
Bacillus amyloliquefaciens: Harnessing Its Potential for Industrial, Medical, and Agricultural Applications—A Comprehensive Review


Abstract
:1. Introduction
2. Genetic Manipulation of Bacillus amyloliquefaciens
2.1. Development and Application of CRISPR-Based Genetic Toolboxes in Bacillus amyloliquefaciens Strains
2.2. Enhancing Alkaline Protease Production and Antifungal Properties of Bacillus amyloliquefaciens through Genetic Engineering
3. Gene Expression Using Bacillus amyloliquefaciens
3.1. Investigating Plant-Bacteria Interaction: Bacillus amyloliquefaciens and Sclerotinia sclerotiorum in Soybean Plants
3.2. Cloning and Expression of Bacillus amyloliquefaciens Transglutaminase Gene in E. coli for Food Industry Applications
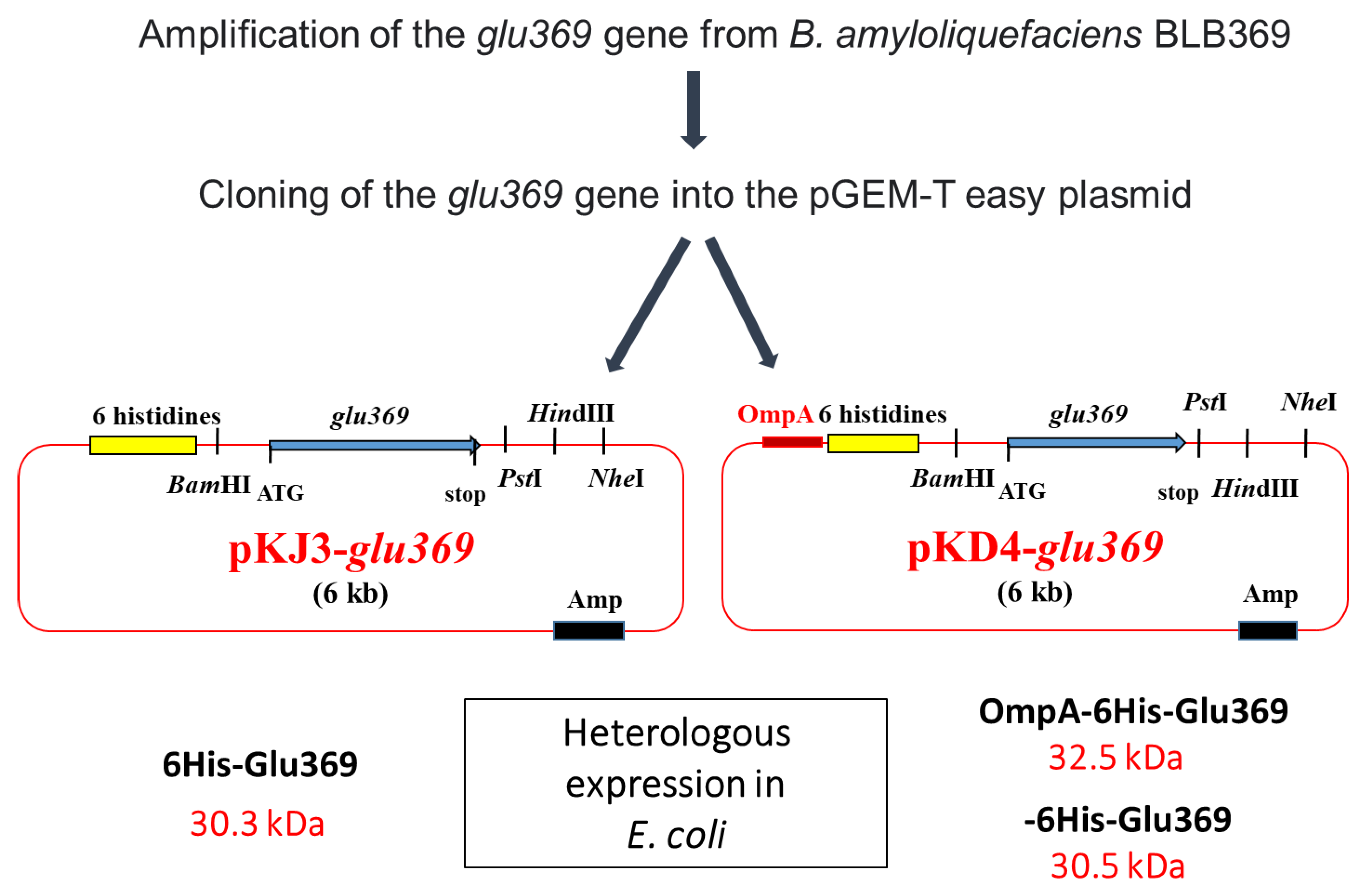
3.3. Heterologous Expression and Periplasmic Secretion of an Antifungal Bacillus amyloliquefaciens BLB 369 Endo-β-1,3-1,4-Glucanase in Escherichia coli
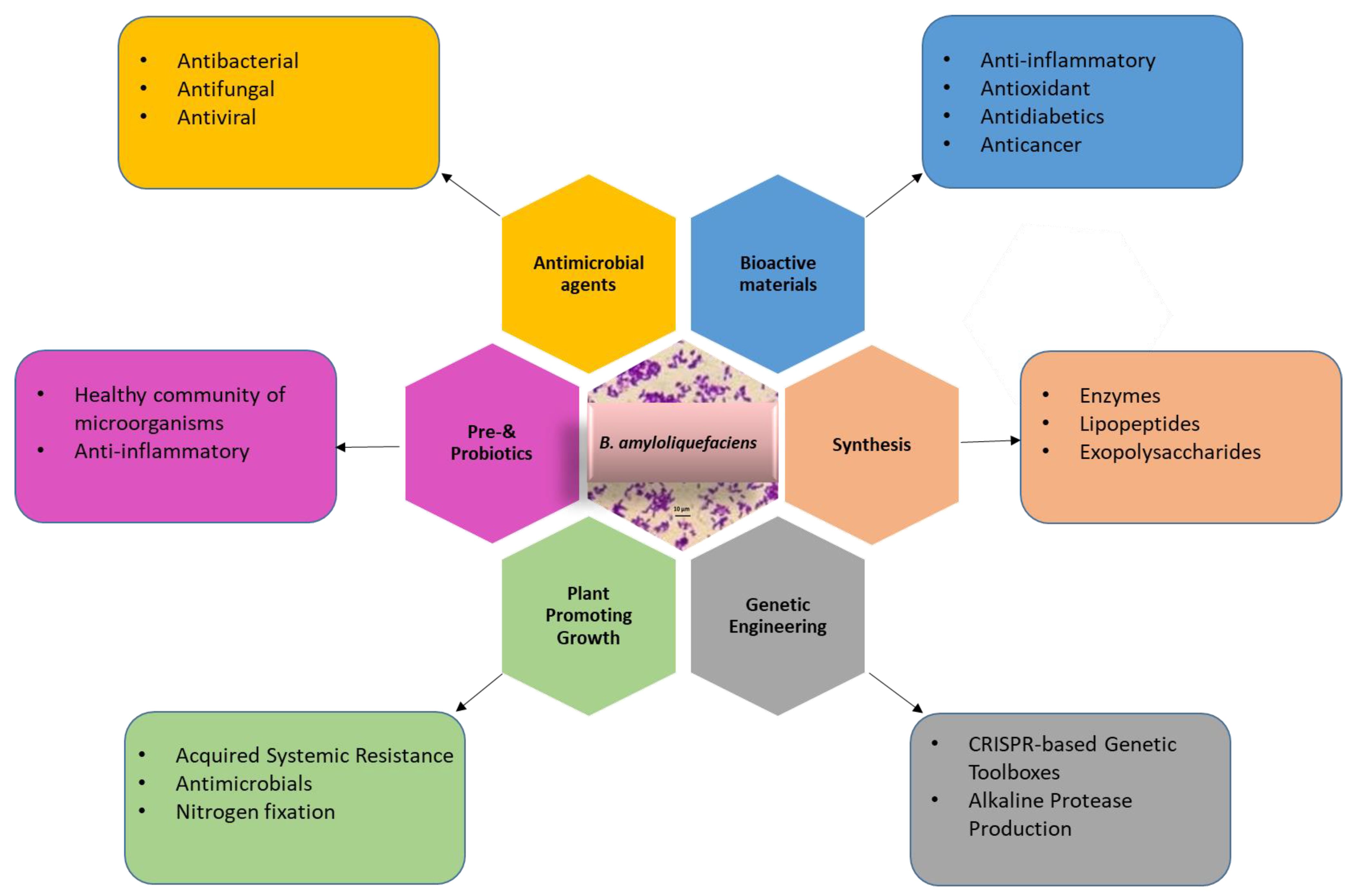
4. The Importance and Applications of the Bacillus amyloliquefaciens
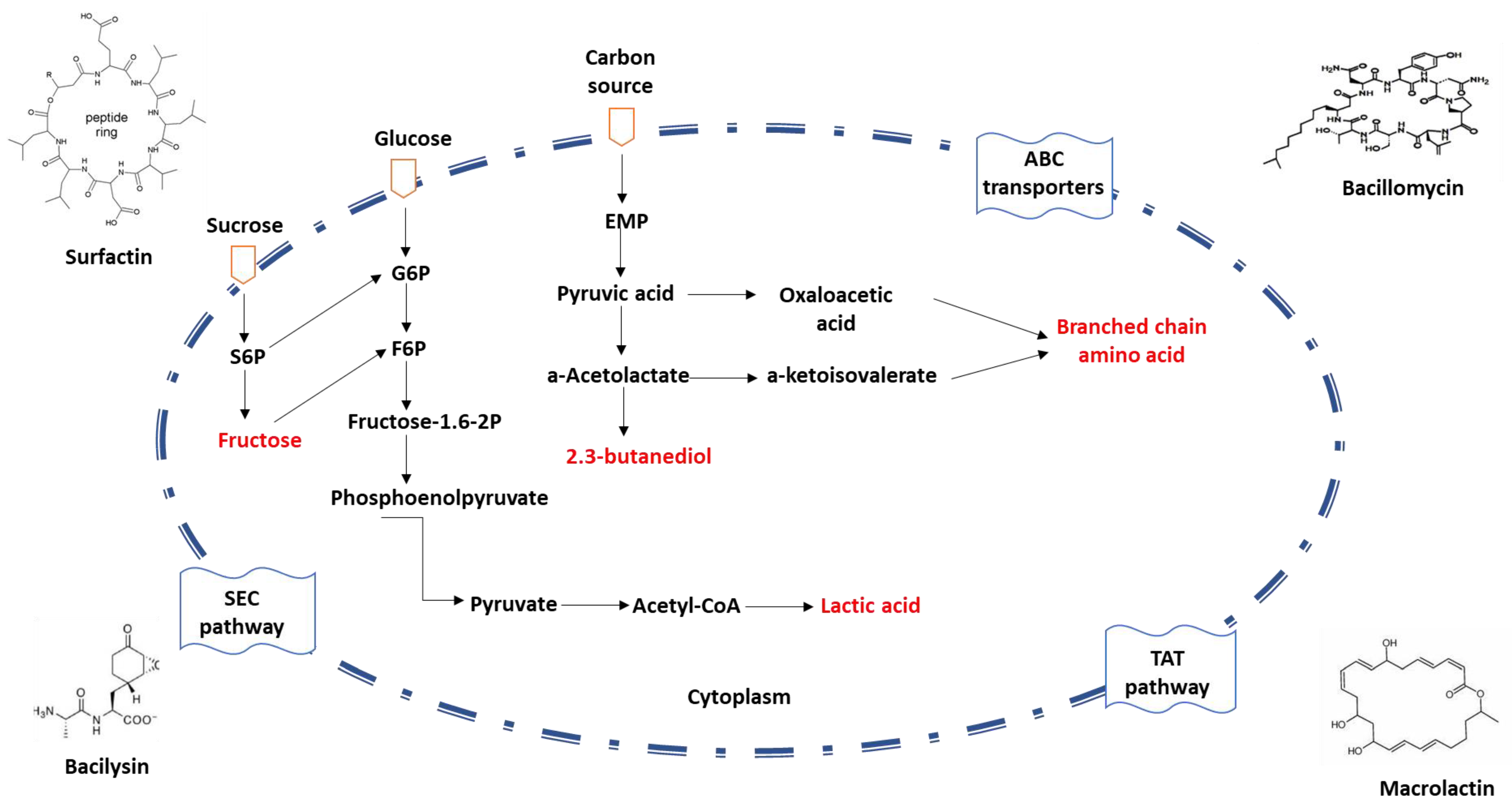
4.1. Industrial Application of Bacillus amyloliquefaciens
4.2. Medical Application of Bacillus amyloliquefaciens
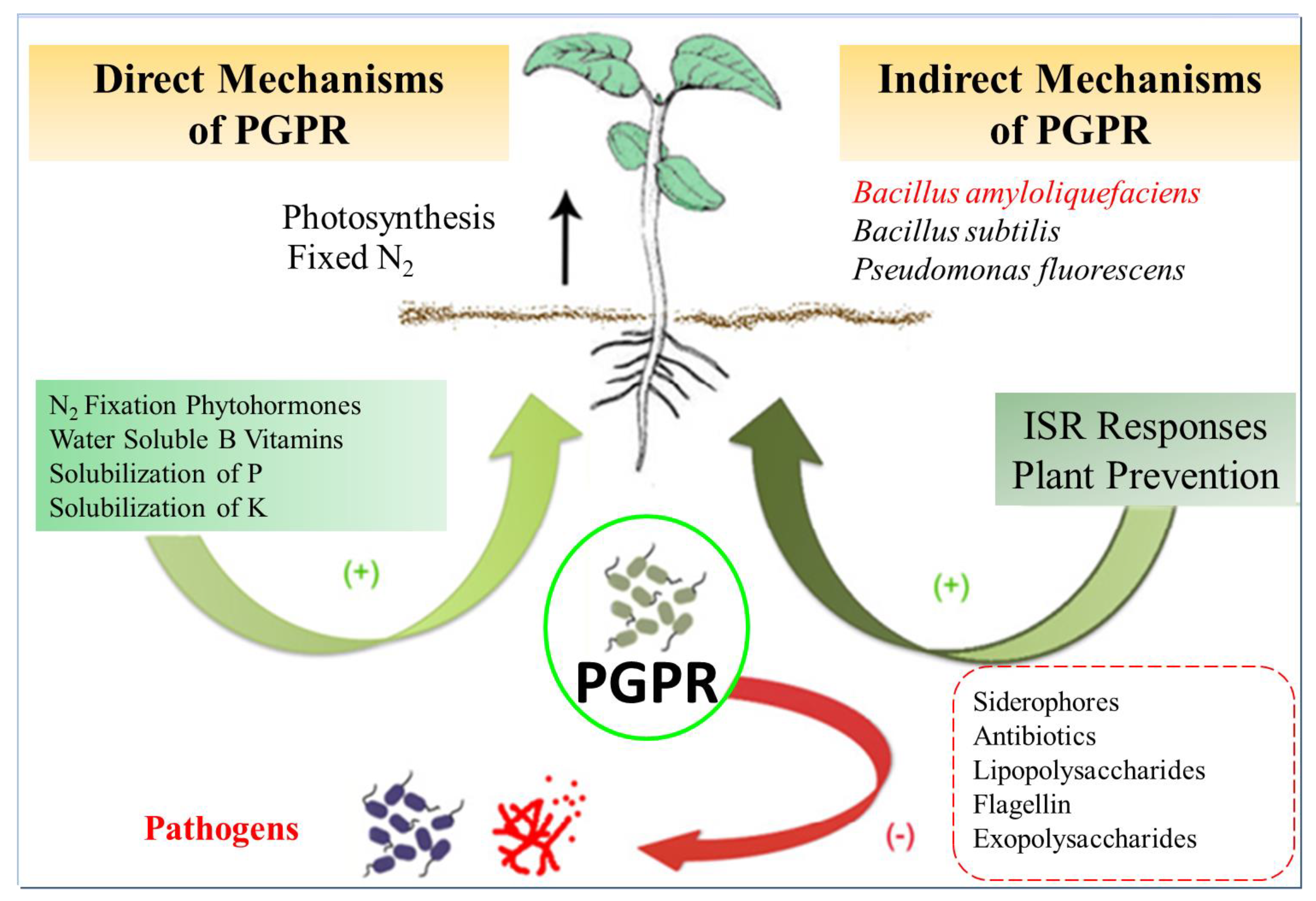
4.3. Application of Bacillus amyloliquefaciens in Agriculture
5. Adaptation of Bacillus amyloliquefaciens to Acidic Environments: Understanding Acid Tolerance Mechanisms
6. Conclusions and Perspectives
Funding
Data Availability Statement
Conflicts of Interest
References
- Zakataeva, N.P.; Nikitina, O.V.; Gronskiy, S.V.; Romanenkov, D.V.; Livshits, V.A. A simple method to introduce marker-free genetic modifications into the chromosome of naturally nontransformable Bacillus amyloliquefaciens strains. Appl. Microbiol. Biotechnol. 2010, 85, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, H.; Lv, Y.; Bai, Y.; Luo, H.; Shi, P.; Huang, H.; Yao, B. Construction of a Rapid Feather-Degrading Bacterium by Overexpression of a Highly Efficient Alkaline Keratinase in Its Parent Strain Bacillus amyloliquefaciens K11. J. Agric. Food Chem. 2016, 64, 78–84. [Google Scholar] [CrossRef]
- Feng, J.; Gu, Y.; Quan, Y.; Cao, M.; Gao, W.; Zhang, W.; Wang, S.; Yang, C.; Song, C. Improved poly-gamma-glutamic acid production in Bacillus amyloliquefaciens by modular pathway engineering. Metab. Eng. 2015, 32, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Shim, H.; Shin, J.H.; Kim, K.S. Antagonistic Activities of Bacillus spp. Strains Isolated from Tidal Flat Sediment towards Anthracnose Pathogens Colletotrichum acutatum and C. gloeosporioides in South Korea. Plant Pathol. J. 2015, 31, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Zalila-Kolsi, I.; Kessentini, S.; Tounsi, S.; Jamoussi, K. Optimization of Bacillus amyloliquefaciens BLB369 Culture Medium by Response Surface Methodology for Low Cost Production of Antifungal Activity. Microorganisms 2022, 10, 830. [Google Scholar] [CrossRef]
