
Gausemycin A-Resistant Staphylococcus aureus Demonstrates Affected Cell Membrane and Cell Wall Homeostasis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Minimum Inhibitory Concentration
2.3. Spa Typing and MLST
2.4. RNA Extraction and Reverse Transcription into cDNA
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
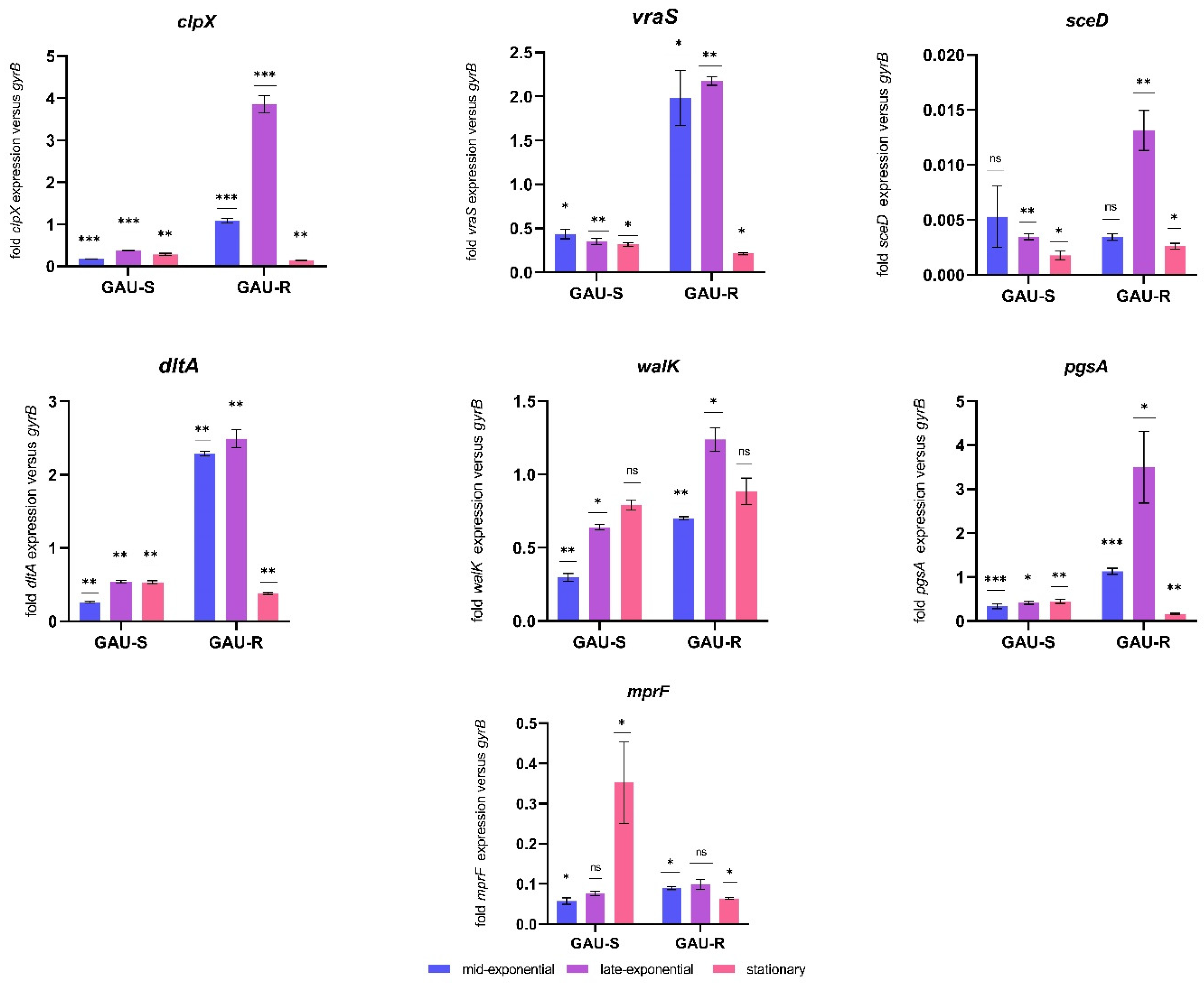
3.1. Comparison of Gene Expression between the GAU-S and GAU-R Strains
3.2. The Gene Expression Depending on Growth Phase
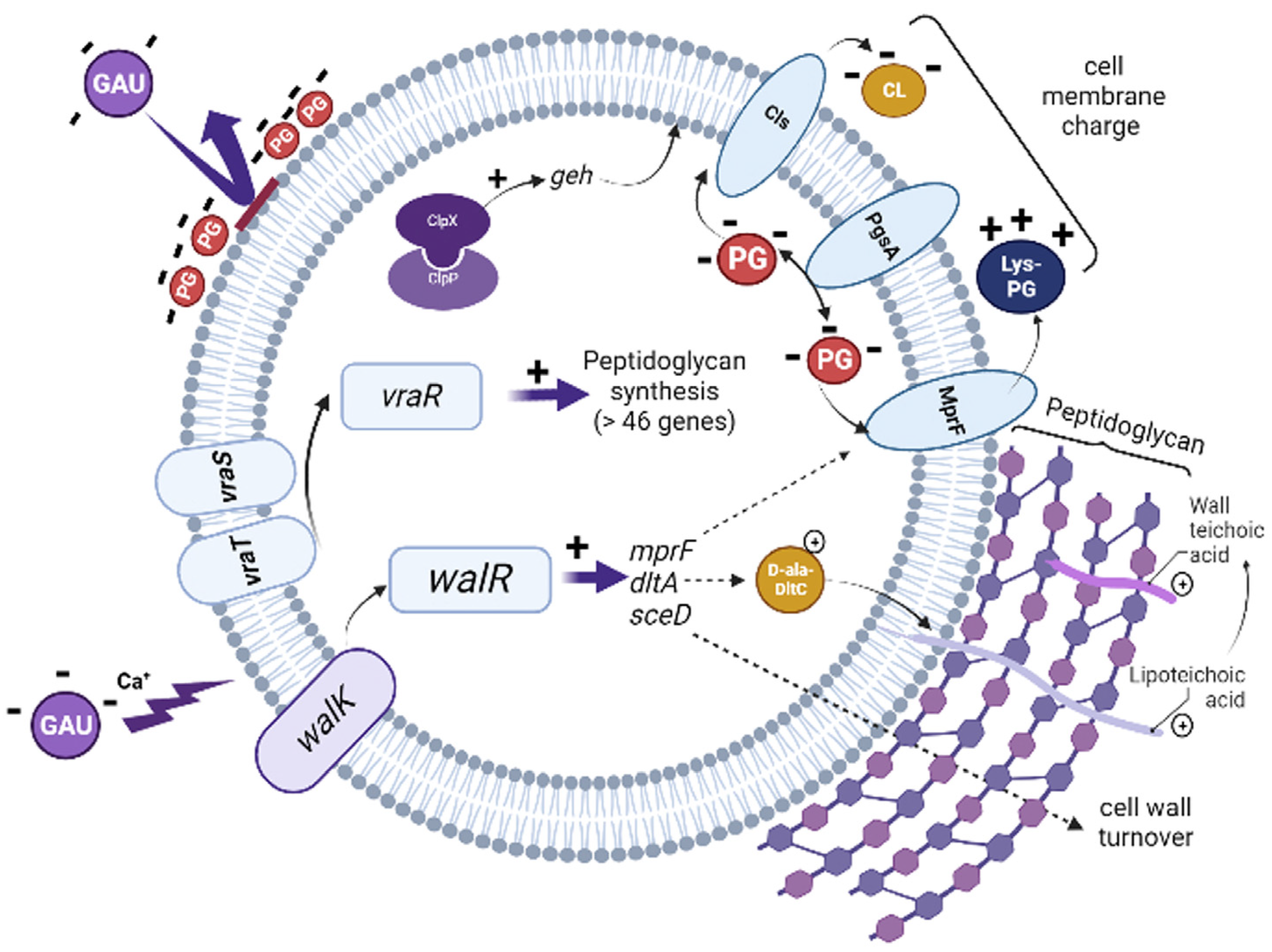
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The antimicrobial resistance crisis: Causes, consequences, and management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef]
- Lade, H.; Kim, J.-S. Bacterial Targets of Antibiotics in Methicillin-Resistant Staphylococcus aureus. Antibiotics 2021, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Casanova, N.G.; Ruiz, M.S.; Bellido, J.L.M. Mechanisms of resistance to daptomycin in Staphylococcus aureus. Rev. Esp. Quimioter. 2017, 30, 391–396. [Google Scholar]
- Tyurin, A.P.; Alferova, V.A.; Paramonov, A.S.; Shuvalov, M.V.; Kudryakova, G.K.; Rogozhin, E.A.; Zherebker, A.Y.; Brylev, V.A.; Chistov, A.A.; Baranova, A.A.; et al. Gausemycins A, B: Cyclic lipoglycopeptides from Streptomyces sp. Angew. Chem. Int. Ed. 2021, 60, 18694–18703. [Google Scholar] [CrossRef]
- Vasilchenko, A.S.; Julian, W.T.; Lapchinskaya, O.A.; Katrukha, G.S.; Sadykova, V.S.; Rogozhin, E.A. A Novel Peptide Antibiotic Produced by Streptomyces roseoflavus Strain INA-Ac-5812 with Directed Activity against Gram-Positive Bacteria. Front. Microbiol. 2020, 11, 556063. [Google Scholar] [CrossRef] [PubMed]
- Kravchenko, T.V.; Paramonov, A.S.; Kudzhaev, A.M.; Efimova, S.S.; Khorev, A.S.; Kudryakova, G.K.; Ivanov, I.A.; Chistov, A.A.; Baranova, A.A.; Krasilnikov, M.S.; et al. Gausemycin antibiotic family act via Ca2+-dependent membrane targeting. In ChemRxiv; Cambridge Open Engage: Cambridge, UK, 2022. [Google Scholar]
- Poshvina, D.V.; Dilbaryan, D.S.; Kasyanov, S.P.; Sadykova, V.S.; Lapchinskaya, O.A.; Rogozhin, E.A.; Vasilchenko, A.S. Staphylococcus aureus is able to generate resistance to novel lipoglycopeptide antibiotic gausemycin A. Front. Microbiol. 2022, 13, 963979. [Google Scholar] [CrossRef]
- Ernst, C.M.; Staubitz, P.; Mishra, N.N.; Yang, S.-J.; Hornig, G.; Kalbacher, H.; Bayer, A.S.; Kraus, D.; Peschel, A. The bacterial defensin resistance protein mprf consists of separable domains for lipid lysinylation and antimicrobial peptide repulsion. PLoS Pathog. 2009, 5, e1000660. [Google Scholar] [CrossRef]
