P.; Ciais, P.; Saunois, M.; Canadell, J.G.; Dlugokencky, E.J.; Bergamaschi, P.; Bergmann, D.; Blake, D.R.; Bruhwiler, L.; et al. Three decades of global methane sources and sinks. Nat. Geosci. 2013, 6, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.S. An Historical Overview of Methanogenesis. In Methanogenesis: Chapman & Hall Microbiology Series; Ferry, J.G., Ed.; Springer: Boston, MA, USA, 1993; ISBN 978-1-4615-2391-8. [Google Scholar]
- Baker, B.J.; de Anda, V.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, ecology and evolution of archaea. Nat. Microbiol. 2020, 5, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the origin of eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Rittmann, S.K.-M.R. Haloarchaea as emerging big players in future polyhydroxyalkanoate bioproduction: Review of trends and perspectives. Curr. Res. Biotechnol. 2022, 4, 377–391. [Google Scholar] [CrossRef]
- Rittmann, S.K.-M.R.; Pfeifer, K.; Palabikyan, H.; Ergal, İ.; Schuster, B. Archaea in biotechnology | Archaea in der Biotechnologie. BioSpektrum 2021, 27, 96–98. [Google Scholar] [CrossRef]
- Pfeifer, K.; Ergal, İ.; Koller, M.; Basen, M.; Schuster, B.; Rittmann, S.K.-M.R. Archaea Biotechnology. Biotechnol. Adv. 2021, 47, 107668. [Google Scholar] [CrossRef]
- Lyu, Z.; Rotaru, A.-E.; Pimentel, M.; Zhang, C.-J.; Rittmann, S.K.-M.R. Editorial: The methane moment—Cross-boundary significance of methanogens: Preface. Front. Microbiol. 2022, 13, 1055494. [Google Scholar] [CrossRef] [PubMed]
- Ermler, U.; Grabarse, W.; Shima, S.; Goubeaud, M.; Thauer, R.K. Crystal structure of methyl-coenzyme M reductase: The key enzyme of biological methane formation. Science 1997, 278, 1457–1462. [Google Scholar] [CrossRef]
- Kumari ChitturiS, C.M.; Jeevana Lakshmi, J.P.; Hymavathi, D. Methanotrophs—Role in environmental engineering specific to reversal of global warming. Int. J. Sci. Manag. Eng. 2017, 2, 3. [Google Scholar]
- Leu, A.O.; Cai, C.; McIlroy, S.J.; Southam, G.; Orphan, V.J.; Yuan, Z.; Hu, S.; Tyson, G.W. Anaerobic methane oxidation coupled to manganese reduction by members of the Methanoperedenaceae. ISME J. 2020, 14, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Wang, M.; Ya, M.; Xu, C. The denitrifying anaerobic methane oxidation process and microorganisms in the environments: A review. Front. Mar. Sci. 2022, 9, 1038400. [Google Scholar] [CrossRef]
- Kouzuma, A.; Kato, S.; Watanabe, K. Microbial interspecies interactions: Recent findings in syntrophic consortia. Front. Microbiol. 2015, 6, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, Z.; Whitman, W.B. Transplanting the pathway engineering toolbox to methanogens. Curr. Opin. Biotechnol. 2019, 59, 46–54. [Google Scholar] [CrossRef] [PubMed]
- IEA. Outlook for Biogas and Biomethane: Prospects for Organic Growth. 2020. Available online: https://iea.blob.core.windows.net/assets/03aeb10c-c38c-4d10-bcec-de92e9ab815f/Outlook_for_biogas_and_biomethane.pdf (accessed on 21 December 2022).
