
Isolation of Bacteria from Freeze-Dried Samples and the Functional Characterization of Species-Specific Lactic Acid Bacteria with a Comparison of Wild and Captive Proboscis Monkeys

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bacteria from Freeze-Dried Samples of Wild, Semi-Wild, or Captive Proboscis Monkeys
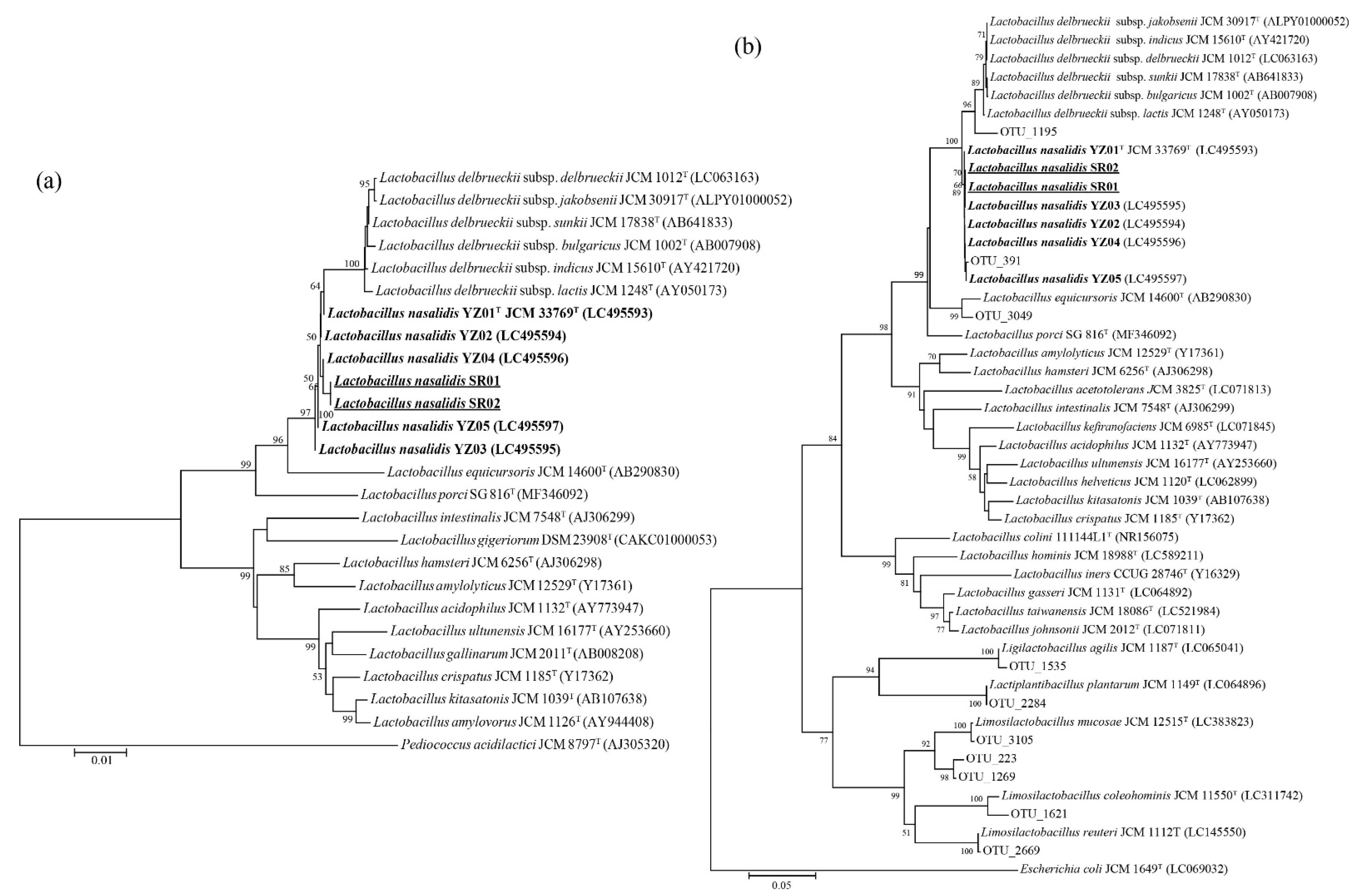
2.2. Phylogenetic Analysis of the 16S rRNA Gene
2.3. Molecular Confirmation of the Presence of L. nasalidis in the Previous 16S Metagenomic DNA Library
2.4. Phenotypic Characterization of Isolated Bacteria
3. Results and Discussions
3.1. Isolation of L. nasalidis from the Freeze-Dried Samples of a Wild Proboscis Monkey
3.2. Comparison of L. nasalidis Proportion between Different Habitat Environments in the 16S DNA Library
3.3. Comparison of Phenotype Characteristics between Captive and Wild Isolates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 42, 15611–15616. [Google Scholar] [CrossRef]
- Perdigón, G.; Fuller, R.; Raya, R. Lactic acid bacteria and their effect on the immune system. Curr. Issues Intest. Microbiol. 2001, 2, 27–42. [Google Scholar] [PubMed]
- Majamaa, H.; Isolauri, E. Probiotics: A novel approach in the management of food allergy. J. Allergy Clin. Immunol. 1997, 99, 179–185. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Narbad, A.; Gasson, M.J.; Woodward, M.J. In vivo characterization of Lactobacillus johnsonii FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry. Lett. Appl. Microbiol. 2004, 38, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Watkins, C.; Stanton, C.; Ross, R.P. Gut bifidobacteria populations in human health and aging. Front. Microbiol. 2016, 7, 1204. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef]
- Endo, A.; Futagawa-Endo, Y.; Dicks, L.M.T. Diversity of Lactobacillus and Bifidobacterium in feces of herbivores, omnivores and carnivores. Anaerobe 2010, 16, 590–596. [Google Scholar] [CrossRef]
- Hanning, I.; Diaz-Sanchez, S. The functionality of the gastrointestinal microbiome in non-human animals. Microbiome 2015, 3, 1–11. [Google Scholar] [CrossRef]
- Pasolli, E.; De Filippis, F.; Mauriello, I.E.; Cumbo, F.; Walsh, A.M.; Leech, J.; Cotter, P.D.; Segata, N.; Ercolini, D. Large-scale genome-wide analysis links lactic acid bacteria from food with the gut microbiome. Nat. Commun. 2020, 11, 2610. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, S.; Kakooza, S.; Nguema, P.P.M.; Wampande, E.M.; Ushida, K. Characteristics of gorilla-specific Lactobacillus isolated from captive and wild gorillas. Microorganisms 2018, 6, 86. [Google Scholar] [CrossRef]