- Ye, M.; Sun, L.; Yang, R.; Wang, Z.; Qi, K. The optimization of fermentation conditions for producing cellulase of Bacillus amyloliquefaciens and its application to goose feed. R. Soc. Open Sci. 2017, 4, 171012. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, B.; Li, X.; Xu, X.; Li, D.; Zeng, F.; Zhou, C.; Liu, Y.; Li, Y.; Lu, F. Multiple Modular Engineering of Bacillus amyloliquefaciens Cell Factories for Enhanced Production of Alkaline Proteases From B. Clausii. Front. Bioeng. Biotechnol. 2022, 10, 866066. [Google Scholar] [CrossRef]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A Review on the Biotechnological Applications of the Operational Group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef]
- Xin, Q.; Chen, Y.; Chen, Q.; Wang, B.; Pan, L. Development and application of a fast and efficient CRISPR-based genetic toolkit in Bacillus amyloliquefaciens LB1ba02. Microb. Cell Fact. 2022, 21, 99. [Google Scholar] [CrossRef]
- Jiang, C.; Ye, C.; Liu, Y.; Huang, K.; Jiang, X.; Zou, D.; Li, L.; Han, W.; Wei, X. Genetic engineering for enhanced production of a novel alkaline protease BSP-1 in Bacillus amyloliquefaciens. Front. Bioeng. Biotechnol. 2022, 10, 977215. [Google Scholar] [CrossRef]
- Wang, S.; Wang, R.; Zhao, X.; Ma, G.; Liu, N.; Zheng, Y.; Tan, J.; Qi, G. Systemically engineering Bacillus amyloliquefaciens for increasing its antifungal activity and green antifungal lipopeptides production. Front. Bioeng. Biotechnol. 2022, 10, 961535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Huo, K.; Song, X.; Quan, Y.; Wang, S.; Zhang, Z.; Gao, W.; Yang, C. Engineering of a genome-reduced strain Bacillus amyloliquefaciens for enhancing surfactin production. Microb. Cell Fact. 2020, 19, 223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, H.; Zhen, J.; Shu, W.; Yang, S.; Xu, J.; Song, H.; Ma, Y. Multiplex genetic engineering improves endogenous expression of mesophilic alpha-amylase gene in a wild strain Bacillus amyloliquefaciens 205. Int. J. Biol. Macromol. 2020, 165, 609–618. [Google Scholar] [CrossRef]
- Su, Y.T.; Liu, C.; Long, Z.; Ren, H.; Guo, X.H. Improved Production of Spores and Bioactive Metabolites from Bacillus amyloliquefaciens in Solid-State Fermentation by a Rapid Optimization Process. Probiotics Antimicrob. Proteins 2019, 11, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Wilson, G.A.; Young, F.E. Recognition sequence of specific endonuclease Bam HI from Bacillus amyloliquefaciens H. Nature 1977, 265, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Long, Z.; Liang, X.; Guo, S.; Ning, N.; Yang, L.; Wang, X.; Lu, B.; Gao, J. The role of a β-1, 3-1, 4-glucanase derived from Bacillus amyloliquefaciens FS6 in the protection of ginseng against Botrytis cinerea and Alternaria panax. Biol. Control 2021, 164, 104765. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Brandán, C.P. Bacillus amyloliquefaciens strain enhances rhizospheric microbial growth and reduces root and stem rot in a degraded agricultural system. Rhizosphere 2022, 22, 100544. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants—With special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef]
- Kasim, W.A.; Gaafar, R.M.; Abou-Ali, R.M.; Omar, M.N.; Hewait, H.M. Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann. Agric. Sci. 2016, 61, 217–227. [Google Scholar] [CrossRef]