- Antukh, T.; Lee, I.; Joo, S.; Kim, H. Hydrogenotrophs-Based Biological Biogas Upgrading Technologies. Front. Bioeng. Biotechnol. 2022, 10, 833482. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, S.K.-M.R. A Critical Assessment of Microbiological Biogas to Biomethane Upgrading Systems. In Biogas Science and Technology, Advances in Biochemical Engineering/Biotechnology; Guebitz, G., Bauer, A., Bochmann, G., Gronauer, A., Weiss, S., Eds.; Springer: Cham, Switzerland, 2015; pp. 117–135. ISBN 978-3-319-21993-6. [Google Scholar]
- Thema, M.; Bauer, F.; Sterner, M. Power-to-Gas: Electrolysis and methanation status review. Renew. Sust. Energ. Rev. 2019, 112, 775–787. [Google Scholar] [CrossRef]
- Mauerhofer, L.M.; Zwirtmayr, S.; Pappenreiter, P.; Bernacchi, S.; Seifert, A.H.; Reischl, B.; Schmider, T.; Taubner, R.S.; Paulik, C.; Rittmann, S.K.-M.R. Hyperthermophilic methanogenic archaea act as high-pressure CH4 cell factories. Commun. Biol. 2021, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, S.K.-M.R.; Seifert, A.H.; Bernacchi, S. Kinetics, multivariate statistical modelling, and physiology of CO2-based biological methane production. Appl. Energy 2018, 216, 751–760. [Google Scholar] [CrossRef]
- Wulf, C.; Zapp, P.; Schreiber, A. Review of Power-to-X demonstration projects in Europe. Front. Energy Res. 2020, 8, 191. [Google Scholar] [CrossRef]
- Thema, M.; Weidlich, T.; Hörl, M.; Bellack, A.; Mörs, F.; Hackl, F.; Kohlmayer, M.; Gleich, J.; Stabenau, C.; Trabold, T.; et al. Biological CO2-Methanation: An Approach to Standardization. Energies 2019, 12, 1670. [Google Scholar] [CrossRef] [Green Version]
- Zauner, A.; Fazeni-Fraisl, K.; Wolf-Zoellner, P.; Veseli, A.; Holzleitner, M.-T.; Lehner, M.; Bauer, S.; Pichler, M. Multidisciplinary Assessment of a Novel Carbon Capture and Utilization Concept including Underground Sun Conversion. Energies 2022, 15, 1021. [Google Scholar] [CrossRef]
- Strobel, G.; Hagemann, B.; Huppertz, T.M.; Ganzer, L. Underground bio-methanation: Concept and potential. Renew. Sustain. Energy Rev. 2020, 123, 109747. [Google Scholar] [CrossRef]
- Blasco-Gómez, R.; Batlle-Vilanova, P.; Villano, M.; Balaguer, M.D.; Colprim, J.; Puig, S. On the edge of research and technological application: A critical review of electromethanogenesis. Int. J. Mol. Sci. 2017, 18, 874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydow, A.; Krieg, T.; Mayer, F.; Schrader, J.; Holtmann, D. Electroactive bacteria—Molecular mechanisms and genetic tools. Appl. Microbiol. Biotechnol. 2014, 98, 8481–8495. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.B.A.; Verstraete, W. 100 years of microbial electricity production: Three concepts for the future. Microb. Biotechnol. 2012, 5, 333–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.E.; Dang, Y.; Walker, D.J.F.; Lovley, D.R. The electrically conductive pili of Geobacter species are a recently evolved feature for extracellular electron transfer. Microb. Genom. 2016, 2, 8. [Google Scholar] [CrossRef]
- Faustino, M.M.; Fonseca, B.M.; Costa, N.L.; Lousa, D.; Louro, R.O.; Paquete, C.M. Crossing the wall: Characterization of the multiheme cytochromes involved in the extracellular electron transfer pathway of Thermincola ferriacetica. Microorganisms 2021, 9, 293. [Google Scholar] [CrossRef]
- Ducluzeau, A.-L.; Nitschke, W. When Did Hemes Enter the Scene of Life? On the Natural History of Heme Cofactors and Heme-Containing Enzymes. In Cytochrome Complexes: Evolution, Structures, Energy Transduction, and Signaling. Advances in Photosynthesis and Respiration; Cramer, W., Kallas, T., Eds.; Springer: Dodrecht, The Netherlands, 2016; Volume 41, pp. 13–24. [Google Scholar] [CrossRef]
- Gong, Z.; Yu, H.; Zhang, J.; Li, F.; Song, H. Microbial electro-fermentation for synthesis of chemicals and biofuels driven by bi-directional extracellular electron transfer. Synth. Syst. Biotechnol. 2020, 5, 304–313. [Google Scholar] [CrossRef]
- Moscoviz, R.; Toledo-Alarcón, J.; Trably, E.; Bernet, N. Electro-Fermentation: How To Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef]
- Slate, A.J.; Whitehead, K.A.; Brownson, D.A.C.; Banks, C.E. Microbial fuel cells: An overview of current technology. Renew. Sust. Energ. Rev. 2019, 101, 60–81. [Google Scholar] [CrossRef]
- Thapa, B.S.; Kim, T.; Pandit, S.; Song, Y.E.; Afsharian, Y.P.; Rahimnejad, M.; Kim, J.R.; Oh, S.E. Overview of electroactive microorganisms and electron transfer mechanisms in microbial electrochemistry. Bioresour. Technol. 2022, 347, 126579. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Ghosh, D. Advances in fermentative biohydrogen production: The way forward. Trends Biotechnol. 2009, 27, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Hasibar, B.; Ergal, İ.; Abreu Dias, S.; Bochmann, G.; Rittmann, S.K.-M.R.; Fuchs, W. Competing acetate consumption and production inside a microbial electrolysis cell. J. Environ. Chem. Eng. 2020, 8, 4. [Google Scholar] [CrossRef]
- Litti, Y.V.; Russkova, Y.I.; Zhuravleva, E.A.; Parshina, S.N.; Kovalev, A.A.; Kovalev, D.A.; Nozhevnikova, A.N. Electromethanogenesis: A Promising Biotechnology for the Anaerobic Treatment of Organic Waste. Appl. Biochem. Microbiol. 2022, 58, 19–36. [Google Scholar] [CrossRef]
- Cambrian. Cambrian Innovation. Available online: https://www.cambrianinnovation.com/resources (accessed on 1 June 2023).
