
Electroactive Bacteria in Natural Ecosystems and Their Applications in Microbial Fuel Cells for Bioremediation: A Review


Abstract
:1. Introduction
2. Electromicrobiomes in Natural Ecosystems
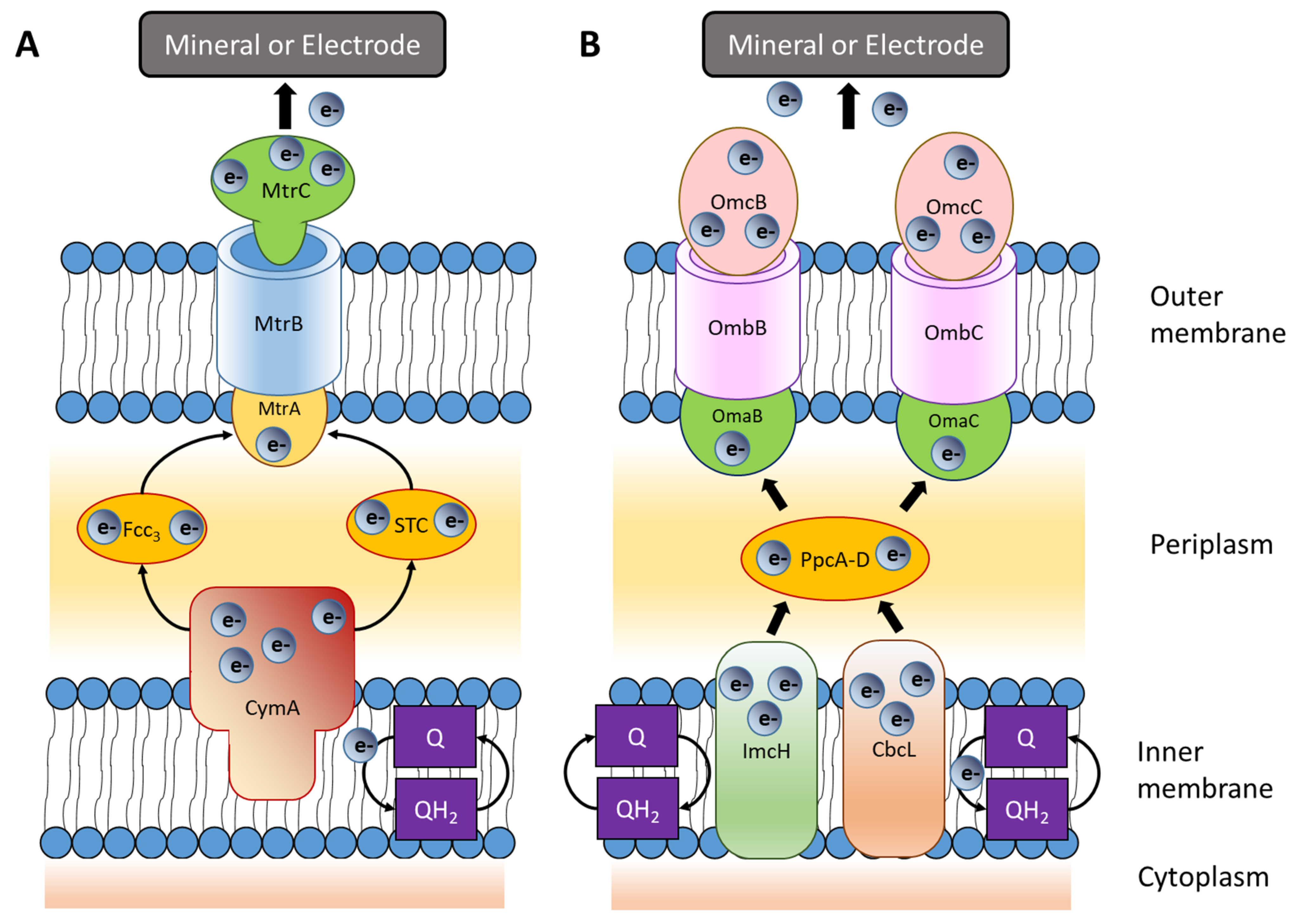
3. Extracellular Electron Transfer Mechanisms
4. Electrogenic Bacteria in Microbial Fuel Cells
5. MFCs for Wastewater Contaminant Removal
6. MFCs for Bioremediation of Persistent Organic Pollutants (POPs) and Heavy Metals
7. Plant-Microbial Fuel Cells and Constructed Wetland MFCs
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Potter, M.C. Electrical Effects Accompanying the Decomposition of Organic Compounds. Proc. R. Soc. London. Ser. B Contain. Pap. Biol. Character 1911, 84, 260–276. [Google Scholar] [CrossRef] [Green Version]
- Kouam Ida, T.; Mandal, B. Microbial Fuel Cell Design, Application and Performance: A Review. Mater. Today Proc. 2023, 76, 88–94. [Google Scholar] [CrossRef]
- Lovley, D.R. Electromicrobiology. Annu. Rev. Microbiol. 2012, 66, 391–409. [Google Scholar] [CrossRef]
- Allen, R.M.; Bennetto, H.P. Microbial Fuel-Cells. Appl. Biochem. Biotechnol. 1993, 39–40, 27–40. [Google Scholar] [CrossRef]
- Kim, B.H.; Kim, H.J.; Hyun, M.S.; Park, D.H. Direct Electrode Reaction of Fe(III)-Reducing Bacterium, Shewanella Putrefaciens. J. Microbiol. Biotechnol. 1999, 9, 127–131. [Google Scholar]
- Ikeda, S.; Takamatsu, Y.; Tsuchiya, M.; Suga, K.; Tanaka, Y.; Kouzuma, A.; Watanabe, K. Shewanella Oneidensis MR-1 as a Bacterial Platform for Electro-Biotechnology. Essays Biochem. 2021, 65, 355–364. [Google Scholar] [CrossRef]
- Bond, D.R.; Lovley, D.R. Electricity Production by Geobacter sulfurreducens Attached to Electrodes. Appl. Environ. Microbiol. 2003, 69, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.-E.E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [PubMed]
- Koch, C.; Harnisch, F. Is There a Specific Ecological Niche for Electroactive Microorganisms? ChemElectroChem 2016, 3, 1282–1295. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E. Electromicrobiology: The Ecophysiology of Phylogenetically Diverse Electroactive Microorganisms. Nat. Rev. Microbiol. 2022, 20, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Sharma, M.; Mohanakrishna, G.; Dominguez Benneton, X.; Strik, D.P.B.T.B.; Sarma, P.M.; Pant, D. An Overview on Emerging Bioelectrochemical Systems (BESs): Technology for Sustainable Electricity, Waste Remediation, Resource Recovery, Chemical Production and Beyond. Renew. Energy 2016, 98, 153–170. [Google Scholar] [CrossRef]
- Mier, A.A.; Olvera-Vargas, H.; Mejía-López, M.; Longoria, A.; Verea, L.; Sebastian, P.J.; Arias, D.M. A Review of Recent Advances in Electrode Materials for Emerging Bioelectrochemical Systems: From Biofilm-Bearing Anodes to Specialized Cathodes. Chemosphere 2021, 283, 131138. [Google Scholar] [CrossRef] [PubMed]
- Ancona, V.; Barra Caracciolo, A.; Borello, D.; Ferrara, V.; Grenni, P.; Pietrelli, A. Microbial Fuel Cell: An Energy Harvesting Technique for Environmental Remediation. Int. J. Environ. Impacts Manag. Mitig. Recover. 2020, 3, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Di Franca, M.L.; Matturro, B.; Crognale, S.; Zeppilli, M.; Armi, E.D.; Majone, M.; Papini, M.P.; Rossetti, S. Microbiome Composition and Dynamics of a Reductive/Oxidative Bioelectrochemical System for Perchloroethylene Removal: Effect of the Feeding Composition Reactor Configuration and Operating. Front. Microbiol. 2022, 13, 951911. [Google Scholar] [CrossRef]
- Abbas, S.Z.; Rafatullah, M. Recent Advances in Soil Microbial Fuel Cells for Soil Contaminants Remediation. Chemosphere 2021, 272, 129691. [Google Scholar] [CrossRef]
- Yaqoob, A.A.; Serrà, A.; Bhawani, S.A.; Ibrahim, M.N.M.; Khan, A.; Alorfi, H.S.; Asiri, A.M.; Hussein, M.A.; Khan, I.; Umar, K. Utilizing Biomass-Based Graphene Oxide–Polyaniline–Ag Electrodes in Microbial Fuel Cells to Boost Energy Generation and Heavy Metal Removal. Polymers 2022, 14, 845. [Google Scholar] [CrossRef]
- Deng, L.; Ngo, H.H.; Guo, W.; Chang, S.W.; Nguyen, D.D.; Pandey, A.; Varjani, S.; Hoang, N.B. Recent Advances in Circular Bioeconomy Based Clean Technologies for Sustainable Environment. J. Water Process Eng. 2022, 46, 102534. [Google Scholar] [CrossRef]
- Karuppiah, T.; Uthirakrishnan, U.; Sivakumar, S.V.; Authilingam, S.; Arun, J.; Sivaramakrishnan, R.; Pugazhendhi, A. Processing of Electroplating Industry Wastewater through Dual Chambered Microbial Fuel Cells (MFC) for Simultaneous Treatment of Wastewater and Green Fuel Production. Int. J. Hydrogen Energy 2022, 47, 37569–37576. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, J.; Shinde, S.; Wang, X.; Li, Y.; Dai, Y.; Ren, J.; Zhang, P.; Liu, X. Simultaneous Wastewater Treatment and Energy Harvesting in Microbial Fuel Cells: An Update on the Biocatalysts. RSC Adv. 2020, 10, 25874–25887. [Google Scholar] [CrossRef]
- James, A. Ceramic-Microbial Fuel Cell (C-MFC) for Waste Water Treatment: A Mini Review. Environ. Res. 2022, 210, 112963. [Google Scholar] [CrossRef]
