
Klebsiella aerogenes Adhesion Behaviour during Biofilm Formation on Monazite

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Minerals
2.2. Micro-Organism and Culture Conditions
2.3. Cell Enumeration with Flow Cytometry
2.4. Bacterial and eDNA Attachment Behaviour
2.4.1. Effects of the Inoculum Size and Available Surface Area for Attachment on Bacterial Adsorption Behaviour
2.4.2. Effects of the Surface Chemical Properties on Bacterial Adsorption Behaviour
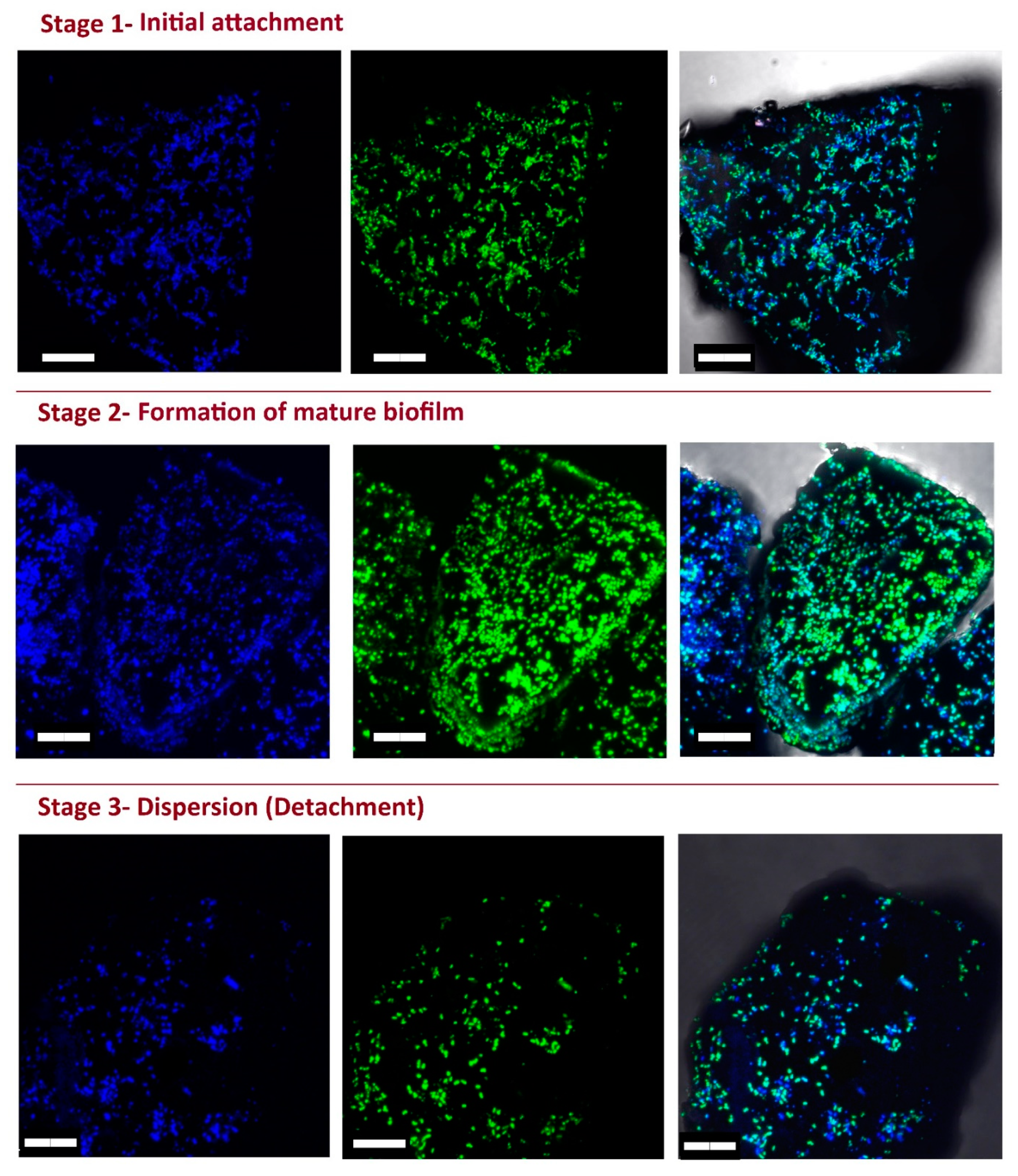
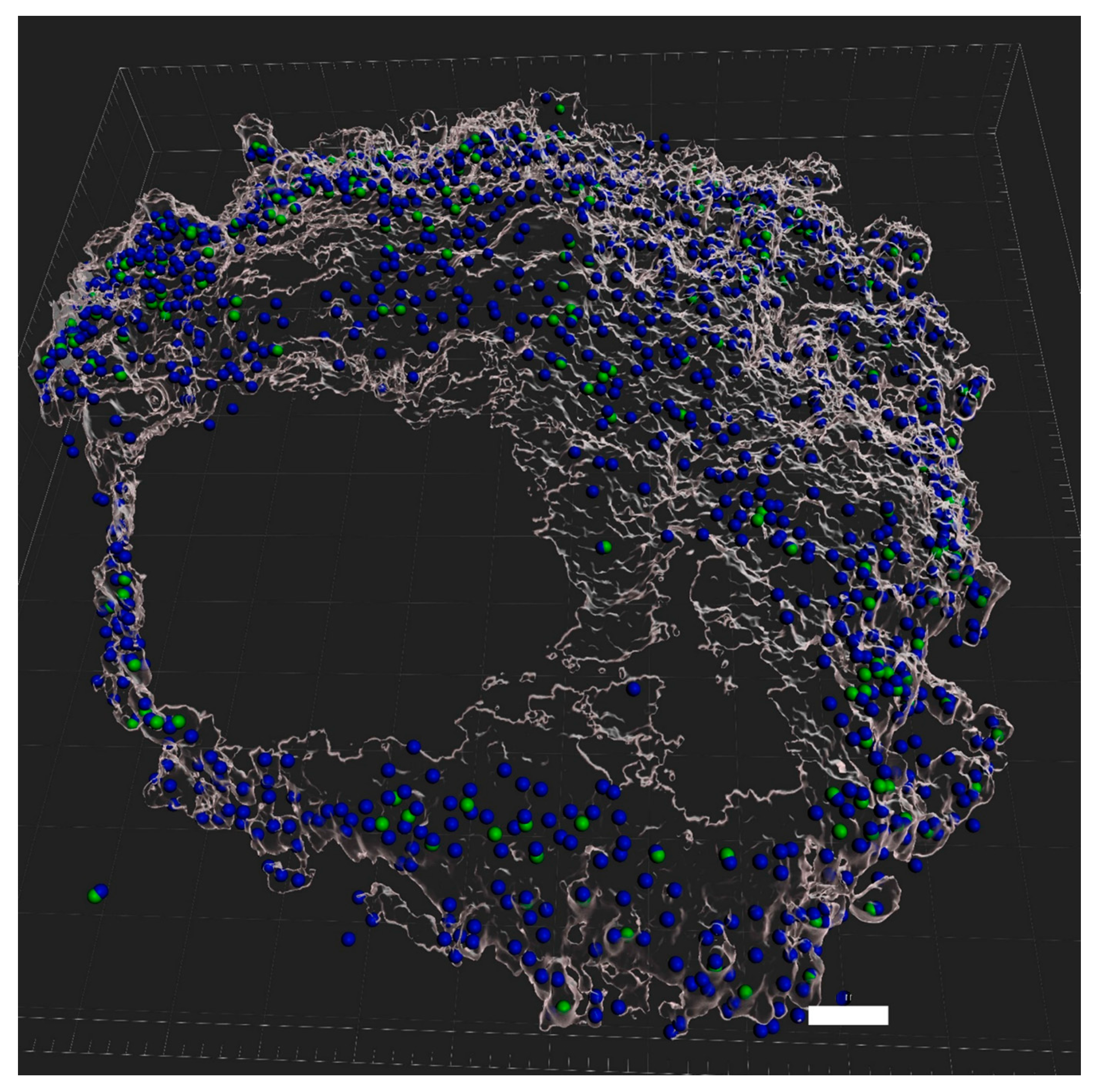
2.5. Live Imaging and Evaluation of eDNA Production
2.6. Fluorescent Microscopy
2.7. Measurement of Surface Electrical Potential
2.8. Statistics
3. Results
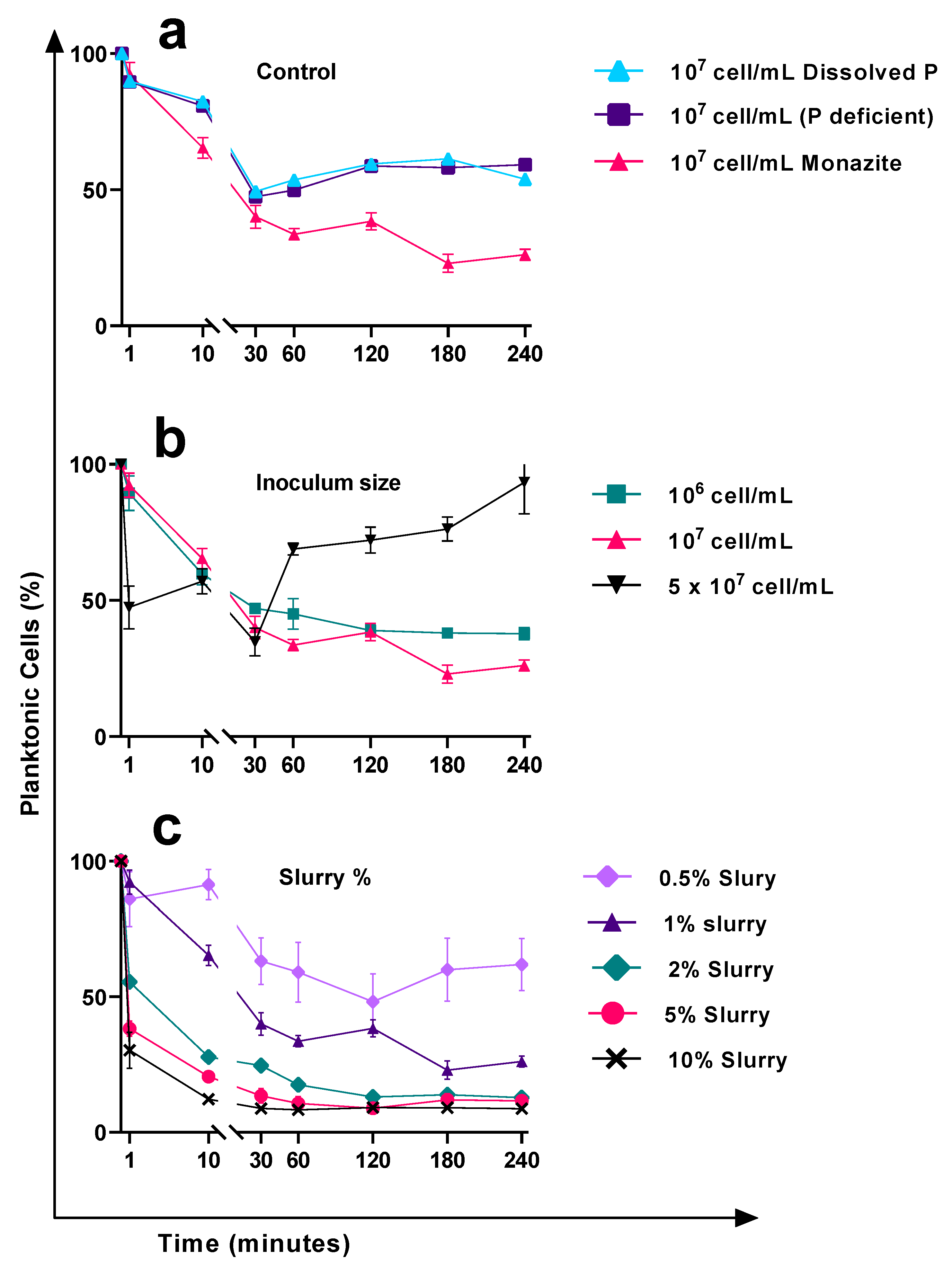
3.1. Attachment Behaviour in Response to the Monazite Particle Size
3.2. Cell Attachment Behaviour in Response to the Nutrient Amendment and Surface Material Available for Attachment
3.3. Cell Attachment Behaviour in Response to the Changes in the Initial Inoculum Size
3.4. The Effects of the Available Area for Attachment on the Attachment Efficiency
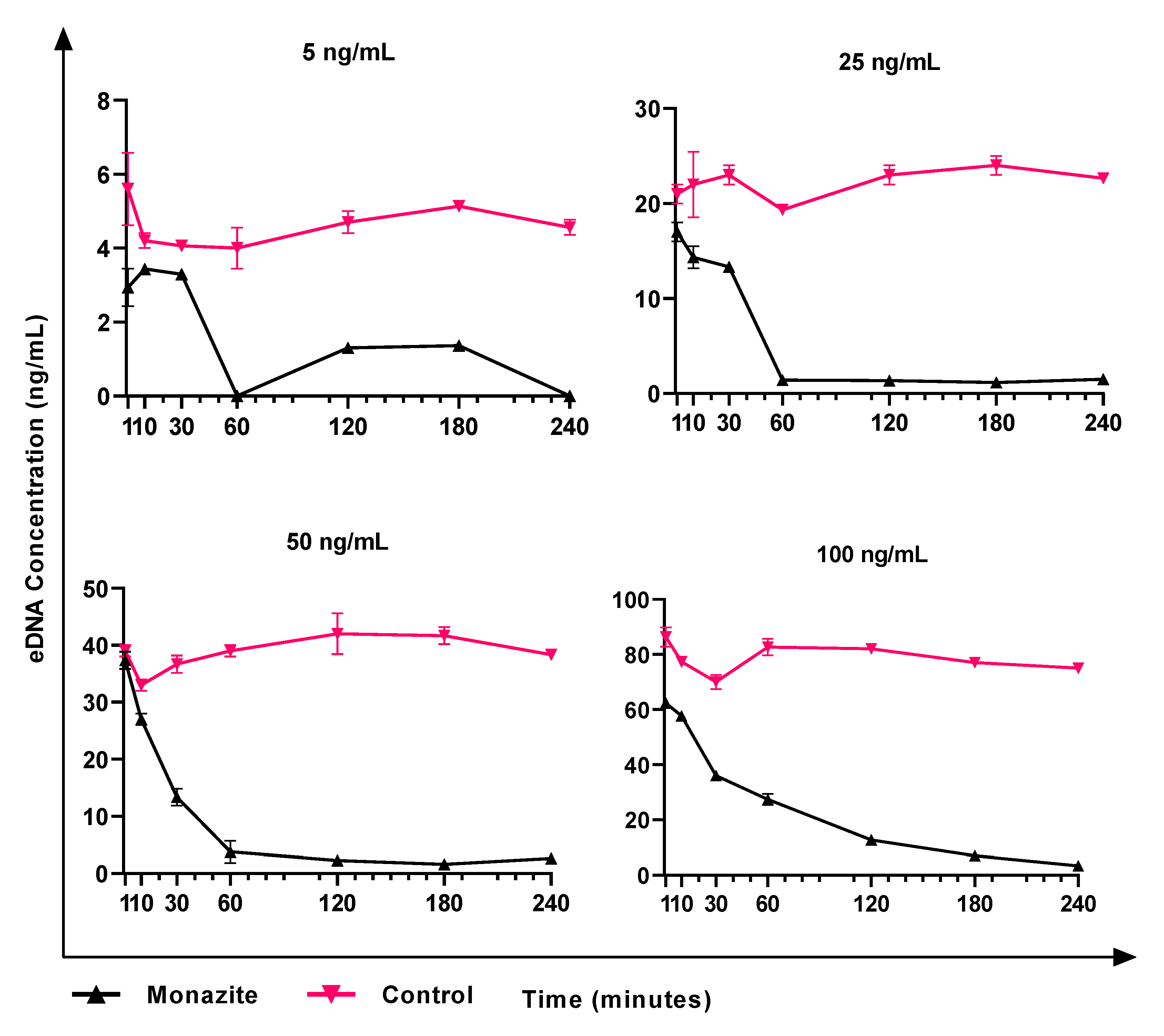
3.5. eDNA Production by K. aerogenes
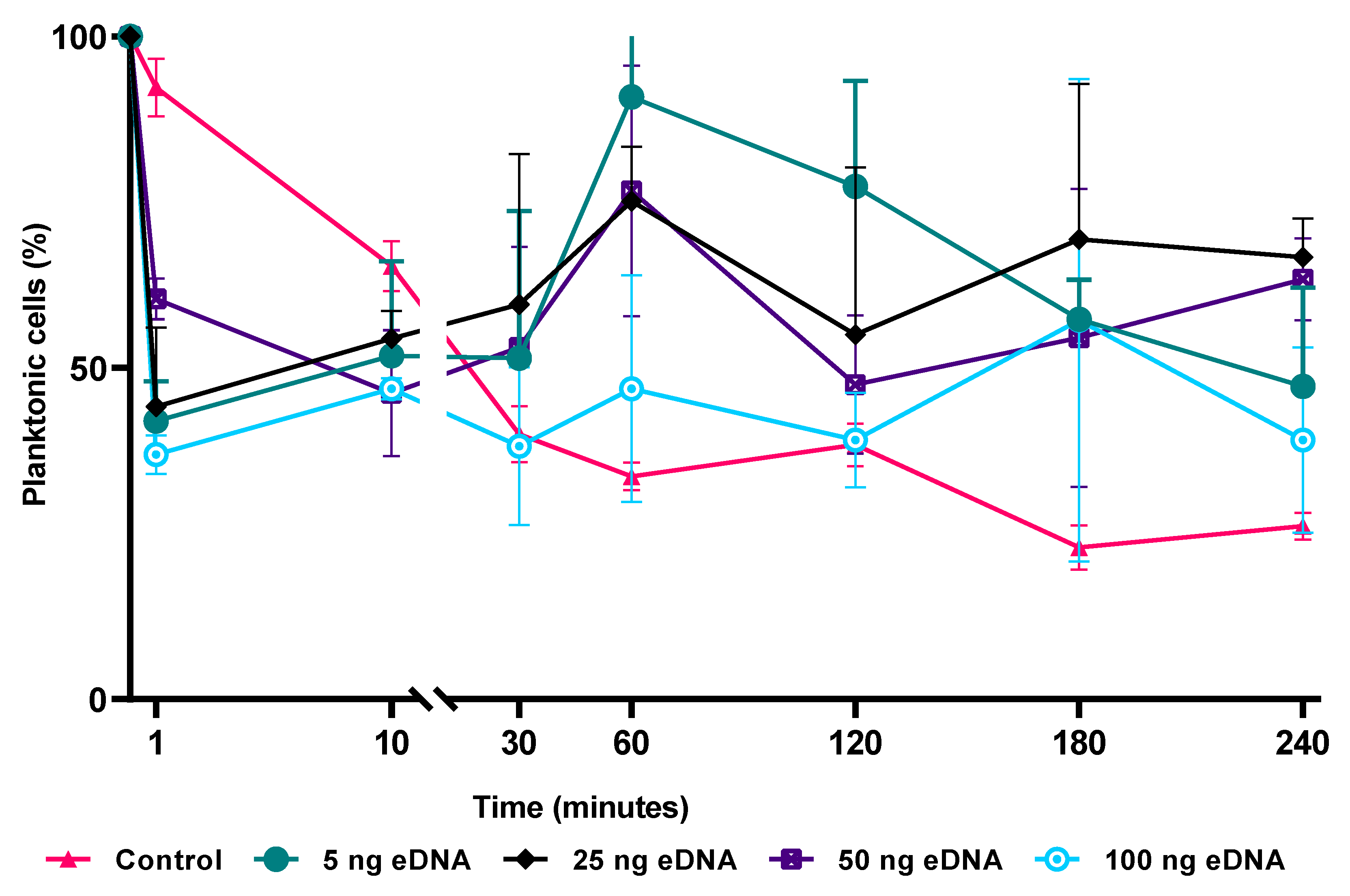
3.6. Role of eDNA in Early Attachment and Interaction with the Surface