- Brahic, C. The electricity eaters. New Sci. 2014, 223, 8–9. [Google Scholar] [CrossRef]
- Walker, G.M.; Ramsey, J.M.; Cavin, R.K.; Herr, D.J.C.; Merzbacher, C.I.; Zhirnov, V. A Framework for Bioelectronics—Discovery and Innovation. 2009. Available online: https://www.nist.gov/system/files/documents/pml/div683/bioelectronics_report.pdf (accessed on 21 December 2022).
- Ukhurebor, K.E.; Adetunji, C.O. Relevance of biosensor in climate smart organic agriculture and their role in environmental sustainability: What has been done and what we need to do? In Biosensors in Agriculture: Recent Trends and Future Perspectives. Concepts and Strategies in Plant Sciences; Pudake, R.N., Jain, U., Kole, C., Eds.; Springer: Cham, Switzerland, 2021; ISBN 978-3-030-66165-6. [Google Scholar]
- Fontecilla-Camps, J.C.; Frey, M.; Garcin, E.; Hatchikian, C.; Montet, Y.; Piras, C.; Vernède, X.; Volbeda, A. Hydrogenase: A hydrogen-metabolizing enzyme. What do the crystal structures tell us about its mode of action. Biochimie 1997, 79, 661–666. [Google Scholar] [CrossRef]
- Vignais, P.M.; Billoud, B. Occurrence, classification, and biological function of hydrogenases: An overview. Chem. Rev. 2007, 107, 4206–4272. [Google Scholar] [CrossRef]
- IEA. Global Hydrogen Review 2021. Available online: https://www.iea.org/reports/global-hydrogen-review-2021/executive-summary (accessed on 21 December 2022).
- Cracknell, J.A.; Vincent, K.A.; Armstrong, F.A. Enzymes as working or inspirational electrocatalysts for fuel cells and electrolysis. Chem. Rev. 2008, 108, 2439–2461. [Google Scholar] [CrossRef]
- Wu, C.H.; McTernan, P.M.; Walter, M.E.; Adams, M.W. Production and Application of a Soluble Hydrogenase from Pyrococcus furiosus. Archaea 2015, 2015, 912582. [Google Scholar] [CrossRef] [Green Version]
- Lubitz, W.; Ogata, H.; Rüdiger, O.; Reijerse, E. Hydrogenases. Chem. Rev. 2014, 114, 4081–4148. [Google Scholar] [CrossRef]
- Burger, Y.; Schwarz, F.M.; Müller, V. Formate-driven H2 production by whole cells of Thermoanaerobacter kivui. Biotechnol. Biofuels Bioprod. 2022, 15, 48. [Google Scholar] [CrossRef]
- Rittmann, S.K.-M.R.; Lee, H.S.; Lim, J.K.; Kim, T.W.; Lee, J.H.; Kang, S.G. One-carbon substrate-based biohydrogen production: Microbes, mechanism, and productivity. Biotechnol. Adv. 2015, 33, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, W.; Ogata, H. Hydrogenases, Structure and Function. In Encyclopedia of Biological Chemistry, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 562–567. [Google Scholar] [CrossRef]
- Thauer, R.K.; Kaster, A.-K.; Goenrich, M.; Schick, M.; Hiromoto, T.; Shima, S. Hydrogenases from Methanogenic Archaea, Nickel, a Novel Cofactor, and H2 Storage. Annu. Rev. Biochem. 2010, 79, 507–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoelmerich, M.C.; Müller, V. Energy-converting hydrogenases: The link between H2 metabolism and energy conservation. Cell Mol. Life Sci. 2020, 77, 1461–1481. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K. The Wolfe cycle comes full circle. Proc. Natl. Acad. Sci. USA 2012, 109, 15084–15085. [Google Scholar] [CrossRef] [Green Version]
- Buckel, W.; Thauer, R.K. Flavin-Based Electron Bifurcation, A New Mechanism of Biological Energy Coupling. Chem. Rev. 2018, 118, 3862–3886. [Google Scholar] [CrossRef] [Green Version]