- Borello, D.; Gagliardi, G.; Aimola, G.; Ancona, V.; Grenni, P.; Bagnuolo, G.; Garbini, G.L.; Rolando, L.; Barra Caracciolo, A. Use of Microbial Fuel Cells for Soil Remediation: A Preliminary Study on DDE. Int. J. Hydrogen Energy 2021, 46, 10131–10142. [Google Scholar] [CrossRef]
- Chabert, N.; Amin Ali, O.; Achouak, W. All Ecosystems Potentially Host Electrogenic Bacteria. Bioelectrochemistry 2015, 106, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Miceli, J.F.; Parameswaran, P.; Kang, D.-W.; Krajmalnik-Brown, R.; Torres, C.I. Enrichment and Analysis of Anode-Respiring Bacteria from Diverse Anaerobic Inocula. Environ. Sci. Technol. 2012, 46, 10349–10355. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pabello, P.U.; Sedov, S.; Solleiro-Rebolledo, E.; Solé, J.; Pi-Puig, T.; Alcántara-Hernández, R.J.; Lebedeva, M.; Shishkov, V.; Villalobos, C. Rock Varnish in La Proveedora/Sonora in the Context of Desert Geobiological Processes and Landscape Evolution. J. South Am. Earth Sci. 2021, 105, 102959. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takaki, Y.; Kashima, H.; Tsuda, M.; Tanizaki, A.; Nakamura, R.; Takai, K. In Situ Electrosynthetic Bacterial Growth Using Electricity Generated by a Deep-Sea Hydrothermal Vent. ISME J. 2023, 17, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Yan, Y.; Nie, Y.; Lu, A.; Wu, X.; Li, Y.; Wang, C.; Ding, H. Natural Extracellular Electron Transfer Between Semiconducting Minerals and Electroactive Bacterial Communities Occurred on the Rock Varnish. Front. Microbiol. 2019, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, Z.; Ma, J.; Yun, H.; Qi, M.; Ma, X.; Wang, H.; Wang, A.; Liang, B. Enhanced Bioelectroremediation of a Complexly Contaminated River Sediment through Stimulating Electroactive Degraders with Methanol Supply. J. Hazard. Mater. 2018, 349, 168–176. [Google Scholar] [CrossRef]
- Shi, K.; Liang, B.; Guo, Q.; Zhao, Y.; Sharif, H.M.A.; Li, Z.; Chen, E.; Wang, A. Accelerated Bioremediation of a Complexly Contaminated River Sediment through ZVI-Electrode Combined Stimulation. J. Hazard. Mater. 2021, 413, 125392. [Google Scholar] [CrossRef]
- Lu, X.; von Haxthausen, K.A.; Brock, A.L.; Trapp, S. Turnover of Lake Sediments Treated with Sediment Microbial Fuel Cells: A Long-Term Study in a Eutrophic Lake. Sci. Total Environ. 2021, 796, 148880. [Google Scholar] [CrossRef]
- Rimboud, M.; Etcheverry, L.; Barakat, M.; Achouak, W.; Bergel, A.; Délia, M.-L. Hypersaline Microbial Fuel Cell Equipped with an Oxygen-Reducing Microbial Cathode. Bioresour. Technol. 2021, 337, 125448. [Google Scholar] [CrossRef]
- Yang, X.; Chen, S. Microorganisms in Sediment Microbial Fuel Cells: Ecological Niche, Microbial Response, and Environmental Function. Sci. Total Environ. 2021, 756, 144145. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, E.; Bouillon, S.; Dittmar, T.; Marchand, C. Organic Carbon Dynamics in Mangrove Ecosystems: A Review. Aquat. Bot. 2008, 89, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Szydlowski, L.; Lan, T.C.T.; Shibata, N.; Goryanin, I. Metabolic Engineering of a Novel Strain of Electrogenic Bacterium Arcobacter Butzleri to Create a Platform for Single Analyte Detection Using a Microbial Fuel Cell. Enzyme Microb. Technol. 2020, 139, 109564. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, Y.; Punamalai, G.; Kandasamy, S.; Kasinathan, K. Influence of Long-Term Organic and Conventional Fertilization on Bacterial Communities Involved in Bioelectricity Production from Paddy Field-Microbial Fuel Cells. Arch. Microbiol. 2020, 202, 2279–2289. [Google Scholar] [CrossRef] [PubMed]
- Gustave, W.; Yuan, Z.-F.; Sekar, R.; Toppin, V.; Liu, J.-Y.; Ren, Y.-X.; Zhang, J.; Chen, Z. Relic DNA Does Not Obscure the Microbial Community of Paddy Soil Microbial Fuel Cells. Res. Microbiol. 2019, 170, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Yang, C.; Li, Z.; Luo, H.; Feng, S.; Zhao, Y.; Lu, L. Plant Endophytic Bacteria: A Potential Resource Pool of Electroactive Micro-Organisms. J. Appl. Microbiol. 2022, 132, 2054–2066. [Google Scholar] [CrossRef]
- Tahernia, M.; Plotkin-Kaye, E.; Mohammadifar, M.; Gao, Y.; Oefelein, M.R.; Cook, L.C.; Choi, S. Characterization of Electrogenic Gut Bacteria. ACS Omega 2020, 5, 29439–29446. [Google Scholar] [CrossRef]
- Harrison, A.P. Acidiphilium Cryptum Gen. Nov., Sp. Nov., Heterotrophic Bacterium From Acidic Mineral Environments. Int. J. Syst. Bacteriol. 1981, 31, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Borole, A.P.; O’Neill, H.; Tsouris, C.; Cesar, S. A Microbial Fuel Cell Operating at Low PH Using the Acidophile Acidiphilium Cryptum. Biotechnol. Lett. 2008, 30, 1367–1372. [Google Scholar] [CrossRef]
