
Intestinal Production of Alpha-Glucosidase Inhibitor by Bacillus coagulans Spores

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Measurement of α-Glucosidase Inhibition Activity
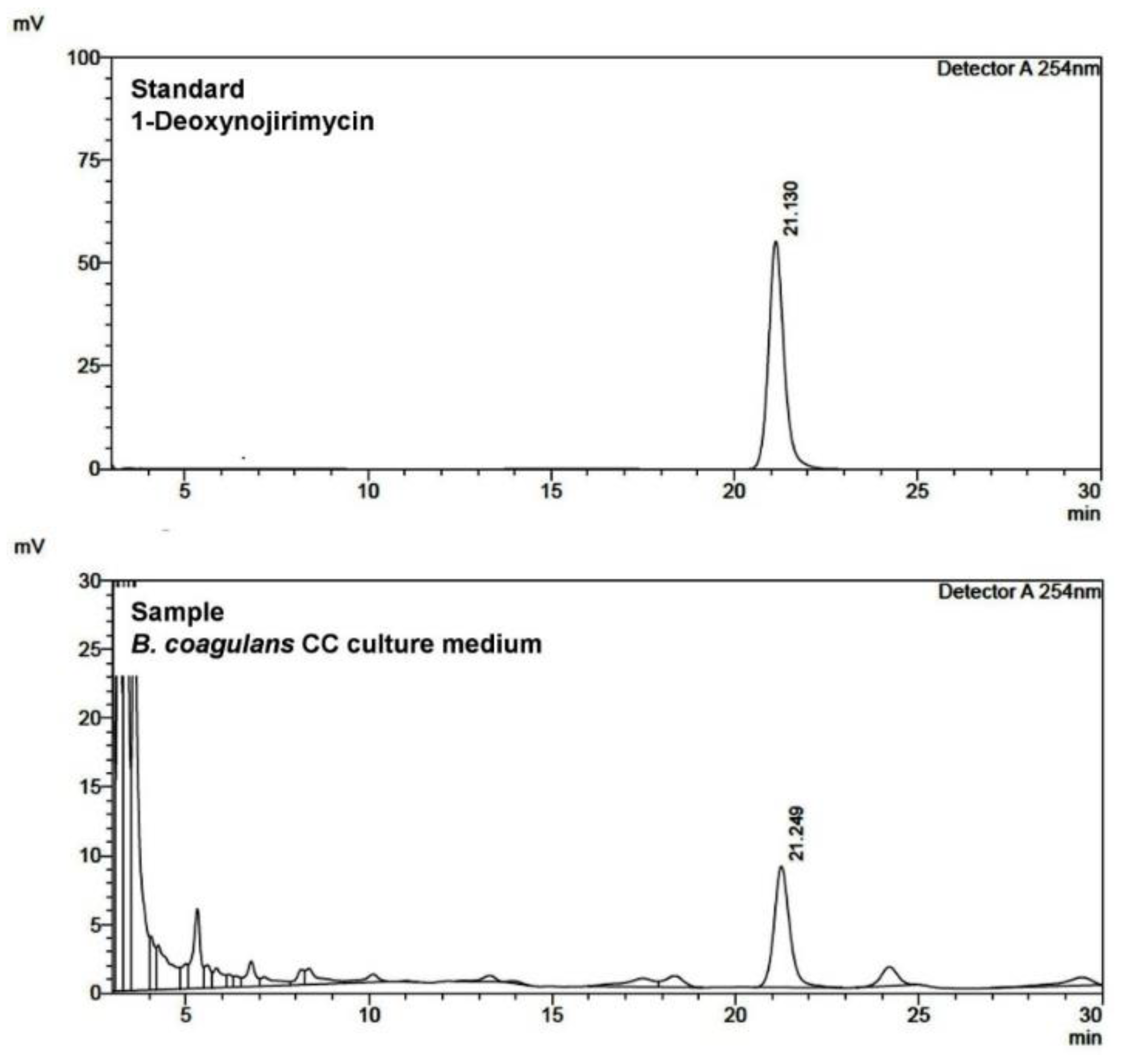
2.3. Identification of AGI
2.4. Sporulation
2.5. Animals
2.6. Statistical Analysis
3. Results
3.1. Strain
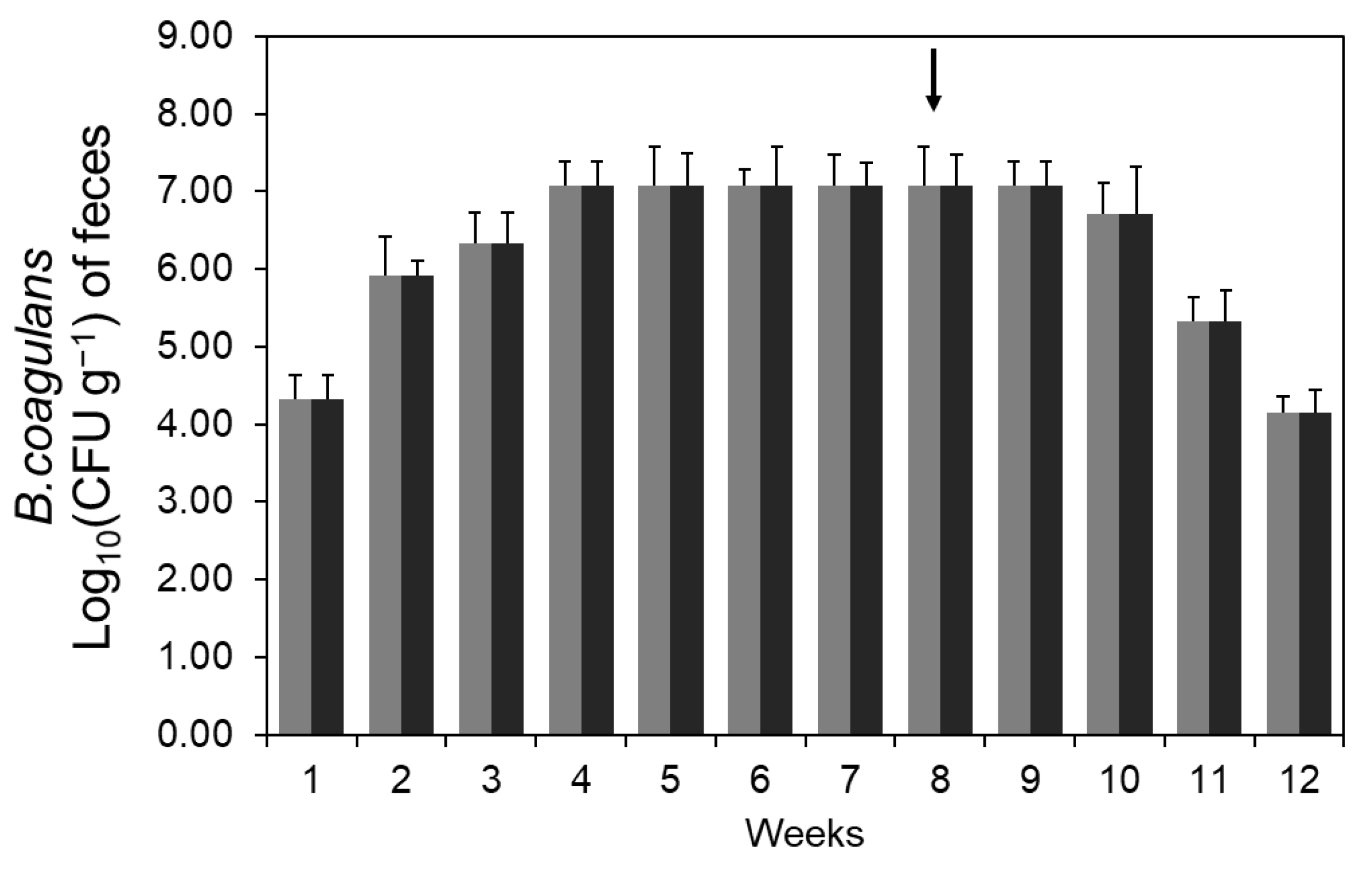
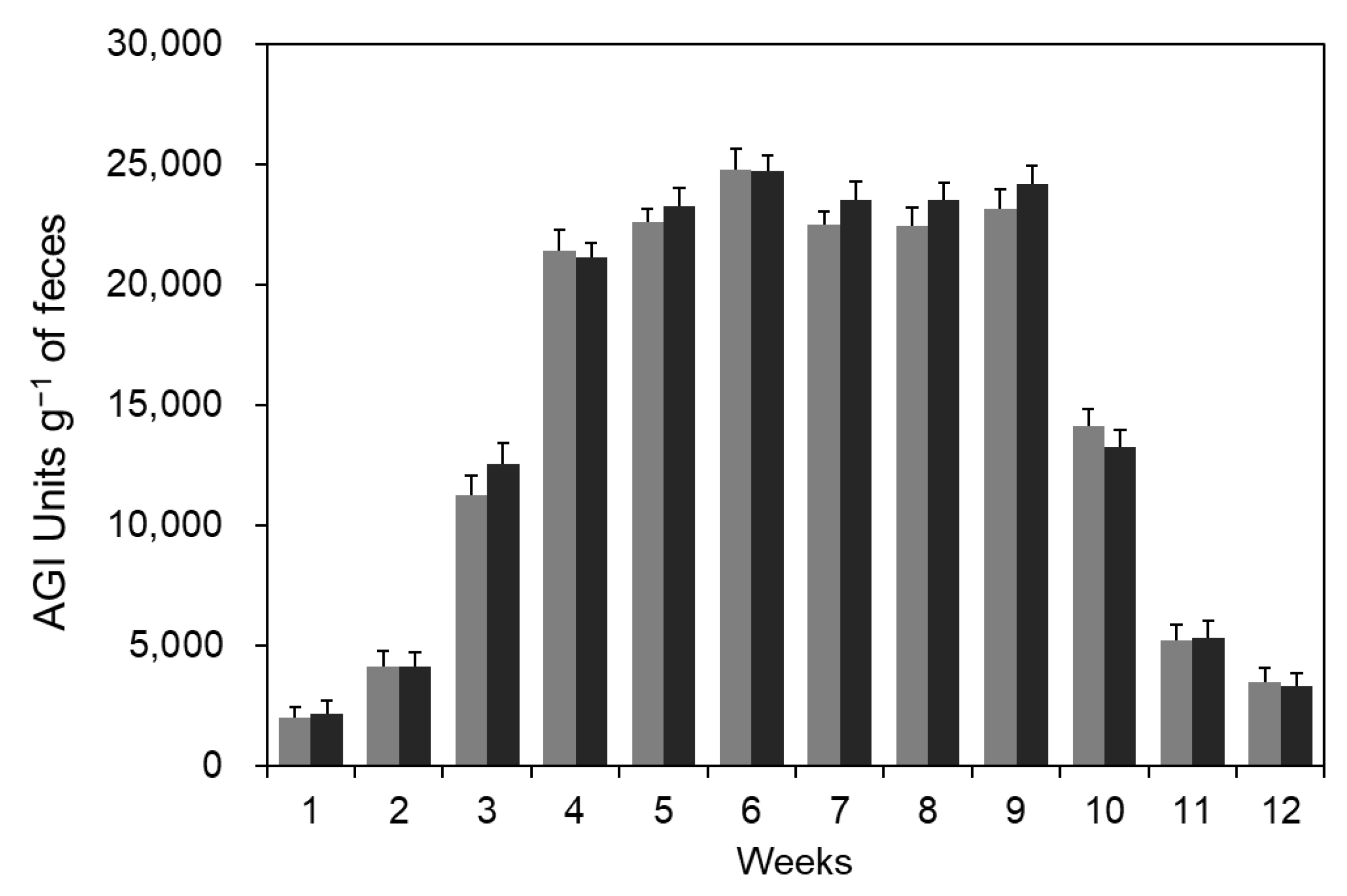
3.2. Production of AGI in Intestine by Spore Administration
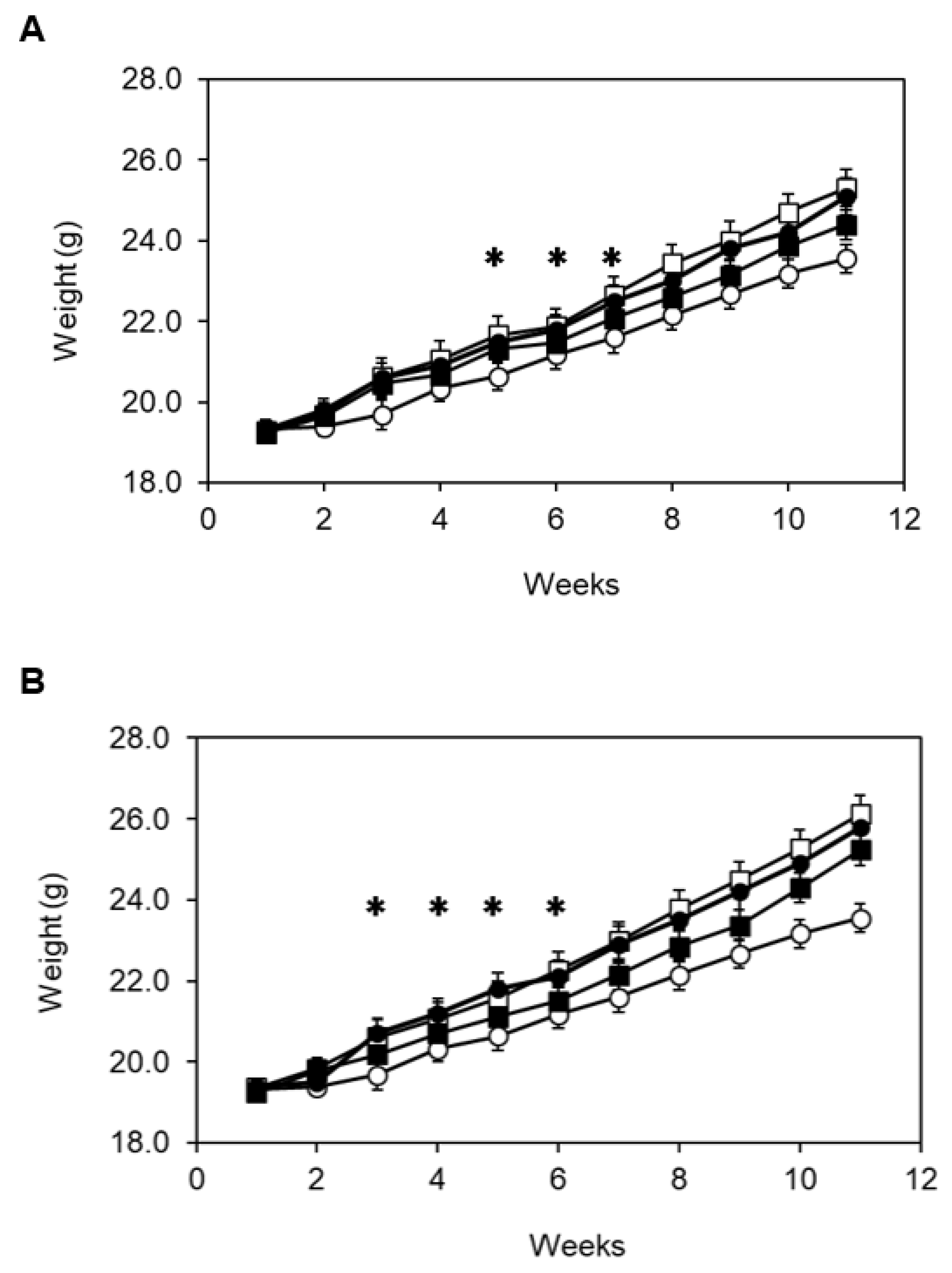
3.3. Spores Administration Effects against High-Calorie Diets
3.4. Feed Intake in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Steinert, R.E.; Lee, Y.K.; Sybesma, W. Vitamins for the gut microbiome. Trends Mol. Med. 2020, 26, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Uebanso, T.; Shimohata, T.; Mawatari, K.; Takahashi, A. Functional roles of B-vitamins in the gut and gut microbiome. Mol. Nutr. Food Res. 2020, 64, 2000426. [Google Scholar] [CrossRef]
- Wang, H.; Shou, Y.; Zhu, X.; Xu, Y.; Shi, L.; Xiang, S.; Feng, X.; Han, J. Stability of vitamin B12 with the protection of whey proteins and their effects on the gut microbiome. Food Chem. 2019, 276, 298–306. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Nochi, H. Physiological role of bile acids modified by the gut microbiome. Microorganisms 2021, 10, 68–84. [Google Scholar] [CrossRef]
- Zhu, C.; Miller, M.; Lusskin, N.; Bergk Pinto, B.; Maccario, L.; Häggblom, M.; Vogel, T.; LaRose, C.; Bromberg, Y. Snow microbiome functional analyses reveal novel aspects of microbial metabolism of complex organic compounds. Microbiologyopen 2020, 9, e1100. [Google Scholar] [CrossRef] [PubMed]