- Bothe, H.; Schmitz, O.; Yates, M.G.; Newton, W.E. Nitrogen fixation and hydrogen metabolism in cyanobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 529–551. [Google Scholar] [CrossRef] [Green Version]
- Mertens, R.; Liese, A. Biotechnological applications of hydrogenases. Curr. Opin. Biotechnol. 2004, 15, 343–348. [Google Scholar] [CrossRef]
- Zaborsky, O.R.; Benemann, J.R.; Matsunaga, T.; Miyake, J.; San Pietro, A. (Eds.) BioHydrogen; Springer: New York, NY, USA, 1998; ISBN 978-0-585-35132-2. [Google Scholar]
- Ahmed, S.F.; Rafa, N.; Mofijur, M.; Badruddin, I.A.; Inayat, A.; Ali, M.S.; Farrok, O.; Yunus Khan, T.M. Biohydrogen production from biomass sources: Metabolic pathways and economic analysis. Front. Energy Res. 2021, 9, 753878. [Google Scholar] [CrossRef]
- Hardt, S.; Stapf, S.; Filmon, D.T.; Birrell, J.A.; Rüdiger, O.; Fourmond, V.; Léger, C.; Plumeré, N. Reversible H2 Oxidation and Evolution by Hydrogenase Embedded in a Redox Polymer Film. Nat. Catal. 2021, 4, 251–258. [Google Scholar] [CrossRef]
- McPherson, I.J.; Vincent, K.A. Electrocatalysis by hydrogenases: Lessons for building bio-inspired devices. J. Braz. Chem. Soc. 2014, 25, 427–441. [Google Scholar] [CrossRef]
- Park, S.-G.; Rajesh, P.P.; Sim, Y.-U.; Jadhav, D.A.; Noori, M.T.; Kim, D.-H.; Al-Qaradawi, S.Y.; Yang, E.; Jang, J.-K.; Chae, K.-J. Addressing scale-up challenges and enhancement in performance of hydrogen-producing microbial electrolysis cell through electrode modifications. Energy Rep. 2022, 8, 2726–2746. [Google Scholar] [CrossRef]
- Dietrich, H.M.; Righetto, R.D.; Kumar, A.; Wietrzynski, W.; Trischler, R.; Schuller, S.K.; Wagner, J.; Schwarz, F.M.; Engel, B.D.; Müller, V.; et al. Membrane-anchored HDCR nanowires drive hydrogen-powered CO2 fixation. Nature 2022, 607, 823–830. [Google Scholar] [CrossRef]
- Prasad, S.; Ingle, A.P. Impacts of sustainable biofuels production from biomass. In Sustainable Bioenergy: Advances and Impacts; Rai, M., Ingle, A.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 327–346. ISBN 978-012-817-654-2. [Google Scholar]
- Chen, H. Lignocellulose Biorefinery Engineering: Principles and Applications; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9780081001455. [Google Scholar]
- Fontes, C.M.G.A.; Gilbert, H.J. Cellulosomes: Highly efficient nanomachines designed to deconstruct plant cell wall complex carbohydrates. Annu. Rev. Biochem. 2010, 79, 655–681. [Google Scholar] [CrossRef]
- Bayer, E.A.; Lamed, R.; White, B.A.; Flints, H.J. From cellulosomes to cellulosomics. Chem. Rec. 2008, 8, 364–377. [Google Scholar] [CrossRef]
- Duarte, M.; Viegas, A.; Alves, V.D.; Prates, J.A.M.; Ferreira, L.M.A.; Najmudin, S.; Cabrita, E.J.; Carvalho, A.L.; Fontes, C.M.G.A.; Bule, P. A dual cohesin-dockerin complex binding mode in Bacteroides cellulosolvens contributes to the size and complexity of its cellulosome. J. Biol. Chem. 2021, 296, 100552. [Google Scholar] [CrossRef]