- Yang, Y.; Wan, M.; Shi, W.; Peng, H.; Qiu, G.; Zhou, J.; Liu, X. Bacterial Diversity and Community Structure in Acid Mine Drainage from Dabaoshan Mine, China. Aquat. Microb. Ecol. 2007, 47, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Erable, B.; Vandecandelaere, I.; Faimali, M.; Delia, M.-L.; Etcheverry, L.; Vandamme, P.; Bergel, A. Marine Aerobic Biofilm as Biocathode Catalyst. Bioelectrochemistry 2010, 78, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yu, P.; Zeng, C.; Ding, H.; Li, Y.; Wang, C.; Lu, A. Enhanced Alcaligenes Faecalis Denitrification Rate with Electrodes as the Electron Donor. Appl. Environ. Microbiol. 2015, 81, 5387–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooda, R.; Bhardwaj, N.K.; Singh, P. Screening and Identification of Ligninolytic Bacteria for the Treatment of Pulp and Paper Mill Effluent. Water Air Soil Pollut. 2015, 226, 305. [Google Scholar] [CrossRef]
- Sydow, A.; Krieg, T.; Mayer, F.; Schrader, J.; Holtmann, D. Electroactive Bacteria—Molecular Mechanisms and Genetic Tools. Appl. Microbiol. Biotechnol. 2014, 98, 8481–8495. [Google Scholar] [CrossRef]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef]
- Sacco, N.J.; Bonetto, M.C.; Cortón, E. Isolation and characterization of a novel electrogenic bacterium, Dietzia Sp. RNV-4. PLoS ONE 2017, 12, e0169955. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, S.; Das, S.; Hossain, M.T.; Islam, A.; Miah, M.R.; Awual, M.R. Sustainable Approach for wastewater treatment using microbial fuel cells and green energy generation—A Comprehensive Review. J. Mol. Liq. 2021, 344, 117795. [Google Scholar] [CrossRef]
- Nelson, Y.M.; Lion, L.W.; Ghiorse, W.C.; Shuler, M.L. Production of Biogenic Mn Oxides by Leptothrix Discophora SS-1 in a Chemically Defined Growth Medium and Evaluation of Their Pb Adsorption Characteristics. Appl. Environ. Microbiol. 1999, 65, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Wasserfallen, A.; Nölling, J.; Pfister, P.; Reeve, J.; Conway de Macario, E. Phylogenetic Analysis of 18 Thermophilic Methanobacterium Isolates Supports the Proposals to Create a New Genus, Methanothermobacter Gen. Nov., and to Reclassify Several Isolates in Three Species, Methanothermobacter Thermautotrophicus Comb. Nov., Methano. Int. J. Syst. Evol. Microbiol. 2000, 50, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Bennur, T.; Kumar, A.R.; Zinjarde, S.; Javdekar, V. Nocardiopsis Species: Incidence, Ecological Roles and Adaptations. Microbiol. Res. 2015, 174, 33–47. [Google Scholar] [CrossRef]
- Zuo, Y.; Xing, D.; Regan, J.M.; Logan, B.E. Isolation of the Exoelectrogenic Bacterium Ochrobactrum Anthropi YZ-1 by Using a U-Tube Microbial Fuel Cell. Appl. Environ. Microbiol. 2008, 74, 3130–3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, N.; Zhou, Z.; Karimi, I.A. Metabolic Processes of Methanococcus Maripaludis and Potential Applications. Microb. Cell Fact. 2016, 15, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkateswaran, K.; Moser, D.P.; Dollhopf, M.E.; Lies, D.P.; Saffarini, D.A.; MacGregor, B.J.; Ringelberg, D.B.; White, D.C.; Nishijima, M.; Sano, H.; et al. Polyphasic Taxonomy of the Genus Shewanella and Description of Shewanella oneidensis Sp. Nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 705–724. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Zhu, J.N.; Zhai, Z.H.; Zhang, Q. Endophytic Bacterial diversity in roots of Phragmites australis in constructed Beijing Cuihu Wetland (China). FEMS Microbiol. Lett. 2010, 309, 84–93. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Ye, Z.; Yang, D.; Yan, J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.; Fang, Z.; Jing, Y. Characterization of Endophytic Rahnella Sp. JN6 from Polygonum Pubescens and Its Potential in Promoting Growth and Cd, Pb, Zn Uptake by Brassica Napus. Chemosphere 2013, 90, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Taulé, C.; Mareque, C.; Barlocco, C.; Hackembruch, F.; Reis, V.M.; Sicardi, M.; Battistoni, F. The Contribution of Nitrogen Fixation to Sugarcane (Saccharum officinarum L.), and the Identification and Characterization of Part of the Associated Diazotrophic Bacterial Community. Plant Soil 2012, 356, 35–49. [Google Scholar] [CrossRef]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive Microorganisms in Bioelectrochemical Systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular Electron Transfer Mechanisms between Microorganisms and Minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Sime-Ngando, T.; Bertrand, J.-C.; Bogusz, D.; Brugère, J.-F.; Franche, C.; Fardeau, M.-L.; Froussart, E.; Geiger, A.; Goñi-Urriza, M.S.; Ollivier, B.; et al. The Evolution of Living Beings Started with Prokaryotes and in Interaction with Prokaryotes. In Prokaryotes and Evolution; Springer International Publishing: Cham, Switzerland, 2018; pp. 241–338. [Google Scholar]
- Edel, M.; Horn, H.; Gescher, J. Biofilm Systems as Tools in Biotechnological Production. Appl. Microbiol. Biotechnol. 2019, 103, 5095–5103. [Google Scholar] [CrossRef]
- Paquete, C.M.; Rosenbaum, M.A.; Bañeras, L.; Rotaru, A.-E.E.; Puig, S. Let’s Chat: Communication between Electroactive Microorganisms. Bioresour. Technol. 2022, 347, 126705. [Google Scholar] [CrossRef]