- Cai, D.; Rao, Y.; Zhan, Y.; Wang, Q.; Chen, S. Engineering Bacillus for efficient production of heterologous protein: Current progress, challenge and prospect. J. Appl. Microbiol. 2019, 126, 1632–1642. [Google Scholar] [CrossRef]
- Luo, L.; Zhao, C.; Wang, E.; Raza, A.; Yin, C. Bacillus amyloliquefaciens as an excellent agent for biofertilizer and biocontrol in agriculture: An overview for its mechanisms. Microbiol. Res. 2022, 259, 127016. [Google Scholar] [CrossRef]
- Vehmaanpera, J.; Steinborn, G.; Hofemeister, J. Genetic manipulation of Bacillus amyloliquefaciens. J. Biotechnol. 1991, 19, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.T.; Ji, J.B.; Liu, Y.C.; Ye, B.; Zhou, C.Y.; Yan, X. Artificial induction of genetic competence in Bacillus amyloliquefaciens isolates. Biotechnol. Lett. 2016, 38, 2109–2117. [Google Scholar] [CrossRef]
- Devaraj, K.; Aathika, S.; Periyasamy, K.; Manickam Periyaraman, P.; Palaniyandi, S.; Subramanian, S. Production of thermostable multiple enzymes from Bacillus amyloliquefaciens KUB29. Nat. Prod. Res. 2019, 33, 1674–1677. [Google Scholar] [CrossRef] [PubMed]
- Duarte, L.S.; Barse, L.Q.; Dalberto, P.F.; da Silva, W.T.S.; Rodrigues, R.C.; Machado, P.; Basso, L.A.; Bizarro, C.V.; Ayub, M.A.Z. Cloning and expression of the Bacillus amyloliquefaciens transglutaminase gene in E. coli using a bicistronic vector construction. Enzym. Microb. Technol. 2020, 134, 109468. [Google Scholar] [CrossRef]
- Gao, W.; He, Y.; Zhang, F.; Zhao, F.; Huang, C.; Zhang, Y.; Zhao, Q.; Wang, S.; Yang, C. Metabolic engineering of Bacillus amyloliquefaciens LL3 for enhanced poly-gamma-glutamic acid synthesis. Microb. Biotechnol. 2019, 12, 932–945. [Google Scholar] [CrossRef]
- Gould, A.R.; May, B.K.; Elliott, W.H. Release of extracellular enzymes from Bacillus amyloliquefaciens. J. Bacteriol. 1975, 122, 34–40. [Google Scholar] [CrossRef]
- Xue, M.; Wu, Y.; Hong, Y.; Meng, Y.; Xu, C.; Jiang, N.; Li, Y.; Liu, W.; Fan, Y.; Zhou, Y. Effects of dietary Bacillus amyloliquefaciens on the growth, immune responses, intestinal microbiota composition and disease resistance of yellow catfish, Pelteobagrus fulvidraco. Front. Cell Infect. Microbiol. 2022, 12, 1047351. [Google Scholar] [CrossRef] [PubMed]
- Yang, W. Components of rhizospheric bacterial communities of barley and their potential for plant growth promotion and biocontrol of Fusarium wilt of watermelon. Braz. J. Microbiol. 2019, 50, 749–757. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, J.; Tan, M.; Yu, Z.; Yang, S.; Zheng, H.; Song, H. Construction of a plasmid interspecific transfer system in Bacillus species with the counter-selectable marker mazF. J. Ind. Microbiol. Biotechnol. 2018, 45, 417–428. [Google Scholar] [CrossRef]
- Tjalsma, H.; Antelmann, H.; Jongbloed, J.D.; Braun, P.G.; Darmon, E.; Dorenbos, R.; Dubois, J.Y.; Westers, H.; Zanen, G.; Quax, W.J.; et al. Proteomics of protein secretion by Bacillus subtilis: Separating the “secrets” of the secretome. Microbiol. Mol. Biol. Rev. 2004, 68, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Jongbloed, J.D.; Antelmann, H.; Hecker, M.; Nijland, R.; Bron, S.; Airaksinen, U.; Pries, F.; Quax, W.J.; van Dijl, J.M.; Braun, P.G. Selective contribution of the twin-arginine translocation pathway to protein secretion in Bacillus subtilis. J. Biol. Chem. 2002, 277, 44068–44078. [Google Scholar] [CrossRef] [PubMed]