- Oshida, T.; Sugai, M.; Komatsuzawa, H.; Hong, Y.M.; Suginaka, H.; Tomasz, A. A Staphylococcus aureus autolysin that has an Nacetylmuramoyl-L-alanine amidase domain and an endo-beta-N-acetylglucosaminidase domain: Cloning, sequence analysis, and characterization. Proc. Natl. Acad. Sci. USA 1995, 92, 285–289. [Google Scholar] [CrossRef]
- Drummelsmith, J.; Winstall, E.; Bergeron, M.G.; Poirier, G.G.; Ouellette, M. Comparative proteomics analyses reveal a potential biomarker for the detection of vancomycin-intermediate Staphylococcus aureus strains. J. Proteome Res. 2007, 6, 4690–4702. [Google Scholar] [CrossRef]
- Tran, T.T.; Munita, J.M.; Arias, C.A. Mechanisms of drug resistance: Daptomycin resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 32–53. [Google Scholar] [CrossRef]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-J.; Xiong, Y.Q.; Dunman, P.M.; Schrenzel, J.; François, P.; Peschel, A.; Bayer, A.S. Regulation of mprF in daptomycin-nonsusceptible Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2009, 53, 2636–2637. [Google Scholar] [CrossRef] [PubMed]
- Sabat, A.J.; Tinelli, M.; Grundmann, H.; Akkerboom, V.; Monaco, M.; Del Grosso, M.; Errico, G.; Pantosti, A.; Friedrich, A.W. Daptomycin resistant Staphylococcus aureus clinical strain with novel non-synonymous mutations in the mprF and vraS genes: A new insight into daptomycin resistance. Front. Microbiol. 2018, 9, 2705. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Yang, S.J.; Bayer, A.S.; Vaezzadeh, A.R.; Herzig, S.; Stenz, L.; Girard, M.; Sakoulas, G.; Scherl, A.; Yeaman, M.R.; et al. Daptomycin resistance mechanisms in clinically derived Staphylococcus aureus strains assessed by a combined transcriptomics and proteomics approach. J. Antimicrob. Chemother. 2011, 66, 1696–1711. [Google Scholar] [CrossRef] [PubMed]
- Bæk, K.T.; Gründling, A.; Mogensen, R.G.; Thøgersen, L.; Petersen, A.; Paulander, W.; Frees, D. β-Lactam resistance in methicillin-resistant Staphylococcus aureus USA300 is increased by inactivation of the ClpXP protease. Antimicrob. Agents Chemother. 2014, 58, 4593–4603. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, V.C.; Fetzer, C.; Sieber, S.A. Global inventory of ClpP- and ClpX-regulated proteins in Staphylococcus aureus. J. Proteome Res. 2021, 20, 867–879. [Google Scholar] [CrossRef]