- Bird, L.J.; Kundu, B.B.; Tschirhart, T.; Corts, A.D.; Su, L.; Gralnick, J.A.; Ajo-Franklin, C.M.; Glaven, S.M. Engineering Wired Life: Synthetic Biology for Electroactive Bacteria. ACS Synth. Biol. 2021, 10, 2808–2823. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.R. Microbial Reduction of Metals and Radionuclides. FEMS Microbiol. Rev. 2003, 27, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Dissimilatory Fe(III) and Mn(IV) Reduction. Microbiol. Rev. 1991, 55, 259–287. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D. Dissimilatory Fe(III)- and Mn(IV)-Reducing Prokaryotes. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 287–308. [Google Scholar]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms Pumping Iron: Anaerobic Microbial Iron Oxidation and Reduction. Nat. Rev. Microbiol. 2006, 4, 752–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, C.R.; Nealson, K.H. Bacterial Manganese Reduction and Growth with Manganese Oxide as the Sole Electron Acceptor. Science 1988, 240, 1319–1321. [Google Scholar] [CrossRef]
- Hartshorne, R.S.; Reardon, C.L.; Ross, D.; Nuester, J.; Clarke, T.A.; Gates, A.J.; Mills, P.C.; Fredrickson, J.K.; Zachara, J.M.; Shi, L.; et al. Characterization of an Electron Conduit between Bacteria and the Extracellular Environment. Proc. Natl. Acad. Sci. USA 2009, 106, 22169–22174. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.J.; Butt, J.N.; Fredrickson, J.K.; Zachara, J.M.; Shi, L.; Edwards, M.J.; White, G.; Baiden, N.; Gates, A.J.; Marritt, S.J.; et al. The ‘Porin-Cytochrome’ Model for Microbe-to-Mineral Electron Transfer. Mol. Microbiol. 2012, 85, 201–212. [Google Scholar] [CrossRef] [Green Version]
- White, G.F.; Shi, Z.; Shi, L.; Wang, Z.; Dohnalkova, A.C.; Marshall, M.J.; Fredrickson, J.K.; Zachara, J.M.; Butt, J.N.; Richardson, D.J.; et al. Rapid Electron Exchange between Surface-Exposed Bacterial Cytochromes and Fe(III) Minerals. Proc. Natl. Acad. Sci. USA 2013, 110, 6346–6351. [Google Scholar] [CrossRef] [Green Version]
- Levar, C.E.; Chan, C.H.; Mehta-Kolte, M.G.; Bond, D.R. An Inner Membrane Cytochrome Required Only for Reduction of High Redox Potential Extracellular Electron Acceptors. MBio 2014, 5, e02034-14. [Google Scholar] [CrossRef] [Green Version]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically Conductive Bacterial Nanowires Produced by Shewanella oneidensis Strain MR-1 and Other Microorganisms. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef] [Green Version]
- Pirbadian, S.; Barchinger, S.E.; Leung, K.M.; Byun, H.S.; Jangir, Y.; Bouhenni, R.A.; Reed, S.B.; Romine, M.F.; Saffarini, D.A.; Shi, L.; et al. Shewanella oneidensis MR-1 Nanowires Are Outer Membrane and Periplasmic Extensions of the Extracellular Electron Transport Components. Proc. Natl. Acad. Sci. USA 2014, 111, 12883–12888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacharoff, L.; Chan, C.H.; Bond, D.R. Reduction of Low Potential Electron Acceptors Requires the CbcL Inner Membrane Cytochrome of Geobacter sulfurreducens. Bioelectrochemistry 2016, 107, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct Exchange of Electrons within Aggregates of an Evolved Syntrophic Coculture of Anaerobic Bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotaru, A.-E.; Shrestha, P.M.; Liu, F.; Markovaite, B.; Chen, S.; Nevin, K.P.; Lovley, D.R. Direct Interspecies Electron Transfer between Geobacter Metallireducens and Methanosarcina Barkeri. Appl. Environ. Microbiol. 2014, 80, 4599–4605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Wang, A.; Ren, H.-Y. Improved 4-Chlorophenol Dechlorination at Biocathode in Bioelectrochemical System Using Optimized Modular Cathode Design with Composite Stainless Steel and Carbon-Based Materials. Bioresour. Technol. 2014, 166, 252–258. [Google Scholar] [CrossRef]
- Hemdan, B.A.; El-Taweel, G.E.; Naha, S.; Goswami, P. Bacterial Community Structure of Electrogenic Biofilm Developed on Modified Graphite Anode in Microbial Fuel Cell. Sci. Rep. 2023, 13, 1255. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, J.; Zhang, Y.; Guo, Q.; Hu, T.; Xiao, H.; Lu, W.; Jia, J. Progress on Anodic Modification Materials and Future Development Directions in Microbial Fuel Cells. J. Power Sources 2023, 556, 232486. [Google Scholar] [CrossRef]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial Fuel Cells: Methodology and Technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef]
- Xie, Q.; Lu, Y.; Tang, L.; Zeng, G.; Yang, Z.; Fan, C.; Wang, J.; Atashgahi, S. The Mechanism and Application of Bidirectional Extracellular Electron Transport in the Field of Energy and Environment. Crit. Rev. Environ. Sci. Technol. 2021, 51, 1924–1969. [Google Scholar] [CrossRef]
- Ahn, Y.-H. Sustainable Nitrogen Elimination Biotechnologies: A Review. Process Biochem. 2006, 41, 1709–1721. [Google Scholar] [CrossRef]
- Tsipa, A.; Varnava, C.K.; Grenni, P.; Ferrara, V.; Pietrelli, A. Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling. Processes 2021, 9, 1038. [Google Scholar] [CrossRef]
- Liu, H.; Ramnarayanan, R.; Logan, B.E. Production of Electricity during Wastewater Treatment Using a Single Chamber Microbial Fuel Cell. Environ. Sci. Technol. 2004, 38, 2281–2285. [Google Scholar] [CrossRef]