- Artzi, L.; Bayer, E.A.; Moraïs, S. Cellulosomes: Bacterial nanomachines for dismantling plant polysaccharides. Nat. Rev. Microbiol. 2017, 15, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Doi, R.H.; Kosugi, A.; Murashima, K.; Tamaru, Y.; Han, S.O. Cellulosomes from mesophilic bacteria. J. Bacteriol. 2003, 185, 5907–5914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, E.A. Cellulosomes and designer cellulosomes: Why toy with Nature? Environ. Microbiol. Rep. 2017, 9, 14–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraïs, S.; Stern, J.; Kahn, A.; Galanopoulou, A.P.; Yoav, S.; Shamshoum, M.; Smith, M.A.; Hatzinikolaou, D.G.; Arnold, F.H.; Bayer, E.A. Enhancement of cellulosome-mediated deconstruction of cellulose by improving enzyme thermostability. Biotechnol. Biofuels 2016, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Arora, R.; Behera, S.; Sharma, N.K.; Kumar, S. Bioprospecting thermostable cellulosomes for efficient biofuel production from lignocellulosic biomass. Bioresour. Bioprocess. 2015, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Li, B.; Feng, Y.; Cui, Q. Consolidated bio-saccharification: Leading lignocellulose bioconversion into the real world. Biotechnol. Adv. 2020, 40, 107535. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.J.; Odaneth, A.A. Industrial application of cellulases. In Current Status and Future Scope of Microbial Cellulases; Tuli, D.K., Kuila, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Chapter 10; pp. 189–209. ISBN 978-012-821-882-2. [Google Scholar]
- Behera, B.C.; Sethi, B.K.; Mishra, R.R.; Dutta, S.K.; Thatoi, H.N. Microbial cellulases—Diversity & biotechnology with reference to mangrove environment: A review. J. Genet. Eng. Biotechnol. 2017, 15, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Resch, M.G.; Donohoe, B.S.; Baker, J.O.; Decker, S.R.; Bayer, E.A.; Beckham, G.T.; Himmel, M.E. Fungal cellulases and complexed cellulosomal enzymes exhibit synergistic mechanisms in cellulose deconstruction. Energy Environ. Sci. 2013, 6, 1858–1867. [Google Scholar] [CrossRef]
- Uekotter, F.; Smil, V. Enriching the earth. Fritz Haber, Carl Bosch, and the transformation of world food production. Environ. Hist. Durh. N. C. 2001, 7, 3. [Google Scholar] [CrossRef]
- Guidehouse Insights. Green Ammonia and the Electrification of the Haber-Bosch Process Reduce Carbon Emissions. 2021. Available online: https://guidehouseinsights.com/news-and-views/green-ammonia-and-the-electrification-of-the-haber-bosch-process-reduce-carbon-emissions (accessed on 21 December 2022).
- Jensen, L.S.; Schjoerring, J.K.; van der Hoek, K.W.; Poulsen, H.D.; Zevenbergen, J.F.; Pallière, C.; Lammel, J.; Brentrup, F.; Jongbloed, A.W.; Willems, J.; et al. Benefits of nitrogen for food, fibre and industrial production from Part I—Nitrogen in Europe: The present position. In The European Nitrogen Assessment Sources, Effects and Policy Perspectives; Sutton, M.A., Howard, C.M., Erisman, J.W., Billen, G., Bleeker, A., Grennfelt, P., van Grinsven, H., Grizzetti, B., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 32–61. [Google Scholar] [CrossRef]
- Hu, Y.; Ribbe, M.W. A journey into the active center of nitrogenase. J. Biol. Inorg. Chem. 2014, 19, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Falkowski, P.G. Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 1997, 387, 272–275. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Peters, J.W. New insights into the evolutionary history of biological nitrogen fixation. Front. Microbiol. 2013, 4, 201. [Google Scholar] [CrossRef] [Green Version]
- Voora, V.; Larrea, C.; Bermúdez, S. Global Market Report: Soybeans. 2020. Available online: https://www.iisd.org/publications/report/global-market-report-soybeans (accessed on 21 December 2022).