3.7. Effects of the Surface Preconditioning with eDNA on Attachment of K. aerogenes Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: Expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Berne, C.; Ellison, C.K.; Ducret, A.; Brun, Y.V. Bacterial adhesion at the single-cell level. Nat. Rev. Microbiol. 2018, 16, 616–627. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Römling, U.; Kjelleberg, S.; Normark, S.; Nyman, L.; Uhlin, B.E.; Åkerlund, B. Microbial biofilm formation: A need to act. J. Intern. Med. 2014, 276, 98–110. [Google Scholar] [CrossRef]
- Li, Q.; Sand, W.; Zhang, R. Enhancement of biofilm formation on pyrite by Sulfobacillus thermosulfidooxidans. Minerals 2016, 6, 71. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Wille, J.; Coenye, T. Biofilm dispersion: The key to biofilm eradication or opening Pandora’s box? Biofilm 2020, 2, 100027. [Google Scholar] [CrossRef]
- Alhede, M.; Kragh, K.N.; Qvortrup, K.; Allesen-Holm, M.; van Gennip, M.; Christensen, L.D.; Jensen, P.Ø.; Nielsen, A.K.; Parsek, M.; Wozniak, D.; et al. Phenotypes of Non-Attached Pseudomonas aeruginosa Aggregates Resemble Surface Attached Biofilm. PLoS ONE 2011, 6, e27943. [Google Scholar] [CrossRef]
- Cai, Y.-M. Non-surface Attached Bacterial Aggregates: A Ubiquitous Third Lifestyle. Front. Microbiol. 2020, 11, 557035. [Google Scholar] [CrossRef]