- Ye, Y.; Ngo, H.H.; Guo, W.; Chang, S.W.; Nguyen, D.D.; Liu, Y.; Ni, B.; Zhang, X. Microbial Fuel Cell for Nutrient Recovery and Electricity Generation from Municipal Wastewater under Different Ammonium Concentrations. Bioresour. Technol. 2019, 292, 121992. [Google Scholar] [CrossRef]
- Sanjay, S.; Udayashankara, T.H. Dairy Wastewater Treatment with Bio-Electricity Generation Using Dual Chambered Membrane-Less Microbial Fuel Cell. Mater. Today Proc. 2021, 35, 308–311. [Google Scholar] [CrossRef]
- Vélez-Pérez, L.S.; Ramirez-Nava, J.; Hernández-Flores, G.; Talavera-Mendoza, O.; Escamilla-Alvarado, C.; Poggi-Varaldo, H.M.; Solorza-Feria, O.; López-Díaz, J.A. Industrial Acid Mine Drainage and Municipal Wastewater Co-Treatment by Dual-Chamber Microbial Fuel Cells. Int. J. Hydrogen Energy 2020, 45, 13757–13766. [Google Scholar] [CrossRef]
- Tacas, A.C.J.; Tsai, P.-W.; Tayo, L.L.; Hsueh, C.-C.; Sun, S.-Y.; Chen, B.-Y. Degradation and Biotoxicity of Azo Dyes Using Indigenous Bacteria-Acclimated Microbial Fuel Cells (MFCs). Process Biochem. 2021, 102, 59–71. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Shahid, M.K.; Zhen, G.; Kumar, G.; Shin, H.-S.; Choi, Y.-G.; Kim, S.-H. A Comprehensive Overview on Electro-Active Biofilms, Role of Exo-Electrogens and Their Microbial Niches in Microbial Fuel Cells (MFCs). Chemosphere 2017, 178, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, G.; Zhang, H.; Wen, H.; Li, W. Effects of Biofilm Transfer and Electron Mediators Transfer on Klebsiella Quasipneumoniae Sp. 203 Electricity Generation Performance in MFCs. Biotechnol. Biofuels 2020, 13, 162. [Google Scholar] [CrossRef]
- Visca, A.; Barra Caracciolo, A.; Grenni, P.; Rolando, L.; Mariani, L.; Rauseo, J.; Spataro, F.; Monostory, K.; Sperlagh, B.; Patrolecco, L. Legacy and Emerging Pollutants in an Urban River Stretch and Effects on the Bacterioplankton Community. Water 2021, 13, 3402. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Li, D. Cometabolic Degradation of Chloramphenicol via a Meta-Cleavage Pathway in a Microbial Fuel Cell and Its Microbial Community. Bioresour. Technol. 2017, 229, 104–110. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Ma, J.; Zhao, F. Rapid Degradation of Sulphamethoxazole and the Further Transformation of 3-Amino-5-Methylisoxazole in a Microbial Fuel Cell. Water Res. 2016, 88, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Jiang, J.; Zhang, S.; Wang, X.; Guo, X.; Li, F. Enzymatic Response and Antibiotic Resistance Gene Regulation by Microbial Fuel Cells to Resist Sulfamethoxazole. Chemosphere 2023, 325, 138410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, R.; Du, C.; Dong, S.; Sun, J. Effects of Continuous Sulfamonomethoxine Shock on the Power Generation Performance and Microbial Community Structure of MFCs under Seasonal Temperature Variation. Biochem. Eng. J. 2021, 167, 107909. [Google Scholar] [CrossRef]
- Tsekouras, G.J.; Deligianni, P.M.; Kanellos, F.D.; Kontargyri, V.T.; Kontaxis, P.A.; Manousakis, N.M.; Elias, C.N. Microbial Fuel Cell for Wastewater Treatment as Power Plant in Smart Grids: Utopia or Reality? Front. Energy Res. 2022, 10, 370. [Google Scholar] [CrossRef]
- Malik, S.; Kishore, S.; Dhasmana, A.; Kumari, P.; Mitra, T.; Chaudhary, V.; Kumari, R.; Bora, J.; Ranjan, A.; Minkina, T.; et al. A Perspective Review on Microbial Fuel Cells in Treatment and Product Recovery from Wastewater. Water 2023, 15, 316. [Google Scholar] [CrossRef]
- Liang, P.; Duan, R.; Jiang, Y.; Zhang, X.; Qiu, Y.; Huang, X. One-Year Operation of 1000-L Modularized Microbial Fuel Cell for Municipal Wastewater Treatment. Water Res. 2018, 141, 1–8. [Google Scholar] [CrossRef]
- Blatter, M.; Delabays, L.; Furrer, C.; Huguenin, G.; Cachelin, C.P.; Fischer, F. Stretched 1000-L Microbial Fuel Cell. J. Power Sources 2021, 483, 229130. [Google Scholar] [CrossRef]
- Goto, Y.; Yoshida, N.; Umeyama, Y.; Yamada, T.; Tero, R.; Hiraishi, A. Enhancement of Electricity Production by Graphene Oxide in Soil Microbial Fuel Cells and Plant Microbial Fuel Cells. Front. Bioeng. Biotechnol. 2015, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Ancona, V.; Rascio, I.; Aimola, G.; Campanale, C.; Grenni, P.; di Lenola, M.; Garbini, G.L.; Uricchio, V.F.; Barra Caracciolo, A. Poplar-Assisted Bioremediation for Recovering a PCB and Heavy-Metal-Contaminated Area. Agriculture 2021, 11, 689. [Google Scholar] [CrossRef]
- Yang, W.; He, W.; Zhang, F.; Hickner, M.A.; Logan, B.E. Single-Step Fabrication Using a Phase Inversion Method of Poly(Vinylidene Fluoride) (PVDF) Activated Carbon Air Cathodes for Microbial Fuel Cells. Environ. Sci. Technol. Lett. 2014, 1, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Jiang, Q.; Liu, J.; Shao, Y.; Feng, Y. Enhanced Electricity Generation and Water Pressure Tolerance Using Carbon Black-Based Sintered Filtration Air-Cathodes in Microbial Fuel Cells. Chem. Eng. J. 2019, 369, 652–659. [Google Scholar] [CrossRef]