- Mbehang, P.P.N.; Tsuchida, S.; Ushida, K. Bacteria culturing and isolation under field conditions of Moukalaba-Doudou National Park, Gabon, and preliminary survey on bacteria carrying antibiotic resistance genes. Tropics 2015, 23, 165–174. [Google Scholar] [CrossRef]
- Suzuki-Hashido, N.; Tsuchida, S.; Hayakawa, T.; Sakamoto, M.; Azumano, A.; Seino, S.; Matsuda, I.; Ohkuma, M.; Ushida, K. Lactobacillus nasalidis sp. nov., isolated from the forestomach of a captive proboscis monkey (Nasalis larvatus). Int. J. Syst. Evol. Microbiol. 2021, 71, 4787. [Google Scholar] [CrossRef] [PubMed]
- Boonratana, R. Feeding ecology of proboscis monkeys (Nasalis larvatus) in the lower Kinabatangan, Sabah, Malaysia. Sabah Parks Nat. J. 2003, 6, 1–26. [Google Scholar]
- Chivers, D.J. Functional anatomy of the gastrointestinal tract. In Colobine Monkeys: Their Ecology, Behaviour and Evolution; Davies, A.G., Oates, J.F., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 205–257. [Google Scholar]
- Lambert, J.E. Primate digestion: Interactions among anatomy, physiology, and feeding ecology. Evol. Anthropol. 1998, 7, 8–20. [Google Scholar] [CrossRef]
- Kay, R.N.B.; Davies, A.G. Digestive physiology. In Colobine Monkeys: Their Ecology, Behaviour and Evolution; Davies, A.G., Oates, J.F., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 229–259. [Google Scholar]
- Hayakawa, T.; Nathan, S.K.S.S.; Stark, D.J.; Saldivar, D.A.R.; Sipangkui, R.; Goossens, B.; Tuuga, A.; Clauss, M.; Sawada, A.; Fukuda, S.; et al. First report of foregut microbial community in proboscis monkeys: Are diverse forests a reservoir for diverse microbiomes? Environ. Microbiol. Rep. 2018, 10, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Amato, K.; Hou, R.; Gomez, A.; Dunn, D.W.; Zhang, J.; Garber, P.A.; Chapman, C.A.; Righini, N.; He, G.; et al. Specialized digestive adaptations within the hindgut of a colobine monkey. Innovation 2022, 3, 100207. [Google Scholar] [CrossRef]
- Bauchop, T.; Martucci, R.W. Ruminant-like digestion of the langur monkey. Science 1968, 161, 698–700. [Google Scholar] [CrossRef]
- Matsuda, I.; Murai, T.; Clauss, M.; Yamada, T.; Tuuga, A.; Bernard, H.; Higashi, S. Regurgitation and remastication in the foregut-fermenting proboscis monkey (Nasalis larvatus). Biol. Lett. 2011, 7, 786–789. [Google Scholar] [CrossRef]