- Moreno-Paz, M.; Gómez, M.J.; Arcas, A.; Parro, V. Environmental transcriptome analysis reveals physiological differences between biofilm and planktonic modes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural microbial community. BMC Genom. 2010, 11, 404. [Google Scholar] [CrossRef]
- Kaksonen, A.H.; Boxall, N.J.; Gumulya, Y.; Khaleque, H.N.; Morris, C.; Bohu, T.; Cheng, K.Y.; Usher, K.M.; Lakaniemi, A.M. Recent progress in biohydrometallurgy and microbial characterisation. Hydrometallurgy 2018, 180, 7–25. [Google Scholar] [CrossRef]
- Fathollahzadeh, H.; Becker, T.; Eksteen, J.J.; Kaksonen, A.H.; Watkin, E.L.J. Microbial contact enhances bioleaching of rare earth elements. Bioresour. Technol. Rep. 2018, 3, 102–108. [Google Scholar] [CrossRef]
- Corbett, M.K.; Eksteen, J.J.; Niu, X.-Z.; Watkin, E.L.J. Syntrophic effect of indigenous and inoculated microorganisms in the leaching of rare earth elements from Western Australian monazite. Res. Microbiol. 2018, 169, 558–568. [Google Scholar] [CrossRef]
- Alin, A.V.; Corbett, M.K.; Fathollahzadeh, H.; Tjiam, M.C.; Rickard, W.D.A.; Sun, X.; Putnis, A.; Eksteen, J.; Kaksonen, A.H.; Watkin, E. Biofilm formation on the surface of monazite and xenotime during bioleaching. 2023; in press. [Google Scholar]