- Funabashi, M.; Grove, T.L.; Wang, M.; Varma, Y.; McFadden, M.E.; Brown, L.C.; Guo, C.; Higginbottom, S.; Almo, S.C.; Fischbach, M.A. A metabolic pathway for bile acid dehydroxylation by the gut microbiome. Nature 2020, 582, 566–570. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized metabolites from the microbiome in health and disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef]
- Hooper, L.V.; Dan, R.L.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15. [Google Scholar] [CrossRef]
- Riedl, R.A.; Atkinson, S.N.; Burnett, C.M.L.; Grobe, J.L.; Kirby, J.R. The gut microbiome, energy homeostasis, and implications for hypertension. Curr. Hypertens. Rep. 2017, 19, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, S.; Wang, Y.; Cai, S.; Yu, H.; Liu, H.; Zeng, X.; Zhang, G.; Qiao, S. Bridging intestinal immunity and gut microbiota by metabolites. Cell. Mol. Life Sci. 2019, 76, 3917–3937. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Yun, C.H.; Choi, Y.J.; Cho, C.S. Microencapsulation of live probiotic bacteria. J. Microbiol. Biotechnol. 2010, 20, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; van Hylckama Vlieg, J.E. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef]
- Henriques, S.F.; Dhakan, D.B.; Serra, L.; Francisco, A.P.; Carvalho-Santos, Z.; Baltazar, C.; Elias, A.P.; Anjos, M.; Zhang, T.; Maddocks, O.D.K.; et al. Metabolic cross-feeding in imbalanced diets allows gut microbes to improve reproduction and alter host behavior. Nat. Commun. 2020, 11, 4236–4250. [Google Scholar] [CrossRef]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef]