- Aires-de-Sousa, M.; Boye, K.; de Lencastre, H.; Deplano, A.; Enright, M.C.; Etienne, J.; Friedrich, A.; Harmsen, D.; Holmes, A.; Huijsdens, X.W.; et al. High interlaboratory reproducibility of DNA sequence based typing of bacteria in a multicenter study. J. Clin. Microbiol. 2006, 44, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef]
- Pfaffl, M.W.A. New mathematical model for relative quantification in real-time RT—PCR. Nucleic Acids Res. 2001, 29, 16–21. [Google Scholar] [CrossRef]
- Kuroda, M.; Sekizuka, T.; Matsui, H.; Ohsuga, J.; Ohshima, T.; Hanaki, H. IS256-Mediated Overexpression of the WalKR Two-Component System Regulon Contributes to Reduced Vancomycin Susceptibility in a Staphylococcus aureus Clinical Isolate. Front. Microbiol. 2019, 10, 1882. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Spina, D.; Purrello, S.; Campanile, F.; Di Pietro, C.; Purrello, M.; Stefani, S. Modulating activity of vancomycin and daptomycin on the expression of autolysis cell-wall turnover and membrane charge genes in hVISA and VISA strains. PLoS ONE 2012, 7, e29573. [Google Scholar] [CrossRef]
- Ji, Q.; Chen, P.; Qin, G.; Deng, X.; Hao, Z.; Wawrzak, Z.; Yeo, W.S.; Quang, J.W.; Cho, H.; Luo, G.Z.; et al. Structure and mechanism of the essential two-component signal-transduction system WalKR in Staphylococcus aureus. Nat. Commun. 2016, 7, 11000. [Google Scholar] [CrossRef]
- Utaida, S.; Dunman, P.M.; Macapagal, D.; Murphy, E.; Projan, S.J.; Singh, V.K.; Jayaswal, R.K.; Wilkinson, B.J. Genome-wide transcriptional profiling of the response of Staphylococcus aureus to cell-wall active antibiotics reveals a cell-wall-stress stimulon. Microbiology 2003, 149, 2719–2732. [Google Scholar] [CrossRef]
- McCallum, N.; Spehar, G.; Bischoff, M.; Berger-Bachi, B. Strain dependence of the cell wall-damage induced stimulon in Staphylococcus aureus. Biochim. Biophys. Acta 2006, 1760, 1475–1481. [Google Scholar] [CrossRef]
- Utaida, S.; Pfeltz, R.F.; Jayaswal, R.K.; Wilkinson, B.J. Autolytic properties of glycopeptide-intermediate Staphylococcus aureus Mu50. Antimicrob. Agents Chemother. 2006, 50, 1541–1545. [Google Scholar] [CrossRef]
- Dubrac, S.; Boneca, I.G.; Poupel, O.; Msadek, T. New insights into the WalK/WalR (YycG/YycF) essential signal transduction pathway reveal a major role in controlling cell wall metabolism and biofilm formation in Staphylococcus aureus. J. Bacteriol. 2007, 189, 8257–8269. [Google Scholar] [CrossRef]
- Howden, B.P.; McEvoy, C.R.E.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; et al. Evolution of Multidrug Resistance during Staphylococcus aureus Infection Involves Mutation of the Essential Two Component Regulator WalKR. PLoS Pathog. 2011, 7, e1002359. [Google Scholar] [CrossRef]