- Tsukahara, T.; Ushida, K. Succinate accumulation in pig large intestine during antibiotic-associated diarrhea and the constitution of succinate-producing flora. J. Gen. Appl. Microbiol. 2002, 48, 143–154. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Deane, J.; Rea, M.C.; O’Sullivan, Ó.; Ross, R.P.; O’Callaghan, G.; Plant, B.J.; Stanton, C. The Effects of Freezing on Faecal Microbiota as Determined Using MiSeq Sequencing and Culture-Based Investigations. PLoS ONE 2015, 10, e0119355. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- Hayakawa, T.; Sawada, A.; Tanabe, A.S.; Fukuda, S.; Kishida, T.; Kurihara, Y.; Matsushima, K.; Liu, J.; Akomo-Okoue, E.F.; Gravena, W.; et al. Improving the standards for gut microbiome analysis of fecal samples: Insights from the field biology of Japanese macaques on Yakushima Island. Primates 2018, 59, 423–436. [Google Scholar] [CrossRef]
- Lewis, Z.T.; Davis, J.C.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B.; Mills, D.A. The impact of freeze-drying infant fecal samples on measures of their bacterial community profiles and milk-derived oligosaccharide content. PeerJ 2016, 4, e1612. [Google Scholar] [CrossRef]
- Miyamoto-Shinohara, Y.; Imaizumi, T.; Sukenobe, J.; Murakami, Y.; Kawamura, S.; Komatsu, Y. Survival rate of microbes after freeze-drying and long-term storage. Cryobiology 2000, 41, 251–255. [Google Scholar] [CrossRef]
- Matsuda, I.; Nakabayashi, M.; Otani, Y.; Yap, S.W.; Tuuga, A.; Wong, A.; Bernard, H.; Wich, S.A.; Kubo, T. Comparison of plant diversity and phenology of riverine and mangrove forests with those of the dryland forest in Sabah, Borneo, Malaysia. In Primates in Flooded Habitats: Ecology and Conservation; Nowak, K., Barnett, A.A., Matsuda, I., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 15–28. [Google Scholar]
- Matsuda, I.; Sha, J.C.M.; Bernard, H. Proboscis Monkey. In Wildlife Atlas of Sabah: Maps and Conservation; Davies, G., Ed.; WWF-Malaysia: Selangor, Malaysia, 2022; pp. 77–84. [Google Scholar]
- Clayton, J.B.; Al-Ghalith, G.A.; Long, H.T.; Tuan, B.V.; Cabana, F.; Huang, H.; Vangay, P.; Ward, T.; Minh, V.V.; Tam, N.A.; et al. Associations between nutrition, gut microbiome, and health in a novel nonhuman primate model. Sci. Rep. 2018, 8, 11159. [Google Scholar] [CrossRef]
- Tsuchida, S.; Maruyama, F.; Ogura, Y.; Toyoda, A.; Hayashi, T.; Okuma, M.; Ushida, K. Genomic characteristics of Bifidobacterium thermacidophilum pig isolates and wild boar isolates reveal the unique presence of a putative mobile genetic element with tetW for pig farm isolates. Front. Microbiol. 2017, 8, 1540. [Google Scholar] [CrossRef]