- Flemming, H.-C.; van Hullebusch, E.D.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. The biofilm matrix: Multitasking in a shared space. Nat. Rev. Microbiol. 2022, 21, 70–86. [Google Scholar] [CrossRef]
- Allison, D.G. The Biofilm Matrix. Biofouling 2003, 19, 139–150. [Google Scholar] [CrossRef]
- Okshevsky, M.; Meyer, R.L. The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit. Rev. Microbiol. 2015, 41, 341–352. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Y.; Chu, H.; Tian, B.; Li, H.; Zhang, M.; Xin, B. Enhanced metal bioleaching mechanisms of extracellular polymeric substance for obsolete LiNixCoyMn1-x-yO2 at high pulp density. J. Environ. Manag. 2022, 318, 115429. [Google Scholar] [CrossRef]
- Zhang, R.; Bellenberg, S.; Neu, T.R.; Sand, W.; Vera, M. The Biofilm Lifestyle of Acidophilic Metal/Sulfur-Oxidizing Microorganisms. In Biotechnology of Extremophiles: Advances and Challenges; Rampelotto, P.H., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 177–213. [Google Scholar]
- Sand, W.; Gehrke, T. Extracellular polymeric substances mediate bioleaching/biocorrosion via interfacial processes involving iron(III) ions and acidophilic bacteria. Res. Microbiol. 2006, 157, 49–56. [Google Scholar] [CrossRef]