- McEvoy, C.R.; Tsuji, B.; Gao, W.; Seemann, T.; Porter, J.L.; Doig, K.; Ngo, D.; Howden, B.P.; Stinear, T.P. Decreased vancomycin susceptibility in Staphylococcus aureus caused by IS256 tempering of WalKR expression. Antimicrob. Agents Chemother. 2013, 57, 3240–3249. [Google Scholar] [CrossRef]
- Bertsche, U.; Yang, S.-J.; Kuehner, D.; Wanner, S.; Mishra, N.N.; Roth, T.; Nega, M.; Schneider, A.; Mayer, C.; Grau, T.; et al. Increased cell wall teichoic acid production and D-alanylation are common phenotypes among daptomycin-resistant methicillin-resistant Staphylococcus aureus (MRSA) clinical isolates. PLoS ONE 2013, 8, e67398. [Google Scholar] [CrossRef]
- Bertsche, U.; Weidenmaier, C.; Kuehner, D.; Yang, S.-J.; Baur, S.; Wanner, S.; Francois, P.; Schrenzel, J.; Yeaman, M.R.; Bayer, A.S. Correlation of daptomycin resistance in a clinical Staphylococcus aureus strain with increased cell wall teichoic acid production and D-alanylation. Antimicrob. Agents Chemother. 2011, 55, 3922–3928. [Google Scholar] [CrossRef]
- Bayer, A.S.; Mishra, N.N.; Cheung, A.L.; Rubio, A.; Yang, S.J. Dysregulation of mprF and dltABCD expression among daptomycin-nonsusceptible MRSA clinical isolates. J. Antimicrob. Chemother. 2016, 71, 2100–2104. [Google Scholar] [CrossRef]
- Ma, Z.; Lasek-Nesselquist, E.; Lu, J.; Schneider, R.; Shah, R.; Oliva, G.; Pata, J.; McDonough, K.; Pai, M.P.; Rose, W.E.; et al. Characterization of genetic changes associated with daptomycin nonsusceptibility in Staphylococcus aureus. PLoS ONE 2018, 13, e0198366. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yao, H.; Li, D.; Liu, Z. The phosphatidylglycerol phosphate synthase PgsA utilizes a trifurcated amphipathic cavity for catalysis at the membrane-cytosol interface. Curr. Res. Struct. Biol. 2021, 3, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Róg, T.; Gurtovenko, A.A.; Vattulainen, I.; Karttunen, M. Role of Phosphatidylglycerols in the Stability of Bacterial Membranes. Biochimie 2008, 90, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.H.; Bhuiyan, M.S.; Shen, H.H.; Cameron, D.R.; Rupasinghe, T.W.T.; Wu, C.M.; Le Brun, A.P.; Kostoulias, X.; Domene, C.; Fulcher, A.J. Antibiotic resistance and host immune evasion in Staphylococcus aureus mediated by a metabolic adaptation. Proc. Natl. Acad. Sci. USA 2019, 116, 3722–3727. [Google Scholar] [CrossRef]