- Parnell, J.J.; Berka, R.; Young, H.A.; Sturino, J.M.; Kang, Y.; Barnhart, D.M.; Dileo, M.V. From the lab to the farm: An industrial perspective of plant beneficial microorganisms. Front. Plant Sci. 2016, 7, 1110. [Google Scholar] [CrossRef]
- Burén, S.; Rubio, L.M. State of the art in eukaryotic nitrogenase engineering. FEMS Microbiol. Lett. 2018, 365, 2. [Google Scholar] [CrossRef] [Green Version]
- Xiang, N.; Guo, C.; Liu, J.; Xu, H.; Dixon, R.; Yang, J.; Wang, Y.-P. Using synthetic biology to overcome barriers to stable expression of nitrogenase in eukaryotic organelles. Proc. Natl. Acad. Sci. USA 2020, 117, 16537–16545. [Google Scholar] [CrossRef]
- Wen, A.; Havens, K.; Bloch, S.; Shah, N.; Higgins, D.; Davis-Richardson, A.; Sharon, J.; Rezaei, F.; Mohiti-Asli, M.; Johnson, A.; et al. Enabling Biological Nitrogen Fixation for Cereal Crops in Fertilized Fields. ACS Synth. Biol. 2021, 10, 3264–3277. [Google Scholar] [CrossRef]
- Yao, Q.; Peng, R.; Tian, Y.; Zhu, Y.; Gao, J.; Li, Z.; Fu, X.; Xu, J.; Han, H.; Wang, L.; et al. Endowing plants with the capacity for autogenic nitrogen fixation. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Pankievicz, V.C.S.; Irving, T.B.; Maia, L.G.S.; Ané, J.-M. Are we there yet? The long walk towards the development of efficient symbiotic associations between nitrogen-fixing bacteria and non-leguminous crops. BMC Biol. 2019, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- Vessey, J.K.; Pawlowski, K.; Bergman, B. Root-based N2-fixing symbioses: Legumes, actinorhizal plants, Parasponia sp. and cycads. Plant Soil 2005, 275, 51–78. [Google Scholar] [CrossRef]
- Levy-Varon, J.H.; Batterman, S.A.; Medvigy, D.; Xu, X.; Hall, J.S.; van Breugel, M.; Hedin, L.O. Tropical carbon sink accelerated by symbiotic dinitrogen fixation. Nat. Commun. 2019, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Nygren, P.; Fernández, M.P.; Harmand, J.-M.; Leblanc, H.A. Symbiotic dinitrogen fixation by trees: An underestimated resource in agroforestry systems. Nutr. Cycl. Agroecosyst. 2012, 94, 123–160. [Google Scholar] [CrossRef]
- Rachbauer, L.; Bochmann, G.; Fuchs, W. Gas fermentation. In The Autotrophic Biorefinery: Raw Materials from Biotechnology; Kourist, R., Schmidt, S., Eds.; De Gruyter: Berlin, Germany, 2021; Chapter 4; pp. 85–112. [Google Scholar] [CrossRef]
- Seefeldt, L.C.; Yang, Z.-Y.; Duval, S.; Dean, D.R. Nitrogenase reduction of carbon-containing compounds. Biochim. Biophys. Acta. Bioenerg. 2013, 1827, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oehlmann, N.N.; Rebelein, J.G. The conversion of carbon monoxide and carbon dioxide by nitrogenases. Chem. Bio. Chem. 2022, 23, 8. [Google Scholar] [CrossRef]
- Fixena, K.R.; Zhenga, Y.; Harrisb, D.F.; Shawb, S.; Yangb, Z.-Y.; Deanc, D.R.; Seefeldt, L.C.; Harwooda, C.S. Light-driven carbon dioxide reduction to methane by nitrogenase in a photosynthetic bacterium. Proc. Natl. Acad. Sci. USA 2016, 113, 10163–10167. [Google Scholar] [CrossRef] [Green Version]
- Porc, O.; Carus, M.; Kähler, F.; vom Berg, C. Global Carbon Demand for Chemicals and Materials by Sectors—Reference Years: 2015–2020. Renewable Carbon Publications 2021. Nova Institute. Available online: https://renewable-carbon.eu/publications/product/global-carbon-demand-for-chemicals-and-materials-by-sectors-pdf/ (accessed on 17 April 2023).
- Saadatlu, E.A.; Barzinpour, F.; Yaghoubi, S. A sustainable model for municipal solid waste system considering global warming potential impact: A case study. Comput. Ind. Eng. 2022, 169, 108127. [Google Scholar] [CrossRef]
- Gao, N.; Kamran, K.; Quan, C.; Williams, P.T. Thermochemical conversion of sewage sludge: A critical review. Prog. Energy Combust. Sci. 2020, 79, 100843. [Google Scholar] [CrossRef]
- Xue, X.; Chen, D.; Song, X.; Xiaohu, D. Hydrothermal and Pyrolysis Treatment for Sewage Sludge: Choice from Product and from Energy Benefit. Energy Procedia 2015, 66, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.; Kopf, M.H. Alternative Carbon Sources for Chemical Value Chains. BASF Research Press Conference 2022. Available online: https://www.basf.com/global/documents/en/news-and-media/events/2022/research-press-conference/BASF-Research-Press-Conference-2022_BASF-and-LanzaTech.pdf (accessed on 28 April 2023).
- International Energy Agency (IEA). Carbon Capture, Utilisation and Storage. 2021. Available online: https://www.iea.org/fuels-and-technologies/carbon-capture-utilisation-and-storage (accessed on 28 March 2023).