- Kuo, J.; Liu, D.; Wang, S.-H.H.; Lin, C.-H.H. Dynamic Changes in Soil Microbial Communities with Glucose Enrichment in Sediment Microbial Fuel Cells. Indian J. Microbiol. 2021, 61, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Magotra, V.K.; Kumar, S.; Kang, T.W.; Inamdar, A.I.; Aqueel, A.T.; Im, H.; Ghodake, G.; Shinde, S.; Waghmode, D.P.; Jeon, H.C. Compost Soil Microbial Fuel Cell to Generate Power Using Urea as Fuel. Sci. Rep. 2020, 10, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magotra, V.K.; Kang, T.W.; Aqueel Ahmed, A.T.; Inamdar, A.I.; Im, H.; Ghodake, G.; Choubey, R.K.; Kumar, V.; Kumar, S. Effect of Gold Nanoparticles Laced Anode on the Bio-Electro-Catalytic Activity and Power Generation Ability of Compost Based Microbial Fuel Cell as a Coin Cell Sized Device. Biomass Bioenergy 2021, 152, 106200. [Google Scholar] [CrossRef]
- Zhang, D.; Ge, Y.; Wang, W. Study of a Terrestrial Microbial Fuel Cell and the Effects of Its Power Generation Performance by Environmental Factors. In Proceedings of the 2013 International Conference on Advanced Mechatronic Systems, Luoyang, China, 25–27 September 2013; pp. 445–448. [Google Scholar]
- Aimola, G.; Gagliardi, G.; Pietrelli, A.; Ancona, V.; Barra Cracciolo, A.; Borello, D.; Ferrara, V.; Grenni, P. Environmental remediation and possible use of terrestrial microbial fuel cells. Disaster Manag. Hum. Health Risk VII WIT Trans. Built Environ. 2021, 207, 121–133. [Google Scholar]
- Yu, B.; Tian, J.; Feng, L. Remediation of PAH Polluted Soils Using a Soil Microbial Fuel Cell: Influence of Electrode Interval and Role of Microbial Community. J. Hazard. Mater. 2017, 336, 110–118. [Google Scholar] [CrossRef]
- Hao, D.-C.; Li, X.-J.; Xiao, P.-G.; Wang, L.-F. The Utility of Electrochemical Systems in Microbial Degradation of Polycyclic Aromatic Hydrocarbons: Discourse, Diversity and Design. Front. Microbiol. 2020, 11, 557400. [Google Scholar] [CrossRef]
- Mohanakrishna, G.; Abu-Reesh, I.M.; Pant, D. Enhanced Bioelectrochemical Treatment of Petroleum Refinery Wastewater with Labaneh Whey as Co-Substrate. Sci. Rep. 2020, 10, 19665. [Google Scholar] [CrossRef]
- Cao, X.; Song, H.; Yu, C.; Li, X. Simultaneous Degradation of Toxic Refractory Organic Pesticide and Bioelectricity Generation Using a Soil Microbial Fuel Cell. Bioresour. Technol. 2015, 189, 87–93. [Google Scholar] [CrossRef]
- Li, X.; Zheng, R.; Zhang, X.; Liu, Z.; Zhu, R.; Zhang, X.; Gao, D. A Novel Exoelectrogen from Microbial Fuel Cell: Bioremediation of Marine Petroleum Hydrocarbon Pollutants. J. Environ. Manag. 2019, 235, 70–76. [Google Scholar] [CrossRef]
- Wang, H.; Song, H.; Yu, R.; Cao, X.; Fang, Z.; Li, X. New Process for Copper Migration by Bioelectricity Generation in Soil Microbial Fuel Cells. Environ. Sci. Pollut. Res. 2016, 23, 13147–13154. [Google Scholar] [CrossRef] [PubMed]
- Jaswal, V.; Rani, G.; Yogalakshmi, K.N. Photosynthetic Microbial Fuel Cells: From Fundamental to Potential Applications. In Bioelectrochemical Systems; Springer: Singapore, 2020; pp. 1–19. [Google Scholar]
- Rosenbaum, M.; He, Z.; Angenent, L.T. Light Energy to Bioelectricity: Photosynthetic Microbial Fuel Cells. Curr. Opin. Biotechnol. 2010, 21, 259–264. [Google Scholar] [CrossRef]
- Apollon, W.; Luna-Maldonado, A.I.; Kamaraj, S.-K.; Vidales-Contreras, J.A.; Rodríguez-Fuentes, H.; Gómez-Leyva, J.F.; Aranda-Ruíz, J. Progress and Recent Trends in Photosynthetic Assisted Microbial Fuel Cells: A Review. Biomass Bioenergy 2021, 148, 106028. [Google Scholar] [CrossRef]
- Ahirwar, A.; Das, S.; Das, S.; Yang, Y.-H.; Bhatia, S.K.; Vinayak, V.; Ghangrekar, M.M. Photosynthetic Microbial Fuel Cell for Bioenergy and Valuable Production: A Review of Circular Bio-Economy Approach. Algal Res. 2023, 70, 102973. [Google Scholar] [CrossRef]
- Kuleshova, T.; Rao, A.; Bhadra, S.; Garlapati, V.K.; Sharma, S.; Kaushik, A.; Goswami, P.; Sreekirshnan, T.R.; Sevda, S. Plant Microbial Fuel Cells as an Innovative, Versatile Agro-Technology for Green Energy Generation Combined with Wastewater Treatment and Food Production. Biomass Bioenergy 2022, 167, 106629. [Google Scholar] [CrossRef]
- Li, B.; Xu, D.; Feng, L.; Liu, Y.; Zhang, L. Advances and Prospects on the Aquatic Plant Coupled with Sediment Microbial Fuel Cell System. Environ. Pollut. 2022, 297, 118771. [Google Scholar] [CrossRef]
- Helder, M.; Strik, D.P.B.T.B.; Hamelers, H.V.M.; Kuhn, A.J.; Blok, C.; Buisman, C.J.N. Concurrent Bio-Electricity and Biomass Production in Three Plant-Microbial Fuel Cells Using Spartina Anglica, Arundinella Anomala and Arundo Donax. Bioresour. Technol. 2010, 101, 3541–3547. [Google Scholar] [CrossRef] [PubMed]
- Wetser, K.; Dieleman, K.; Buisman, C.; Strik, D. Electricity from Wetlands: Tubular Plant Microbial Fuels with Silicone Gas-Diffusion Biocathodes. Appl. Energy 2017, 185, 642–649. [Google Scholar] [CrossRef] [Green Version]
- De Schamphelaire, L.; Bossche, L.V.D.; Dang, H.S.; Höfte, M.; Boon, N.; Rabaey, K.; Verstraete, W. Microbial Fuel Cells Generating Electricity from Rhizodeposits of Rice Plants. Environ. Sci. Technol. 2008, 42, 3053–3058. [Google Scholar] [CrossRef]
- Tongphanpharn, N.; Chou, C.-H.; Guan, C.-Y.; Yu, C.-P. Plant Microbial Fuel Cells with Oryza rufipogon and Typha orientalis for Remediation of Cadmium Contaminated Soil. Environ. Technol. Innov. 2021, 24, 102030. [Google Scholar] [CrossRef]