- Jones, T.; Yeaman, M.R.; Sakoulas, G.; Yang, S.J.; Proctor, R.A.; Sahl, H.G.; Schrenzel, J.; Xiong, Y.Q.; Bayer, A.S. Failures in clinical treatment of Staphylococcus aureus infection with daptomycin are associated with alterations in surface charge, membrane phospholipid asymmetry, and drug binding. Antimicrob. Agents Chemother. 2008, 52, 269–278. [Google Scholar] [CrossRef]
- Nikolic, P.; Mudgil, P. The Cell Wall, Cell Membrane and Virulence Factors of Staphylococcus aureus and Their Role in Antibiotic Resistance. Microorganisms 2023, 11, 259. [Google Scholar] [CrossRef]
- Hu, Q.; Peng, H.; Rao, X. Molecular events for promotion of vancomycin resistance in vancomycin intermediate Staphylococcus aureus. Front. Microbiol. 2016, 7, 1601. [Google Scholar] [CrossRef]
- Bhowmik, D.; Das, B.J.; Hazarika, M.; Chanda, D.D.; Bhattacharjee, A. Transcriptional analysis of prsA and vraTS regulatory system in methicillin resistant Staphylococcus aureus against oxacillin stress. Indian J. Med. Microbiol. 2022, 40, 57–60. [Google Scholar] [CrossRef]
- Gardete, S.; Kim, C.; Hartmann, B.M.; Mwangi, M.; Roux, C.M.; Dunman, P.M.; Chambers, H.F.; Tomasz, A. Genetic pathway in acquisition and loss of vancomycin resistance in a methicillin resistant Staphylococcus aureus (MRSA) strain of clonal type USA300. PLoS Pathog. 2012, 8, e1002505. [Google Scholar] [CrossRef]
- Olivares, A.O.; Baker, T.A.; Sauer, R.T. Mechanistic insights into bacterial AAA+ proteases and protein-remodelling machines. Nat. Rev. Microbiol. 2016, 14, 33–44. [Google Scholar] [CrossRef]
- Claunch, K.M.; Bush, M.; Evans, C.R.; Malmquist, J.A.; Hale, M.C.; McGillivray, S.M. Transcriptional profiling of the clpX mutant in Bacillus anthracis reveals regulatory connection with the lrgAB operon. Microbiology 2018, 164, 659–669. [Google Scholar] [CrossRef]
- Jensen, C.; Bæk, K.T.; Gallay, C.; Thalsø-Madsen, I.; Xu, L.; Jousselin, A.; Torrubia, F.R.; Paulander, W.; Pereira, A.R.; Veening, J.-W.; et al. The ClpX chaperone controls autolytic splitting of Staphylococcus aureus daughter cells, but is bypassed by β-lactam antibiotics or inhibitors of WTA biosynthesis. PLoS Pathog. 2019, 15, e1008044. [Google Scholar] [CrossRef]
- Frees, D.; Chastanet, A.; Qazi, S.; Sørensen, K.; Hill, P.; Msadek, T.; Ingmer, H. Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus. Mol. Microbiol. 2004, 54, 1445–1462. [Google Scholar] [CrossRef]
- McGillivra, S.M.; Ebrahimi, C.M.; Fisher, N.; Sabet, M.; Zhang, D.X.; Chen, Y.; Haste, N.M.; Aroian, R.V.; Gallo, R.L.; Guiney, D.G.; et al. ClpX contributes to innate defense peptide resistance and virulence phenotypes of Bacillus anthracis. J. Innate Immun. 2009, 1, 494–506. [Google Scholar] [CrossRef]