- Ushida, K.; Tsuchida, S.; Ogura, Y.; Toyoda, A.; Maruyama, F. Domestication and cereal feeding developed domestic pig-type intestinal microbiota in animals of suidae. Anim. Sci. J. 2016, 87, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Hayakawa, T.; Kiyono, M.; Yamabata, N.; Hanya, G. Gut microbiota composition of Japanese macaques associates with extent of human encroachment. Am. J. Primatol. 2019, 81, e23072. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Zhang, L.; Xie, H.W.; Simmons, N.B.; Liu, H.; Zhao, H. Trehalase gene as a molecular signature of dietary diversification in mammals. Mol. Biol. Evol. 2019, 36, 2171–2183. [Google Scholar] [CrossRef]
- Jaszczak-Wilke, E.; Polkowska, Ż.; Koprowski, M.; Owsianik, K.; Mitchell, A.E.; Bałczewski, P. Amygdalin: Toxicity, anticancer activity and analytical procedures for its determination in plant seeds. Molecules 2021, 26, 2253. [Google Scholar] [CrossRef]
- Waterman, P.G.; Kool, K.M. Colobine food selection and plant chemistry. In Colobine Monkeys: Their Ecology, Behaviour and Evolution; Davies, A.G., Oates, J.F., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 251–284. [Google Scholar]
- Tsuchida, S.; Murata, K.; Ohkuma, M.; Ushida, K. Isolation of Streptococcus gallolyticus with very high degradability of condensed tannins from feces of the wild Japanese rock ptarmigans on Mt. Tateyama. J. Gen. Appl. Microbiol. 2017, 63, 195–198. [Google Scholar] [CrossRef]
- Tsuchida, S.; Ohara, Y.; Kuramochi, K.; Murata, K.; Ushida, K. Effective degradation of phenolic glycoside rhododendron and its aglycone rhododendron by feces of wild Japanese rock ptarmigans. J. Zoo Wildl. Med. 2017, 22, 41–45. [Google Scholar] [CrossRef]
- Hoshino, S.; Seino, S.; Funahashi, T.; Hoshino, T.; Clauss, M.; Matsuda, I.; Yayota, M. Apparent diet digestibility of captive colobines in relation to stomach types with special reference to fibre digestion. PLoS ONE 2021, 16, e0256548. [Google Scholar] [CrossRef] [PubMed]
- Yeager, C.P.; Silver, S.C.; Dierenfeld, E.S. Mineral and phytochemical influences on foliage selection by the proboscis monkey (Nasalis larvatus). Am. J. Primatol. 1997, 41, 117–128. [Google Scholar]
- Agoramoorthy, G.; Alagappasamy, C.; Hsu, M.J. Can proboscis monkeys be successfully maintained in captivity? A case of swings and roundabouts. Zoo Biol. 2004, 23, 533–544. [Google Scholar] [CrossRef]
- Agoramoorthy, G.; Hsu, M.J. Borneo’s proboscis monkey–a study of its diet of mineral and phytochemical concentrations. Curr. Sci. 2005, 89, 454–457. [Google Scholar]
- Clayton, J.B.; Shields-Cutler, R.R.; Hoops, S.L.; Al-Ghalith, G.A.; Sha, J.C.; Johnson, T.J.; Knights, D. Bacterial community structure and function distinguish gut sites in captive red-shanked doucs (Pygathrix nemaeus). Am. J. Primatol. 2019, 81, e22977. [Google Scholar] [CrossRef] [PubMed]