- Fares, C.; Menga, V.; Martina, A.; Pellegrini, N.; Scazzina, F.; Torriani, S. Nutritional profile and cooking quality of a new functional pasta naturally enriched in phenolic acids, added with β-glucan and Bacillus coagulans GBI-30, 6086. J. Cereal Sci. 2015, 65, 260–266. [Google Scholar] [CrossRef]
- Kim, H.W.; Lee, D.C.; Rhee, H.I. Production of α-glucosidase inhibitor in the intestines by Bacillus licheniformis. Enzyme. Microb. Technol. 2022, 158, 110032. [Google Scholar] [CrossRef]
- Park, M.H.; Ju, J.W.; Park, M.J.; Han, J.S. Daidzein inhibits carbohydrate digestive enzymes in vitro and alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2013, 712, 48–52. [Google Scholar] [CrossRef]
- Li, Y.; Ji, D.; Zhong, S.; Lin, T.; Lv, Z. Hypoglycemic effect of deoxynojirimycin–polysaccharide on high fat diet and streptozotocin-induced diabetic mice via regulation of hepatic glucose metabolism. Chem. Biol. Interact. 2015, 225, 70–79. [Google Scholar] [CrossRef]
- Wang, G.; Zhu, L.; Ma, M.; Chen, X.; Gao, Y.; Yu, T.; Yang, G.; Pang, W. Mulberry 1-deoxynojirimycin inhibits adipogenesis by repression of the ERK/PPARγ signaling pathway in porcine intramuscular adipocytes. J. Agric. Food Chem. 2015, 63, 6212–6220. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, J.Y.; Hwang, K.Y.; Cho, Y.S.; Park, Y.S.; Kang, K.D.; Seong, S.I. Isolation and identification of a Bacillus sp. producing α-glucosidase Inhibitor 1-deoxynojirimycin. Korean J. Microbiol. Biotechnol. 2011, 39, 49–55. [Google Scholar]