- da Silveira Cachola, C.; Ciotta, M.; dos Santos, A.A.; Peyerl, D. Deploying of the carbon capture technologies for CO2 emission mitigation in the industrial sectors. Carbon Capture Sci. Technol. 2023, 7, 100102. [Google Scholar] [CrossRef]
- Talekar, S.; Byung Hoon Jo, B.H.; Dordick, J.S.; Kim, J. Carbonic anhydrase for CO2 capture, conversion and utilization. Curr. Opin. Biotechnol. 2022, 74, 230–240. [Google Scholar] [CrossRef]
- Soong, Y.-H.V.; Sobkowicz, M.J.; Xie, D. Recent advances in biological recycling of polyethylene terephthalate (PET) plastic wastes. Bioengineering 2022, 9, 98. [Google Scholar] [CrossRef]
- Lamberti, F.M.; Román-Ramírez, L.A.; Wood, J. Recycling of bioplastics: Routes and benefits. J. Polym. Environ. 2020, 28, 2551–2571. [Google Scholar] [CrossRef]
- Turner, D.A.; Williams, I.D.; Kemp, S. Greenhouse gas emission factors for recycling of source-segregated waste materials. Resour. Conserv. Recy. 2015, 105, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Guidehouse. Biomethane Production Potentials in the EU—Feasibility of REPowerEU 2030 Targets, Production Potentials in the Member States and Outlook to 2050. July 2022. Available online: https://gasforclimate2050.eu/wp-content/uploads/2022/10/Guidehouse_GfC_report_design_final_v3.pdf (accessed on 3 April 2023).
- Market Observatory for Energy DG Energy. Quarterly Report on European Gas Markets—With Focus on 2021, an Extraordinary Year on the European and Global Gas Markets. 2021. Available online: https://energy.ec.europa.eu/system/files/2022-04/Quarterly%20report%20on%20European%20gas%20markets_Q4%202021.pdf (accessed on 3 April 2023).
- Zhang, M.; Ma, Y. Energy use and challenges in current wastewater treatment plants. In A-B Processes: Towards Energy Self-sufficient Municipal Wastewater Treatment; IWA Publishing: London, UK, 2019; Chapter 1. [Google Scholar] [CrossRef] [Green Version]
- Flörke, M.; Kynast, E.; Bärlund, I.; Eisner, S.; Wimmer, F.; Alcamo, J. Domestic and industrial water uses of the past 60 years as a mirror of socio-economic development: A global simulation study. Glob. Environ. Chang. 2013, 23, 144–156. [Google Scholar] [CrossRef]
- Sato, T.; Qadir, M.; Yamamoto, S.; Endo, T.; Zahoor, A. Global, regional, and country level need for data on wastewater generation, treatment, and use. Agric. Water Manag. 2013, 130, 1–13. [Google Scholar] [CrossRef]
- European Investment Bank (EIB). EIB Project Carbon Footprint Methodologies—Methodologies for the Assessment of Project Greenhouse Gas Emissions and Emission Variations, Version 11.3. 2023. Available online: https://www.eib.org/attachments/lucalli/eib_project_carbon_footprint_methodologies_2023_en.pdf (accessed on 27 April 2023).
- Ning, X.; Lin, R.; O’Shea, R.; Wall, D.; Deng, C.; Wu, B.; Murphy, J.D. Emerging bioelectrochemical technologies for biogas production and upgrading in cascading circular bioenergy systems. iScience 2021, 24, 102998. [Google Scholar] [CrossRef]
- International Energy Agency (IEA). The Future of Hydrogen—Seizing Today’s Opportunities. 2019. Available online: https://iea.blob.core.windows.net/assets/9e3a3493-b9a6-4b7d-b499-7ca48e357561/The_Future_of_Hydrogen.pdf (accessed on 30 March 2023).
- International Renewable Energy Agency (IRENA). Global Energy Transformation: A Roadmap to 2050. 2018. Available online: https://www.irena.org/-/media/Files/IRENA/Agency/Publication/2018/Apr/IRENA_Report_GET_2018.pdf (accessed on 20 March 2023).
- International Energy Agency (IEA). Hydrogen. 2021. Available online: https://www.iea.org/reports/hydrogen (accessed on 20 March 2023).