- Freudl, R. Signal peptides for recombinant protein secretion in bacterial expression systems. Microb. Cell Fact. 2018, 17, 52. [Google Scholar] [CrossRef]
- Liu, J.; Hu, X.; He, H.; Zhang, X.; Guo, J.; Bai, J.; Cheng, Y. Digital gene expression profiling of the transcriptional response to Sclerotinia sclerotiorum and its antagonistic bacterium Bacillus amyloliquefaciens in soybean. Front. Microbiol. 2022, 13, 1025771. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Sellami, S.; Tounsi, S.; Jamoussi, K. Heterologous expression and periplasmic secretion of an antifungal Bacillus amyloliquefaciens BLB 369 endo-β-1, 3-1, 4-glucanase in Escherichia coli. J. Phytopathol. 2018, 166, 28–33. [Google Scholar] [CrossRef]
- Contesini, F.J.; Melo, R.R.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; He, P.; Munir, S.; He, P.; Li, X.; Li, Y.; Wu, J.; Wu, Y.; Yang, L.; He, P. Efficacy of plant growth promoting bacteria Bacillus amyloliquefaciens B9601-Y2 for biocontrol of southern corn leaf blight. Biol. Control 2019, 139, 104080. [Google Scholar] [CrossRef]
- do Nascimento Silva, J.; Mendes, L.W.; Antunes, J.E.L.; Melo, V.M.M.; da Silva Oliveira, F.A.; de Almeida Lopes, A.C.; da Silva, V.B.; de Araujo Pereira, A.P.; Valente, S.E.S.; Araujo, A.S.F. Diversity, structure, and composition of plant growth-promoting bacteria in soil from Brazilian Cerrado. Rhizosphere 2021, 20, 100435. [Google Scholar] [CrossRef]
- Luo, Z.; Yan, Y.; Du, S.; Zhu, Y.; Pan, F.; Wang, R.; Xu, Z.; Xu, X.; Li, S.; Xu, H. Recent advances and prospects of Bacillus amyloliquefaciens as microbial cell factories: From rational design to industrial applications. Crit. Rev. Biotechnol. 2022, 1–19. [Google Scholar] [CrossRef]
- Ren, L.; Yuan, Z.; Xie, T.; Wu, D.; Kang, Q.; Li, J.; Li, J. Extraction and characterization of cyclic lipopeptides with antifungal and antioxidant activities from Bacillus amyloliquefaciens. J. Appl. Microbiol. 2022, 133, 3573–3584. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. Microbiologyopen 2019, 8, e00813. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Sun, B.; Yang, Y.; Jin, B.; Zhuang, G.; Bai, Z.; Zhuang, X. Efficiency and mechanism of reducing ammonia volatilization in alkaline farmland soil using Bacillus amyloliquefaciens biofertilizer. Environ. Res. 2021, 202, 111672. [Google Scholar] [CrossRef]
- Yamamoto, S.; Shiraishi, S.; Kawagoe, Y.; Mochizuki, M.; Suzuki, S. Impact of Bacillus amyloliquefaciens S13-3 on control of bacterial wilt and powdery mildew in tomato. Pest. Manag. Sci. 2015, 71, 722–727. [Google Scholar] [CrossRef]