- Xu, P.; Xiao, E.; Zeng, L.; He, F.; Wu, Z. Enhanced Degradation of Pyrene and Phenanthrene in Sediments through Synergistic Interactions between Microbial Fuel Cells and Submerged Macrophyte Vallisneria spiralis. J. Soils Sediments 2019, 19, 2634–2649. [Google Scholar] [CrossRef]
- Guan, C.-Y.; Tseng, Y.-H.; Tsang, D.C.W.; Hu, A.; Yu, C.-P. Wetland Plant Microbial Fuel Cells for Remediation of Hexavalent Chromium Contaminated Soils and Electricity Production. J. Hazard. Mater. 2019, 365, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.-Y.; Hu, A.; Yu, C.-P. Stratified Chemical and Microbial Characteristics between Anode and Cathode after Long-Term Operation of Plant Microbial Fuel Cells for Remediation of Metal Contaminated Soils. Sci. Total Environ. 2019, 670, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Jiang, H.; Cai, H.; Zhou, Y.; Krumholz, L.R. Complex Interactions Between the Macrophyte Acorus Calamus and Microbial Fuel Cells During Pyrene and Benzo[a]Pyrene Degradation in Sediments. Sci. Rep. 2015, 5, 10709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Qiu, D.; Lu, F.; Wang, Y.; Wang, Z.; Feng, X.; Pyo, S.-H. Acorus calamus L. Constructed Wetland-Microbial Fuel Cell for Cr(VI)-Containing Wastewater Treatment and Bioelectricity Production. J. Environ. Chem. Eng. 2022, 10, 107801. [Google Scholar] [CrossRef]
- Lin, C.-W.; Alfanti, L.K.; Cheng, Y.-S.; Liu, S.-H. Enhancing Bioelectricity Production and Copper Remediation in Constructed Single-Medium Plant Sediment Microbial Fuel Cells. Desalination 2022, 542, 116079. [Google Scholar] [CrossRef]
- Habibul, N.; Hu, Y.; Wang, Y.-K.; Chen, W.; Yu, H.-Q.; Sheng, G.-P. Bioelectrochemical Chromium(VI) Removal in Plant-Microbial Fuel Cells. Environ. Sci. Technol. 2016, 50, 3882–3889. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency. Guiding Principles for Constructed Treatment Wetlands: Providing for Water Quality and Wildlife Habitat. 2000. Available online: www.epa.gov/owow/wetlands/%0Aconstructed/guide.html (accessed on 7 April 2023).
- Kataki, S.; Chatterjee, S.; Vairale, M.G.; Dwivedi, S.K.; Gupta, D.K. Constructed Wetland, an Eco-Technology for Wastewater Treatment: A Review on Types of Wastewater Treated and Components of the Technology (Macrophyte, Biolfilm and Substrate). J. Environ. Manag. 2021, 283, 111986. [Google Scholar] [CrossRef]
- Ravikumar, Y.; Yun, J.; Zhang, G.; Zabed, H.M.; Qi, X. A Review on Constructed Wetlands-Based Removal of Pharmaceutical Contaminants Derived from Non-Point Source Pollution. Environ. Technol. Innov. 2022, 26, 102504. [Google Scholar] [CrossRef]
- Gupta, S.; Mittal, Y.; Tamta, P.; Srivastava, P.; Yadav, A.K. Textile Wastewater Treatment Using Microbial Fuel Cell and Coupled Technology: A Green Approach for Detoxification and Bioelectricity Generation. In Integrated Microbial Fuel Cells for Wastewater Treatment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 73–92. [Google Scholar]
- Gupta, S.; Srivastava, P.; Patil, S.A.; Yadav, A.K. A Comprehensive Review on Emerging Constructed Wetland Coupled Microbial Fuel Cell Technology: Potential Applications and Challenges. Bioresour. Technol. 2021, 320, 124376. [Google Scholar] [CrossRef]
- Gupta, S.; Srivastava, P.; Yadav, A.K. Integration of Microbial Fuel Cell into Constructed Wetlands: Effects, Applications, and Future Outlook. In Integrated Microbial Fuel Cells for Wastewater Treatment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 273–293. [Google Scholar]
- Wang, J.; Song, X.; Wang, Y.; Abayneh, B.; Li, Y.; Yan, D.; Bai, J. Nitrate Removal and Bioenergy Production in Constructed Wetland Coupled with Microbial Fuel Cell: Establishment of Electrochemically Active Bacteria Community on Anode. Bioresour. Technol. 2016, 221, 358–365. [Google Scholar] [CrossRef]
- Yadav, A.K.; Dash, P.; Mohanty, A.; Abbassi, R.; Mishra, B.K. Performance Assessment of Innovative Constructed Wetland-Microbial Fuel Cell for Electricity Production and Dye Removal. Ecol. Eng. 2012, 47, 126–131. [Google Scholar] [CrossRef]
- Lu, L.; Xing, D.; Ren, Z.J. Microbial Community Structure Accompanied with Electricity Production in a Constructed Wetland Plant Microbial Fuel Cell. Bioresour. Technol. 2015, 195, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Cao, F.; Kong, Q.; Zhou, L.; Yuan, Q.; Zhu, Y.; Wang, Q.; Du, Y.; Wang, Z. Electricity Production and Evolution of Microbial Community in the Constructed Wetland-Microbial Fuel Cell. Chem. Eng. J. 2018, 339, 479–486. [Google Scholar] [CrossRef]
- Srivastava, P.; Abbassi, R.; Garaniya, V.; Lewis, T.; Yadav, A.K. Performance of Pilot-Scale Horizontal Subsurface Flow Constructed Wetland Coupled with a Microbial Fuel Cell for Treating Wastewater. J. Water Process Eng. 2020, 33, 100994. [Google Scholar] [CrossRef]
- Srivastava, P.; Yadav, A.K.; Garaniya, V.; Lewis, T.; Abbassi, R.; Khan, S.J. Electrode Dependent Anaerobic Ammonium Oxidation in Microbial Fuel Cell Integrated Hybrid Constructed Wetlands: A New Process. Sci. Total Environ. 2020, 698, 134248. [Google Scholar] [CrossRef]