- Kinzler, K.; Gehrke, T.; Telegdi, J.; Sand, W. Bioleaching—A result of interfacial processes caused by extracellular polymeric substances (EPS). Hydrometallurgy 2003, 71, 83–88. [Google Scholar] [CrossRef]
- Rodriguez-Leiva, M.; Tributsch, H. Morphology of bacterial leaching patterns by Thiobacillus ferrooxidans on synthetic pyrite. Arch. Microbiol. 1988, 149, 401–405. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Secchi, E.; Savorana, G.; Vitale, A.; Eberl, L.; Stocker, R.; Rusconi, R. The structural role of bacterial eDNA in the formation of biofilm streamers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113723119. [Google Scholar] [CrossRef]
- Yu, R.; Hou, C.; Liu, A.; Peng, T.; Xia, M.; Wu, X.; Shen, L.; Liu, Y.; Li, J.; Yang, F.; et al. Extracellular DNA enhances the adsorption of Sulfobacillus thermosulfidooxidans strain ST on chalcopyrite surface. Hydrometallurgy 2018, 176, 97–103. [Google Scholar] [CrossRef]
- Tuck, B.; Watkin, E.; Somers, A.; Forsyth, M.; Machuca, L.L. Conditioning of metal surfaces enhances Shewanella chilikensis adhesion. Biofouling 2022, 38, 207–222. [Google Scholar] [CrossRef]
- Campoccia, D.; Montanaro, L.; Arciola, C.R. Extracellular DNA (eDNA). A Major Ubiquitous Element of the Bacterial Biofilm Architecture. Int. J. Mol. Sci. 2021, 22, 9100. [Google Scholar] [CrossRef]