- Zayabaatar, E.; Huang, C.M.; Pham, M.T.; Ganzorig, B.; Wang, S.M.; Chen, C.C. Bacillus amyloliquefaciens Increases the GABA in Rice Seed for Upregulation of Type I Collagen in the Skin. Curr. Microbiol. 2023, 80, 128. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-J.; Zayabaatar, E.; Wang, S.-M.; Keshari, S.; Peng, W.-H.; Kung, H.-N.; Lee, Y.-H. Bacillus amyloliquefaciens-Inoculated GABA-Rich Rice Upregulate Neuropeptide Y to Relieve Psychological Stress through Mediations of GABAB Receptor and Vagus Nerves. Biol. Bull. 2023, 50, 186–193. [Google Scholar] [CrossRef]
- Zhang, Z.; He, P.; Cai, D.; Chen, S. Genetic and metabolic engineering for poly-gamma-glutamic acid production: Current progress, challenges, and prospects. World J. Microbiol. Biotechnol. 2022, 38, 208. [Google Scholar] [CrossRef]
- Fang, J.; Liu, Y.; Huan, C.; Xu, L.; Ji, G.; Yan, Z. Comparison of poly-γ-glutamic acid production between sterilized and non-sterilized solid-state fermentation using agricultural waste as substrates. J. Clean. Prod. 2020, 255, 120248. [Google Scholar] [CrossRef]
- Doan, C.T.; Chen, C.L.; Nguyen, V.B.; Tran, T.N.; Nguyen, A.D.; Wang, S.L. Conversion of Pectin-Containing By-Products to Pectinases by Bacillus amyloliquefaciens and Its Applications on Hydrolyzing Banana Peels for Prebiotics Production. Polymers 2021, 13, 1483. [Google Scholar] [CrossRef]
- Du, L.; Wang, J.; Chen, W.; Chen, J.; Zheng, Q.; Fang, X.; Liao, Z. Isolation and Purification of Bacillus amyloliquefaciens D1 Protease and Its Application in the Fermentation of Soybean Milk to Produce Large Amounts of Free Amino Acids. Appl. Biochem. Biotechnol. 2023, 195, 451–466. [Google Scholar] [CrossRef]
- Lin, L.Z.; Zheng, Q.W.; Wei, T.; Zhang, Z.Q.; Zhao, C.F.; Zhong, H.; Xu, Q.Y.; Lin, J.F.; Guo, L.Q. Isolation and Characterization of Fengycins Produced by Bacillus amyloliquefaciens JFL21 and Its Broad-Spectrum Antimicrobial Potential against Multidrug-Resistant Foodborne Pathogens. Front. Microbiol. 2020, 11, 579621. [Google Scholar] [CrossRef]
- Soussi, S.; Essid, R.; Hardouin, J.; Gharbi, D.; Elkahoui, S.; Tabbene, O.; Cosette, P.; Jouenne, T.; Limam, F. Utilization of Grape Seed Flour for Antimicrobial Lipopeptide Production by Bacillus amyloliquefaciens C5 Strain. Appl. Biochem. Biotechnol. 2019, 187, 1460–1474. [Google Scholar] [CrossRef]
- Gao, L.; She, M.; Shi, J.; Cai, D.; Wang, D.; Xiong, M.; Shen, G.; Gao, J.; Zhang, M.; Yang, Z.; et al. Enhanced production of iturin A by strengthening fatty acid synthesis modules in Bacillus amyloliquefaciens. Front. Bioeng. Biotechnol. 2022, 10, 974460. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Qian, S.; Muhammad, U.; Jiang, X.; Han, J.; Lu, Z. Effect of fructose on promoting fengycin biosynthesis in Bacillus amyloliquefaciens fmb-60. J. Appl. Microbiol. 2016, 121, 1653–1664. [Google Scholar] [CrossRef]
- Lu, J.Y.; Zhou, K.; Huang, W.T.; Zhou, P.; Yang, S.; Zhao, X.; Xie, J.; Xia, L.; Ding, X. A comprehensive genomic and growth proteomic analysis of antitumor lipopeptide bacillomycin Lb biosynthesis in Bacillus amyloliquefaciens X030. Appl. Microbiol. Biotechnol. 2019, 103, 7647–7662. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, B.; Xu, H.; Mei, X.; Xu, X.; Zhang, X.; Ni, J.; Li, W. Bacillus amyloliquefaciens SC06 Protects Mice against High-Fat Diet-Induced Obesity and Liver Injury via Regulating Host Metabolism and Gut Microbiota. Front. Microbiol. 2019, 10, 1161. [Google Scholar] [CrossRef]