- Zhu, Y.P.; Li, X.T.; Teng, C.; Sun, B.G. Enhanced production of α-glucosidase inhibitor by a newly isolated strain of B. subtilis B2 using response surface methodology. Food Bioprod. Process. 2013, 91, 264–270. [Google Scholar] [CrossRef]
- Chen, J.; Cheng, Y.Q.; Yamaki, K.; Li, L.T. Anti-glucosidase activity of Chinese traditionally fermented soybean (douchi). Food Chem. 2007, 103, 1091–1096. [Google Scholar] [CrossRef]
- Fujita, H.; Yamagami, T.; Ohshima, K. Long-term ingestion of Touchi-extract, α-glucosidase inhibitor, by borderline and mild type-2 diabetic subjects is safe and significantly reduces blood glucose levels. Nutr. Res. 2003, 23, 713–722. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Yin, L.J.; Cheng, Y.Q.; Yamaki, K.; Mori, Y.; Su, Y.C.; Li, L.T. Effect of sources of carbon and nitrogen on production of α-glucosidase inhibitory by a newly isolated strain of B. subtilis B2. Food Chem. 2008, 109, 737–742. [Google Scholar] [CrossRef]
- Rombouts, J.L.; Kranendonk, E.M.M.; Regueira, A.; Weissbrodt, D.G.; Kleerebezem, R.; van Loosdrecht, M.C.M. Selecting for lactic acid producing and utilising bacteria in anaerobic enrichment cultures. Biotechnol. Bioeng. 2020, 117, 1281–1293. [Google Scholar] [CrossRef]
- Rhinehart, B.L.; Robinson, K.M.; Liu, P.S.; Payne, A.J.; Wheatley, M.E.; Wagner, S.R. Inhibition of intestinal disaccharidases and suppression of blood glucose by a new α-glucohydrolase inhibitor- MDL 25637. J. Pharmacol. Exp. Ther. 1987, 241, 915–920. [Google Scholar]
- Lim, C.S.; Li, C.Y.; Kim, Y.M.; Lee, W.Y.; Rhee, H.I. The inhibitory effect of Cornus walteri extract against α–amylase. J Korean Soc. Appl. Biol. Chem. 2005, 48, 103–108. [Google Scholar]