- Yakar, A.; Türe, C.; Türker, O.C.; Vymazal, J.; Saz, Ç. Impacts of Various Filtration Media on Wastewater Treatment and Bioelectric Production in Up-Flow Constructed Wetland Combined with Microbial Fuel Cell (UCW-MFC). Ecol. Eng. 2018, 117, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Hao, L.; Cao, J.; Zhou, K.; Fang, F.; Feng, Q.; Luo, J. Mechanism of Fe–C Micro-Electrolysis Substrate to Improve the Performance of CW-MFC with Different Factors: Insights of Microbes and Metabolic Function. Chemosphere 2022, 304, 135410. [Google Scholar] [CrossRef]
- Tamta, P.; Rani, N.; Mittal, Y.; Yadav, A.K. Evaluating the Potential of Multi-Anodes in Constructed Wetlands Coupled with Microbial Fuel Cells for Treating Wastewater and Bioelectricity Generation under High Organic Loads. Energies 2023, 16, 784. [Google Scholar] [CrossRef]
- Kesarwani, S.; Panwar, D.; Mal, J.; Pradhan, N.; Rani, R. Constructed Wetland Coupled Microbial Fuel Cell: A Clean Technology for Sustainable Treatment of Wastewater and Bioelectricity Generation. Fermentation 2022, 9, 6. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Terenzi, V. Rhizosphere Microbial Communities and Heavy Metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Habitat | References |
|---|---|---|
| Acidiphilium cryptum | Coal/copper strip mine soil/sediment | [38,39] |
| Acidithiobacillus ferrooxidans | Acid mine drainage water | [40] |
| Acinetobacter johsonii | Marine water | [41] |
| Alcaligenes faecalis | Ubiquitous/wastewater | [42] |
| Arcobacter butzleri | Freshwater/seawater | [33] |
| Brevibacillus agri | Paper wastewater treatment sludge | [43] |
| Clostridium ljungdahlii | Anaerobic digesters | [44] |
| Clostridium spp. | Ubiquitous in environment | [44,45] |
| Desulfuromonas sp. | Salt marsh sediment | [30] |
| Dietzia sp. | River estuary sediment | [46] |
| Enterococcus faecalis | Human gut | [37] |
| Escherichia coli | Ubiquitous/wastewater | [47] |
| Geobacter metallireducens | Soil/sediment | [47] |
| Geobacter sulfurreducens | Soil/sediment | [8] |
| Leptothrix sp. | Aquatic environments/wastewater | [48] |
| Methanobacterium palustre | Hot springs/anaerobic digesters | [49] |
| Methanococcus maripaludis | Salt marsh sediment | [44] |
| Nocardiopsis sp. | Saline and alkaline soil/marine ecosystem | [50] |
| Ochrobactrum anthropi YZ-1 | Wastewater | [51] |
| Pyrococcus furiosus | Salt marsh sediment | [52] |
| Shewanella oneidensis | Deep sea anaerobic habitats/soil | [53] |
| Sporomusa ovata | Sugar beet leaf (endophyte) | [53] |
| Thioalobacter | Salt marsh sediment | [30] |
| Thiomicrorhabdus spp. | Deep-sea hydrothermal vents | [25] |
| Pleomorphomonas sp. | Plant roots (endophyte) | [36,54] |
| Rahnella sp. | Plant roots (endophyte) | [36,55] |
| Shinella sp. | Sugar cane steam (endophyte) | [36,56] |
| Staphylococcus aureus | Human gut | [37] |
| Streptococcus agalactiae | Human gut | [37] |
| Winogradskyella poriferorum | Marine water | [41] |
| Plant Species | Habitat, Experimental Time | Max. Voltage (mV) | Contaminant Removal (%) | Reference |
|---|---|---|---|---|
Typha orientalis | Soil, 150 days | 137.12 ± 13.08 | Spiked Cd(II), 20 mg/kg: 30.2%, mainly bioaccumulation in roots | [124] |
Oryza rufipogon | Soil, 150 days | 350.50 ± 74.89 | Spiked Cd(II), 20 mg/kg: 22.8%, mainly bioaccumulation in roots | [124] |
| Oryza rufipogon + Chestnut biochar (5%) | 350.50 ± 74.89 | Cd(II), 20 mg/kg: 31.7% | ||
Pennisetum alopecuroides | Soil, 96 days | 220 at day 0 | Spiked Cr(VI), 50–500 mg/Kg: 75% | [126] |
| Soil, 10 months | Spiked Cr(VI), 50 mg/kg: 65% | [127] | ||
Phragmites communis | Soil, 96 days | 220 at day 0 | Spiked Cr(VI), 50–500 mg/Kg: 75% | [126] |
| Soil, 10 months | Spiked Cr(VI), 50 mg/kg: 78% | [127] | ||
Acorus Calamus | Sediment, 367 days | 61.4 mV (medium) | Pyrene, 3.2 mg/kg (87%) and benzo[a]pyrene, 1.7 mg/kg: 75% | [128] |
| Sediment (Constructed wetlands) | 36.43 mW/m2 | Cr(VI), 12.07 mg/L in wastewater: 99% | [129] | |
Vallisneria spiralis | Sediment | 121.7 mW | Spiked Pyrene and phenantrene, 10 mg/kg: 88.2% | [125] |
Ipomoea aquatica | Sediment | 114 ± 5.89 mV | Spiked Cu, 170 mg/kg: 60% | [130] |
Lolium perenne | Plant floated in water; 120–200 h | 55 mA/m2 | Cr(VI), 10–20 mg/L: 90% | [131] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garbini, G.L.; Barra Caracciolo, A.; Grenni, P. Electroactive Bacteria in Natural Ecosystems and Their Applications in Microbial Fuel Cells for Bioremediation: A Review. Microorganisms 2023, 11, 1255. https://doi.org/10.3390/microorganisms11051255
Garbini GL, Barra Caracciolo A, Grenni P. Electroactive Bacteria in Natural Ecosystems and Their Applications in Microbial Fuel Cells for Bioremediation: A Review. Microorganisms. 2023; 11(5):1255. https://doi.org/10.3390/microorganisms11051255
Chicago/Turabian StyleGarbini, Gian Luigi, Anna Barra Caracciolo, and Paola Grenni. 2023. "Electroactive Bacteria in Natural Ecosystems and Their Applications in Microbial Fuel Cells for Bioremediation: A Review" Microorganisms 11, no. 5: 1255. https://doi.org/10.3390/microorganisms11051255