- Huan, Z.; Yao, Y.; Yu, J.; Chen, H.; Li, M.; Yang, C.; Zhao, B.; Ni, Q.; Zhang, M.; Xie, M.; et al. Differences in the gut microbiota between Cercopithecinae and Colobinae in captivity. J. Microbiol. 2020, 58, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, I.; Clauss, M. Morphology and physiology of colobine digestive tracts. In The Colobines: Natural History, Behaviour and Ecological Diversity; Matsuda, I., Grueter, C.C., Teichroeb, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 64–77. [Google Scholar]


{kind=link}
| OUT_ID | Related Species | Similarity to Type Strain | Wild | Wild | Semi-Wild | Captive | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Riverine Forest | Mangrove Forest | Mangrove Forest | Malaysian Zoo | |||||||
| Number a | Proportion b | Number a | Proportion b | Number a | Proportion b | Number a | Proportion b | |||
| OUT_391 | Lactobacillus nasalidis | 100.0 | 8 | 100.0 | 0 | - | 277 | 89.9 | 148 | 12.7 |
| OUT_1195 | Lactobacillus delbrueckii subsp. lactis | 97.8 | 0 | 0.0 | 0 | - | 27 | 8.8 | 0 | 0.0 |
| OUT_3049 | Lactobacillus equicursoris | 97.9 | 0 | 0.0 | 0 | - | 0 | 0.0 | 217 | 18.6 |
| OUT_2284 | Lactobacillus plantarum | 100.0 | 0 | 0.0 | 0 | - | 1 | 0.3 | 0 | 0.0 |
| OUT_1621 | Lactobacillus coleohominis | 97.6 | 0 | 0.0 | 0 | - | 0 | 0.0 | 6 | 0.5 |
| OUT_223 | Lactobacillus mucosae | 95.2 | 0 | 0.0 | 0 | - | 0 | 0.0 | 575 | 49.4 |
| OUT_3105 | Lactobacillus mucosae | 99.7 | 0 | 0.0 | 0 | - | 3 | 1.0 | 164 | 14.1 |
| OUT_1269 | Lactobacillus mucosae | 95.7 | 0 | 0.0 | 0 | - | 0 | 0.0 | 35 | 3.0 |
| OUT_2669 | Lactobacillus reuteri | 100.0 | 0 | 0.0 | 0 | - | 0 | 0.0 | 19 | 1.6 |
| OUT_1535 | Lactobacillus agilis | 99.4 | 0 | 0.0 | 0 | - | 0 | 0.0 | 1 | 0.1 |
| Subtotal reads for Lactobacillaceae | 8 | 0 | 308 | 1165 | ||||||
| Total reads c | 40,809 | 40,809 | 40,809 | 40,809 | ||||||
| L. nasalidis | L. delbrueckii subsp. indicus | L. equicursoris | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Isolated from | Wild Proboscis Monkey | Captive Proboscis Monkey | Indian Dairy Products | Thoroughbred Racehorse | |||||
| SR01 | SR02 | YZ01T * | YZ02 * | YZ03 * | YZ04 * | YZ05 * | JCM 15610T * | JCM 14600T * | |
| Acid production from (API 50 CHL) | |||||||||
| D-Galactose | − | + | − | − | − | − | − | − | + |
| D-Mannose | − | + | + | + | + | + | + | + | + |
| Amygdalin | + | + | + | + | + | + | + | − | W |
| Arbutin | + | + | + | + | + | + | + | − | + |
| Esculin ferric citrate | + | W | + | + | + | + | + | + | + |
| Salicin | + | + | + | + | + | + | + | − | + |
| D-Cellobiose | + | + | + | + | + | + | + | − | + |
| D-Maltose | + | + | + | + | + | + | + | − | + |
| D-Lactose | + | + | + | + | W | − | W | + | + |
| D-Melibiose | − | − | − | − | − | − | − | − | W |