- Jeong, D.Y.; Jeong, S.Y.; Zhang, T.; Wu, X.; Qiu, J.Y.; Park, S. Chungkookjang, a soy food, fermented with Bacillus amyloliquefaciens protects gerbils against ishcmeic stroke injury, and post-stroke hyperglycemia. Food Res. Int. 2020, 128, 108769. [Google Scholar] [CrossRef]
- WoldemariamYohannes, K.; Wan, Z.; Yu, Q.; Li, H.; Wei, X.; Liu, Y.; Wang, J.; Sun, B. Prebiotic, Probiotic, Antimicrobial, and Functional Food Applications of Bacillus amyloliquefaciens. J. Agric. Food Chem. 2020, 68, 14709–14727. [Google Scholar] [CrossRef]
- Zhou, P.; Chen, W.; Zhu, Z.; Zhou, K.; Luo, S.; Hu, S.; Xia, L.; Ding, X. Comparative Study of Bacillus amyloliquefaciens X030 on the Intestinal Flora and Antibacterial Activity against Aeromonas of Grass Carp. Front. Cell Infect. Microbiol. 2022, 12, 815436. [Google Scholar] [CrossRef]
- Al-Dulaimi, M.; Algburi, A.; Abdelhameed, A.; Mazanko, M.S.; Rudoy, D.V.; Ermakov, A.M.; Chikindas, M.L. Antimicrobial and Anti-Biofilm Activity of Polymyxin E Alone and in Combination with Probiotic Strains of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against Clinical Isolates of Selected Acinetobacter spp.: A Preliminary Study. Pathogens 2021, 10, 1574. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.; Sheikh, A.; Ibrahim, H.I.M. Bacillus amyloliquefaciens Enriched Camel Milk Attenuated Colitis Symptoms in Mice Model. Nutrients 2022, 14, 1967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, Y.; Xu, H.; Liang, C.; Zhai, Z. Bacillus amyloliquefaciens BLCC1-0238 Alone or in Combination with Mannan-Oligosaccharides Alleviates Subclinical Necrotic Enteritis in Broilers. Probiotics Antimicrob. Proteins 2022, 14, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Morozova, M.V.; Kalmykova, G.V.; Akulova, N.I.; Ites, Y.V.; Korkina, V.I.; Litvinova, E.A. Autoclaved Diet with Inactivated Spores of Bacillus spp. Decreased Reproductive Performance of Muc2(−/−) and Muc2(+/−) Mice. Animals 2022, 12, 2399. [Google Scholar] [CrossRef]
- Al-Mutar, D.M.K.; Alzawar, N.S.A.; Noman, M.; Azizullah; Li, D.; Song, F. Suppression of Fusarium Wilt in Watermelon by Bacillus amyloliquefaciens DHA55 through Extracellular Production of Antifungal Lipopeptides. J. Fungi 2023, 9, 336. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Z.; Liu, Z.; Wang, K.; Xu, W. Membrane disruption of Fusarium oxysporum f. sp. niveum induced by myriocin from Bacillus amyloliquefaciens LZN01. Microb. Biotechnol. 2021, 14, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, Z.; Li, N.; Wang, Y.; Feng, J.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil. Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- Gautam, S.; Chauhan, A.; Sharma, R.; Sehgal, R.; Shirkot, C.K. Potential of Bacillus amyloliquefaciens for biocontrol of bacterial canker of tomato incited by Clavibacter michiganensis ssp. michiganensis. Microb. Pathog. 2019, 130, 196–203. [Google Scholar] [CrossRef]