- Kim, Y.M.; Jeong, Y.K.; Wang, M.H.; Lee, W.Y.; Rhee, H.I. Inhibitory effect of pine extract on α-glucosidase activity and postprandial hyperglycemia. Nutrition 2005, 21, 756–761. [Google Scholar] [CrossRef]
- Uono, M.; Hacker, S.; Stephano, M.A.; Viana, V.C.; Manfrinato, C.V.; Bogsan, C.S.B. Technological development of biomass and the challenge of complete and efficient sporulation of Bacillus coagulans BVB5. Chem. Eng. Trans. 2019, 75, 457–462. [Google Scholar] [CrossRef]
- Wu, H.; Guo, Y.; Chen, L.; Chen, G.; Liang, Z. A novel strategy to regulate 1-deoxynojirimycin production based on its biosynthetic pathway in Streptomyces lavendulae. Front. Microbiol. 2019, 10, 1968. [Google Scholar] [CrossRef]
- Zhang, W.; Mu, W.; Wu, H.; Liang, Z. An overview of the biological production of 1-deoxynojirimycin: Current status and future perspective. Appl. Microbiol. Biotechnol. 2019, 103, 9335–9344. [Google Scholar] [CrossRef] [PubMed]
- Onose, S.; Ikeda, R.; Nakagawa, K.; Kimura, T.; Yamagishi, K.; Higuchi, O.; Miyazawa, T. Production of the α-glycosidase inhibitor 1-deoxynojirimycin from Bacillus species. Food Chem. 2013, 13, 516–523. [Google Scholar] [CrossRef]
- Thakur, K.; Zhang, Y.Y.; Mocan, A.; Zhang, F.; Zhang, J.G.; Wei, Z. 1-Deoxynojirimycin, its potential for management of non-communicable metabolic diseases. Trends Food Sci. Technol. 2019, 89, 88–99. [Google Scholar] [CrossRef]