- McGillivray, S.M.; Tran, D.N.; Ramadoss, N.S.; Alumasa, J.N.; Okumura, C.Y.; Sakoulas, G.; Vaughn, M.M.; Zhang, D.X.; Keiler, K.C.; Nizet, V. Pharmacological inhibition of the ClpXP protease increases bacterial susceptibility to host cathelicidin antimicrobial peptides and cell envelope-active antibiotics. Antimicrob. Agents Chemother. 2012, 56, 1854–1861. [Google Scholar] [CrossRef]
- Zou, L.; Evans, C.R.; Do, V.D.; Losefsky, Q.P.; Ngo, D.Q.; McGillivray, S.M. Loss of the ClpXP Protease Leads to Decreased Resistance to Cell-Envelope Targeting Antimicrobials in Bacillus anthracis Sterne. Front. Microbiol. 2021, 12, 719548. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S. aureus Strains | Spa Type | MLST | MIC µg/mL (Broth Microdilution Method) of Selected Antibiotics | |
|---|---|---|---|---|
| Daptomycin | Gausemycin A | |||
| GAU-R | t3297 | ST464 | 5.00 | >200 |
| GAU-S | t3297 | ST464 | 1.25 | 2.5 |
| Gene | Mid-Exponential Phase | p-Value | Late-Exponential Phase | p-Value | Stationary Phase | p-Value |
|---|---|---|---|---|---|---|
| vraS | 4.53 up | 0.003508 | 6.19 up | 0.000015 | 1.48 down | 0.004713 |
| mprF | 1.56 up | 0.007591 | 1.30 up | 0.060267 | 5.5 down | 0.015934 |
| sceD | 1.54 down | 0.331873 | 3.79 up | 0.002776 | 1.47 up | 0.004713 |
| dltA | 8.72 up | 0.000002 | 4.58 up | 0.000109 | 0.71 down | 0.001534 |
| walK | 2.35 up | 0.000133 | 1.93 up | 0.0011 | 1.12 up | 0.190768 |
| pgsA | 3.39 up | 0.000553 | 8.32 up | 0.007331 | 0.36 down | 0.002019 |
| clpX | 6.24 up | 0.000112 | 10.25 up | 0.000085 | 6.78 down | 0.003211 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poshvina, D.V.; Dilbaryan, D.S.; Vasilchenko, A.S. Gausemycin A-Resistant Staphylococcus aureus Demonstrates Affected Cell Membrane and Cell Wall Homeostasis. Microorganisms 2023, 11, 1330. https://doi.org/10.3390/microorganisms11051330
Poshvina DV, Dilbaryan DS, Vasilchenko AS. Gausemycin A-Resistant Staphylococcus aureus Demonstrates Affected Cell Membrane and Cell Wall Homeostasis. Microorganisms. 2023; 11(5):1330. https://doi.org/10.3390/microorganisms11051330
Chicago/Turabian StylePoshvina, Darya V., Diana S. Dilbaryan, and Alexey S. Vasilchenko. 2023. "Gausemycin A-Resistant Staphylococcus aureus Demonstrates Affected Cell Membrane and Cell Wall Homeostasis" Microorganisms 11, no. 5: 1330. https://doi.org/10.3390/microorganisms11051330