- European Food Safety, A.; Anastassiadou, M.; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Crivellente, F.; et al. Peer review of the pesticide risk assessment of the active substance Bacillus amyloliquefaciens strain AH2. EFSA J. 2020, 18, e06156. [Google Scholar] [CrossRef] [PubMed]
- Sunita, K.; Mishra, I.; Mishra, J.; Prakash, J.; Arora, N.K. Secondary Metabolites from Halotolerant Plant Growth Promoting Rhizobacteria for Ameliorating Salinity Stress in Plants. Front. Microbiol. 2020, 11, 567768. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Wani, S.H.; Pande, A.; Imran, M.; Kang, S.M.; Rahim, W.; Khan, S.A.; Bhatta, D.; Kwon, E.H.; et al. A Review on the Role of Endophytes and Plant Growth Promoting Rhizobacteria in Mitigating Heat Stress in Plants. Microorganisms 2022, 10, 1286. [Google Scholar] [CrossRef]
- Tiwari, S.; Prasad, V.; Chauhan, P.S.; Lata, C. Bacillus amyloliquefaciens Confers Tolerance to Various Abiotic Stresses and Modulates Plant Response to Phytohormones through Osmoprotection and Gene Expression Regulation in Rice. Front. Plant Sci. 2017, 8, 1510. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Choudhary, M.; Chandra, P.; Dixit, B.; Nehra, V.; Choudhary, U.; Choudhary, S. Plant growth-promoting microbes: Role and prospective in amelioration of salt stress. Commun. Soil Sci. Plant Anal. 2022, 53, 1692–1711. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Veronican Njeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Li, R.; Tao, J.; Huang, D.; Zhou, W.; Gao, L.; Wang, X.; Chen, H.; Huang, H. Investigating the effects of biodegradable microplastics and copper ions on probiotic (Bacillus amyloliquefaciens): Toxicity and application. J. Hazard. Mater. 2023, 443, 130081. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, N.; Goswami, G.; Boro, R.C.; Barooah, M. A pH-Dependent Gene Expression Enables Bacillus amyloliquefaciens MBNC to Adapt to Acid Stress. Curr. Microbiol. 2021, 78, 3104–3114. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Used Method | Product | References |
|---|---|---|---|
| B. amyloliquefaciens EH-9 | Fermentation of Germinated rice seed | GABA | [44] |
| B. amyloliquefaciens NK-A11 | Genetic modification of B. amyloliquefaciens LL3 a glutamic acid-independent γ-PGA-producing strain | γ-PGA | [26] |
| B. amyloliquefaciens JX-6 | Fermentation of corn stalk and soybean milk in sterilize and non-sterilized conditions | γ-PGA | [47] |
| B. amyloliquefaciens TKU050 | Bioprocessing of banana peel to reduce production cost | Pectinase/probiotic | [48] |
| B. amyloliquefaciens C5 | Growth and antimicrobial activity were optimized using grape seed flour substrate | Surfactin and bacillomycin D | [51] |
| B. amyloliquefaciens HZ-ADFTL2 | Genetic modification of B. amyloliquefaciens HZ-12 to enhance the yield | Iturin A | [52] |
| B. amyloliquefaciens fmb-60 | Addition of fructose to culture media to enhance the yield | Fengycin | [53] |
| B. amyloliquefaciens WH1 | Genetic modification was introduced to enhance the yield | Iturin and fengycin | [11] |
| B. amyloliquefaciens X030 | Added to various cancer cell lines to test anticancer activity | Bacillomycin Lb | [54] |