- Wang, N.; Zhu, F.; Chen, K. 1-Deoxynojirimycin: Sources, extraction, analysis and biological functions. Nat. Prod. Commun. 2017, 12, 1521–1526. [Google Scholar] [CrossRef]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Potential use of Bacillus coagulans in the food industry. Foods 2018, 7, 92–102. [Google Scholar] [CrossRef]
- Schroedter, L.; Schneider, R.; Remus, L.; Venus, J. L-(+)-lactic acid from reed: Comparing various resources for the nutrient provision of B. coagulans. Resources 2020, 9, 89–100. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, Z.; Xu, Y.; Ying, J.; Wang, B.; Majeed, M.; Majeed, S.; Pande, A.; Li, W. Application of Bacillus coagulans in animal husbandry and its underlying mechanisms. Animals 2020, 10, 454–463. [Google Scholar] [CrossRef]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef]
- Majeed, M.; Majeed, S.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Ali, F. Evaluation of the stability of Bacillus coagulans MTCC 5856 during processing and storage of functional foods. Int. J. Food Sci. 2016, 51, 894–901. [Google Scholar] [CrossRef]
- Poudel, P.; Tashiro, Y.; Sakai, K. New application of Bacillus strains for optically pure L-lactic acid production: General overview and future prospects. Biosci. Biotechnol. Biochem. 2016, 80, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, A.; Keller, D.; Farmer, S. Survival and metabolic activity of the GanedenBC30 strain of Bacillus coagulans in a dynamic in vitro model of the stomach and small intestine. Benef. Microbes 2009, 1, 31–36. [Google Scholar] [CrossRef]
- Ramappa, V.K.; Srivastava, D.; Singh, P.; Kumar, U.; Singh, V. Mulberry 1-deoxynojirimycin (DNJ): An exemplary compound for therapeutics. J. Hortic. Sci. Biotechnol. 2020, 95, 679–686. [Google Scholar] [CrossRef]