| D-Trehalose | + | + | + | + | + | + | + | − | − |
| D-Raffinose | W | W | − | − | − | − | − | − | W |
| Starch | − | − | − | − | − | − | − | − | W |
| Gentiobiose | + | + | + | + | W | + | W | − | + |
| API ZYM results: | |||||||||
| Phosphate alkaline | − | − | − | − | − | W | W | − | − |
| Esterase (C 4) | W | − | W | W | W | W | W | W | W |
| Esterase lipase (C 8) | − | − | W | W | W | W | − | W | W |
| Lipase (C 14) | − | − | − | − | − | W | − | − | − |
| Leucine aminopeptidase | + | + | W | + | W | + | + | + | + |
| Valine aminopeptidase | − | − | − | W | W | W | W | W | − |
| Cystine aminopeptidase | W | W | − | W | − | W | − | W | W |
| Trypsin | − | − | − | − | − | − | − | − | W |
| Chymotrypsin | W | W | − | − | − | W | − | W | W |
| Phosphatase acid | W | W | + | + | + | + | + | + | W |
| Naphthol-AS-BI-phosphohydrolase | + | W | + | + | + | + | + | + | + |
| α-Galactosidase | − | − | − | − | − | − | − | W | + |
| β-Galactosidase | + | + | + | + | W | W | W | + | + |
| α-Glucosidase | W | W | W | W | W | W | W | − | + |
| α-Glucosidase | + | + | W | + | + | + | + | W | W |
| Growth at: | |||||||||
| 15 °C | W | − | − | − | W | W | W | W | W |
| 14 °C | + | + | + | + | + | + | + | + | + |
| Growth with NaCl (w/v) | |||||||||
| 4.0% | + | + | + | + | + | + | + | + | − |
| 5.0% | − | − | + | + | − | − | + | − | − |
| 6.0% | − | − | − | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki-Hashido, N.; Tsuchida, S.; Azumano, A.; Goossens, B.; Saldivar, D.A.R.; Stark, D.J.; Tuuga, A.; Ushida, K.; Matsuda, I. Isolation of Bacteria from Freeze-Dried Samples and the Functional Characterization of Species-Specific Lactic Acid Bacteria with a Comparison of Wild and Captive Proboscis Monkeys. Microorganisms 2023, 11, 1458. https://doi.org/10.3390/microorganisms11061458
Suzuki-Hashido N, Tsuchida S, Azumano A, Goossens B, Saldivar DAR, Stark DJ, Tuuga A, Ushida K, Matsuda I. Isolation of Bacteria from Freeze-Dried Samples and the Functional Characterization of Species-Specific Lactic Acid Bacteria with a Comparison of Wild and Captive Proboscis Monkeys. Microorganisms. 2023; 11(6):1458. https://doi.org/10.3390/microorganisms11061458
Chicago/Turabian StyleSuzuki-Hashido, Nami, Sayaka Tsuchida, Akinori Azumano, Benoit Goossens, Diana A. Ramirez Saldivar, Danica J. Stark, Augustine Tuuga, Kazunari Ushida, and Ikki Matsuda. 2023. "Isolation of Bacteria from Freeze-Dried Samples and the Functional Characterization of Species-Specific Lactic Acid Bacteria with a Comparison of Wild and Captive Proboscis Monkeys" Microorganisms 11, no. 6: 1458. https://doi.org/10.3390/microorganisms11061458