- de Kleijne, K.; de Coninck, H.; van Zelm, R.; Huijbregts, M.A.J.; Hanssen, S.V. The many greenhouse gas footprints of green hydrogen. Sustain. Energy Fuels 2022, 6, 4383. [Google Scholar] [CrossRef] [PubMed]
- Zappi, A.; Hernandez, R.; Holmes, W.E. A review of hydrogen production from anaerobic digestion. Int. J. Environ. Sci. Technol. 2021, 18, 4075–4090. [Google Scholar] [CrossRef]
- Rajesh Banu, J.; Rajesh Banu, J.; Kavitha, S.; Tyagi, V.K.; Gunasekaran, M.; Karthikeyan, O.P.; Kumar, G. Lignocellulosic biomass based biorefinery: A successful platform towards circular bioeconomy. Fuel 2021, 302, 121086. [Google Scholar] [CrossRef]
- Dahmen, N.; Lewandowski, I.; Zibek, S.; Weidtmann, A. Integrated lignocellulosic value chains in a growing bioeconomy: Status quo and perspectives. GCB Bioenergy 2019, 11, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Moraïs, S.; Himmel, M.E.; Bayer, E.A. New Paradigms for Engineering Plant Cell Wall Degrading Enzymes. In Direct Microbial Conversion of Biomass to Advanced Biofuels; Elsevier: Amsterdam, The Netherlands, 2015; Chapter 8; pp. 129–149. [Google Scholar] [CrossRef]
- Reducing enzyme costs, novel combinations, and advantages of enzymes could lead to improved cost-effective biofuels’ production. In Bioenergy: Biomass to Biofuels and Waste to Energy; Elsevier: Amsterdam, The Netherlands, 2020; pp. 573–579. [CrossRef]
- Biomass R&D Board. The Billion Ton Bioeconomy Initiative: Challenges & Opportunities. 2016. Available online: https://biomassboard.gov/sites/default/files/pdfs/the_bioeconomy_initiative.pdf (accessed on 13 May 2023).
- Rogers, J.N.; Stokes, B.; Dunn, J.; Cai, H.; Wu, M.; Haq, Z.; Baumes, H. An assessment of the potential products and economic and environmental impacts resulting from a billion ton bioeconomy. Biofuels Bioprod. 2017, 11, 110–128. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriclture Organization of the United Nations. Fertilizers by Nutrient. Available online: https://fenix.fao.org/faostat/internal/en/#data/RFN (accessed on 10 May 2023).
- Menegat, S.; Ledo, A.; Tirado, R. Greenhouse gas emissions from global production and use of nitrogen synthetic fertilisers in agriculture. Sci. Rep. 2022, 12, 14490. [Google Scholar] [CrossRef]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef]
- Hoben, J.P.; Gehl, R.J.; Millar, N.; Grace, P.R.; Robertson, G.P. Nonlinear nitrous oxide (N2O) response to nitrogen fertilizer in on-farm corn crops of the US Midwest. Glob. Chang. Biol. 2011, 17, 1140–1152. [Google Scholar] [CrossRef]
- Minamisawa, K. Mitigation of greenhouse gas emission by nitrogen-fixing bacteria. Biosci. Biotechnol. Biochem. 2023, 87, 7–12. [Google Scholar] [CrossRef]
- Guo, K.; Yang, J.; Yu, N.; Luo, L.; Wang, E. Biological nitrogen fixation in cereal crops: Progress, strategies, and perspectives. Plant Commun. 2023, 4, 100499. [Google Scholar] [CrossRef]
- European Commission; Directorate-General for Research and Innovation; Platt, R.; Bauen, A.; Reumerman, P.; Geier, C.; van Ree, R.; Gursel, I.V.; Garcia, L.; Behrens, M.; et al. EU Biorefinery Outlook to 2030: Studies on Support to Research and Innovation Policy in the Area of Bio-Based Products and Services. 2021. Available online: https://data.europa.eu/doi/10.2777/103465 (accessed on 21 December 2022).
- Bioenergy Technologies Office. Sustainable Aviation Fuel Grand Challenge. Available online: https://www.energy.gov/eere/bioenergy/sustainable-aviation-fuel-grand-challenge (accessed on 21 December 2022).
- Chandel, A.K.; Segato, F. (Eds.) Production of Top 12 Biochemicals Selected by USDOE from Renewable Resources, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2021; ISBN 978-012-823-654-3. [Google Scholar]
- Republic of Korea. 2050 Carbon Neutral Strategy of the Republic of Korea—Towards a Sustainable and Green Society. 2020. Available online: https://unfccc.int/sites/default/files/resource/LTS1_RKorea.pdf (accessed on 21 December 2022).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuchs, W.; Rachbauer, L.; Rittmann, S.K.-M.R.; Bochmann, G.; Ribitsch, D.; Steger, F. Eight Up-Coming Biotech Tools to Combat Climate Crisis. Microorganisms 2023, 11, 1514. https://doi.org/10.3390/microorganisms11061514
Fuchs W, Rachbauer L, Rittmann SK-MR, Bochmann G, Ribitsch D, Steger F. Eight Up-Coming Biotech Tools to Combat Climate Crisis. Microorganisms. 2023; 11(6):1514. https://doi.org/10.3390/microorganisms11061514
Chicago/Turabian StyleFuchs, Werner, Lydia Rachbauer, Simon K.-M. R. Rittmann, Günther Bochmann, Doris Ribitsch, and Franziska Steger. 2023. "Eight Up-Coming Biotech Tools to Combat Climate Crisis" Microorganisms 11, no. 6: 1514. https://doi.org/10.3390/microorganisms11061514