- Tsuduki, T.; Kikuchi, I.; Kimura, T.; Nakagawa, K.; Miyazawa, T. Intake of mulberry 1-deoxynojirimycin prevents diet-induced obesity through increases in adiponectin in mice. Food Chem. 2013, 139, 16–23. [Google Scholar] [CrossRef]
- Lee, J.Y.; Aravinthan, A.; Park, Y.S.; Hwang, K.Y.; Seong, S.I.; Hwang, K. Supplementation of a fermented soybean extract reduces body mass and prevents obesity in high fat diet-induced c57bl/6j obese mice. Prev. Nutr. Food Sci. 2016, 21, 187–196. [Google Scholar] [CrossRef]
- Kojima, Y.; Kimura, T.; Nakagawa, K.; Asai, A.; Hasumi, K.; Oikawa, S.; Miyazawa, T. Effects of mulberry leaf extract rich in 1-deoxynojirimycin on blood lipid profiles in humans. J. Clin. Biochem. Nutr. 2010, 47, 155–161. [Google Scholar] [CrossRef]
- Forootanfar, H.; Faramarzi, M.A. Insights into laccase producing organisms, fermentation states, purification strategies, and biotechnological applications. Biotechnol. Prog. 2015, 31, 1443–1463. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Chotani, G.K.; Dodge, T.C.; Gaertner, A.L.; Arbige, M.V. Industrial biotechnology: Discovery to delivery. In Handbook of Industrial Chemistry and Biotechnology, 13th ed.; Kent, J.A., Bommaraju, T.V., Barnicki, S.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1495–1570. [Google Scholar]

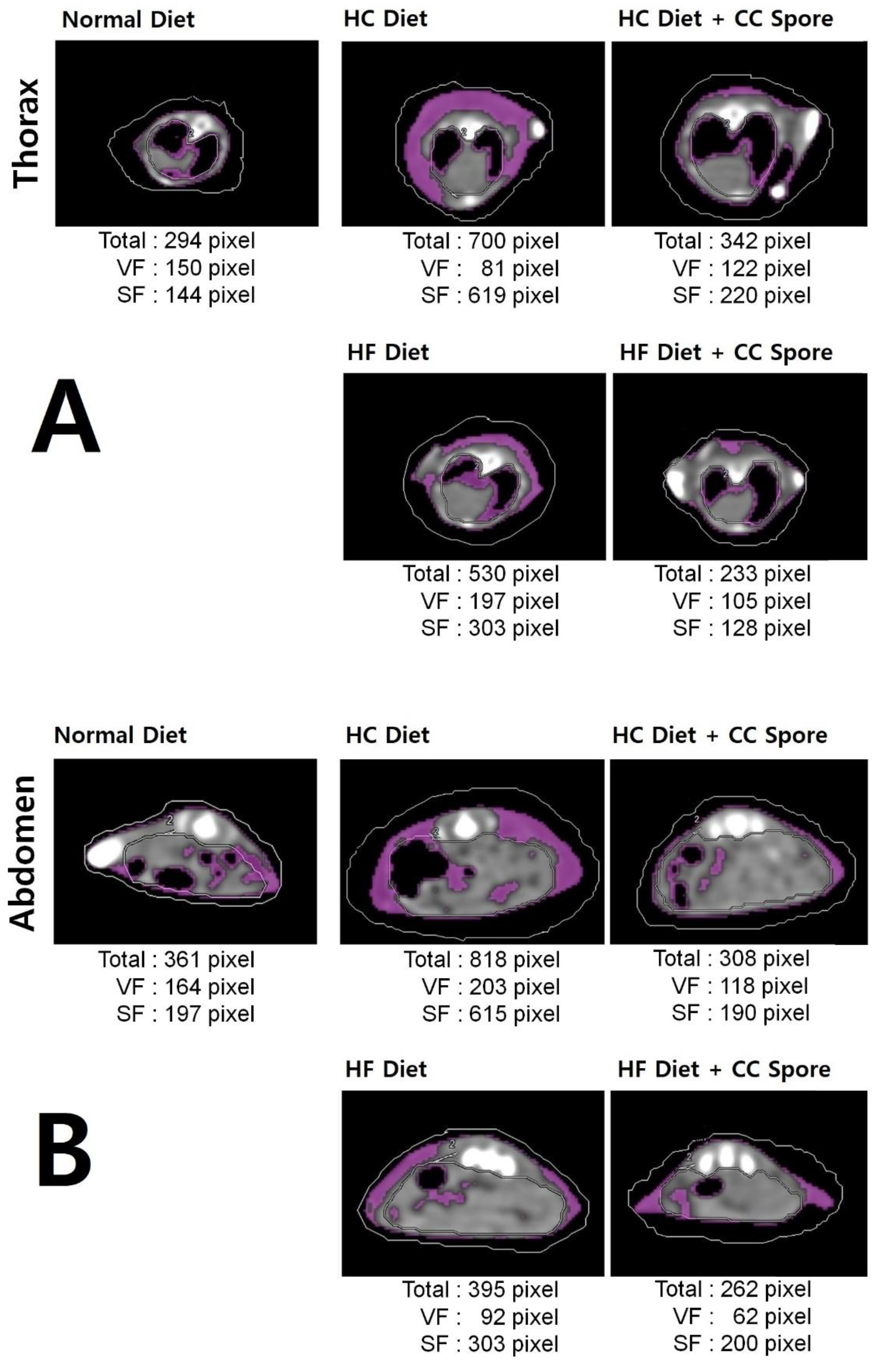
 ) and an HF diet with B. coagulans CC spores (
) and an HF diet with B. coagulans CC spores ( ). The arrow mark means the cessation of spore administration.
) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of spore administration.
). The arrow mark means the cessation of spore administration.
) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of spore administration. ) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of the diet point of B. coagulans CC spores.
) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of the diet point of B. coagulans CC spores.
) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of the diet point of B. coagulans CC spores.
) and an HF diet with B. coagulans CC spores (). The arrow mark means the cessation of the diet point of B. coagulans CC spores.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Incubation Time (Hours) | AGI (Unit mL−1) | |
|---|---|---|---|
| Aerobic | Anaerobic | ||
| B. subtilis DC-15 | 24 | 83,257 | N.D. a |
| B. coagulans ATCC7050 | 48 | N.D. a | N.D. a |
| B. coagulans CC | 48 | 19,478 | 12,879 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-W.; Choi, S.-Y.; Lee, D.-C.; Rhee, H.-I. Intestinal Production of Alpha-Glucosidase Inhibitor by Bacillus coagulans Spores. Microorganisms 2023, 11, 1462. https://doi.org/10.3390/microorganisms11061462
Kim H-W, Choi S-Y, Lee D-C, Rhee H-I. Intestinal Production of Alpha-Glucosidase Inhibitor by Bacillus coagulans Spores. Microorganisms. 2023; 11(6):1462. https://doi.org/10.3390/microorganisms11061462
Chicago/Turabian StyleKim, Hee-Woong, Soo-Young Choi, Deug-Chan Lee, and Hae-Ik Rhee. 2023. "Intestinal Production of Alpha-Glucosidase Inhibitor by Bacillus coagulans Spores" Microorganisms 11, no. 6: 1462. https://doi.org/10.3390/microorganisms11061462