- Berne, C.; Kysela, D.T.; Brun, Y.V. A bacterial extracellular DNA inhibits settling of motile progeny cells within a biofilm. Mol. Microbiol. 2010, 77, 815–829. [Google Scholar] [CrossRef]
- Fathollahzadeh, H.; Hackett, M.J.; Khaleque, H.N.; Eksteen, J.J.; Kaksonen, A.H.; Watkin, E.L.J. Better together: Potential of co-culture microorganisms to enhance bioleaching of rare earth elements from monazite. Bioresour. Technol. Rep. 2018, 3, 109–118. [Google Scholar] [CrossRef]
- Corbett, M.K.; Eksteen, J.J.; Niu, X.-Z.; Croue, J.-P.; Watkin, E.L.J.J.B.; Engineering, B. Interactions of phosphate solubilising microorganisms with natural rare-earth phosphate minerals: A study utilizing Western Australian monazite. Bioprocess Biosyst. Eng. 2017, 40, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Fathollahzadeh, H.; Eksteen, J.J.; Kaksonen, A.H.; Watkin, E.L.J. Role of microorganisms in bioleaching of rare earth elements from primary and secondary resources. Appl. Microbiol. Biotechnol. 2019, 103, 1043–1057. [Google Scholar] [CrossRef]
- Allard, S.; Gutierrez, L.; Fontaine, C.; Croué, J.-P.; Gallard, H. Organic matter interactions with natural manganese oxide and synthetic birnessite. Sci. Total Environ. 2017, 583, 487–495. [Google Scholar] [CrossRef]