| B. amyloliquefaciens JFL21 | Tested against multidrug-resistant foodborne pathogens | Multiple lipopeptides | [50] |
| Strains | Test Subject | Condition | Bacterial Inoculating Method | Observed Effects | References |
|---|---|---|---|---|---|
| B. amyloliquefaciens SC06 | Mice | High-fat diet (HFD)-induced obesity | Bacteria combined with the HFD fed to the mice |
| [29] |
| B. amyloliquefaciens CKJ730 & CKJ731 | Mongolian gerbils | Ischemic stroke model | They were fed chungkookjang, which was fermented using the bacteria |
| [56] |
| B. amyloliquefaciens X030 | Grass carp | Infections | Fed to the grass carps |
| [58] |
| B. amyloliquefaciens B-1895 | Acinetobacter spp. Isolates | Acinetobacter growth and biofilm formation | Bacterial cell-free supernatant added to the Acinetobacter spp. isolates |
| [59] |
| B. amyloliquefaciens PBT-3 | Mice | 2,4,6-trinitro-benzenesulfonic acid (TNBS)-induced colitis model | Fed camel milk enriched with B. amyloliquefaciens PBT-3 (BEY) |
| [60] |
| B. amyloliquefaciens BLCC1-0238 | Broilers | Subclinical necrotic enteritis | Pretreatment with the BLCC1-0238 |
| [61] |
| Bacillus amyloliquefaciens B14 | Degraded soil | Degraded soil | Inoculation of the seeds |
| [17] |
| B. amyloliquefaciens L3 B. amyloliquefa-ciens DHA55 B. amyloliquefaciens LZN01 B. amyloliquefaciens subsp. plantarum FZB42 | Watermelon | Fusarium wilt caused by Fusarium oxysporum f.sp. niveum (Fon) | Agricultural use |
| [29,41,63,64] |
| Bacillus amyloliquefaciens B1408 | Cucumber | Fusarium wilt caused by Fusarium oxysporum f.sp. cucumerinum (Foc) | Agricultural use |
| [65] |
| B. amyloliquefaciens B9601-Y2 | Maize | southern corn leaf blight caused by Bipolaris maydis | Agricultural use |
| [37] |
| B. amyloliquefaciens S1 | Tomato | Bacterial canker caused by Clavibacter michiganensis | Agricultural use |
| [66] |
| B. amyloliquefaciens AH2 | Grapes | grey mold | Agricultural use |
| [67] |
| Bacillus amyloliquefa-ciens SQR9 | Saline soil | Saline soil | Agricultural use |
| [23,68,69] |
| Bacillus amyloliquefaciens NBRI-SN13 | rice cv. Saryu-52 | Seedlings subjected to various abiotic stresses (salt, drought, heat, cold, freezing, and desiccation) and phytohormone treatments | Agricultural use |
| [70] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalila-Kolsi, I.; Ben-Mahmoud, A.; Al-Barazie, R. Bacillus amyloliquefaciens: Harnessing Its Potential for Industrial, Medical, and Agricultural Applications—A Comprehensive Review. Microorganisms 2023, 11, 2215. https://doi.org/10.3390/microorganisms11092215
Zalila-Kolsi I, Ben-Mahmoud A, Al-Barazie R. Bacillus amyloliquefaciens: Harnessing Its Potential for Industrial, Medical, and Agricultural Applications—A Comprehensive Review. Microorganisms. 2023; 11(9):2215. https://doi.org/10.3390/microorganisms11092215
Chicago/Turabian StyleZalila-Kolsi, Imen, Afif Ben-Mahmoud, and Ray Al-Barazie. 2023. "Bacillus amyloliquefaciens: Harnessing Its Potential for Industrial, Medical, and Agricultural Applications—A Comprehensive Review" Microorganisms 11, no. 9: 2215. https://doi.org/10.3390/microorganisms11092215