- Rafael, R.B.A.; FernÁNdez-Marcos, M.L.; Cocco, S.; Ruello, M.L.; Weindorf, D.C.; Cardelli, V.; Corti, G. Assessment of Potential Nutrient Release from Phosphate Rock and Dolostone for Application in Acid Soils. Pedosphere 2018, 28, 44–58. [Google Scholar] [CrossRef]
- Feng, S.; Yang, H.; Wang, W. Insights to the effects of free cells on community structure of attached cells and chalcopyrite bioleaching during different stages. Bioresour. Technol. 2016, 200, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Alin, A.; Corbett, M.K.; Fathollahzadeh, H.; Tjiam, M.C.; Putnis, A.; Eksteen, J.; Kaksonen, A.H.; Watkin, E. Klebsiella aerogenes Adhesion Behaviour during Biofilm Formation on Monazite. Microorganisms 2023, 11, 1331. https://doi.org/10.3390/microorganisms11051331
Van Alin A, Corbett MK, Fathollahzadeh H, Tjiam MC, Putnis A, Eksteen J, Kaksonen AH, Watkin E. Klebsiella aerogenes Adhesion Behaviour during Biofilm Formation on Monazite. Microorganisms. 2023; 11(5):1331. https://doi.org/10.3390/microorganisms11051331
Chicago/Turabian StyleVan Alin, Arya, Melissa K. Corbett, Homayoun Fathollahzadeh, M. Christian Tjiam, Andrew Putnis, Jacques Eksteen, Anna H. Kaksonen, and Elizabeth Watkin. 2023. "Klebsiella aerogenes Adhesion Behaviour during Biofilm Formation on Monazite" Microorganisms 11, no. 5: 1331. https://doi.org/10.3390/microorganisms11051331