
Phenotypic Variation in Clinical S. aureus Isolates Did Not Affect Disinfection Efficacy Using Short-Term UV-C Radiation
 , , ,
, , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Quantification of Staphyloxanthin Expression
2.2. Measuring Catalase Activity
2.3. Quantification of Biofilm Formation
2.4. UV-C Disinfection Procedure
2.5. Testing UV-C Tolerance Using Reality-Simulating Experimental Conditions
3. Results
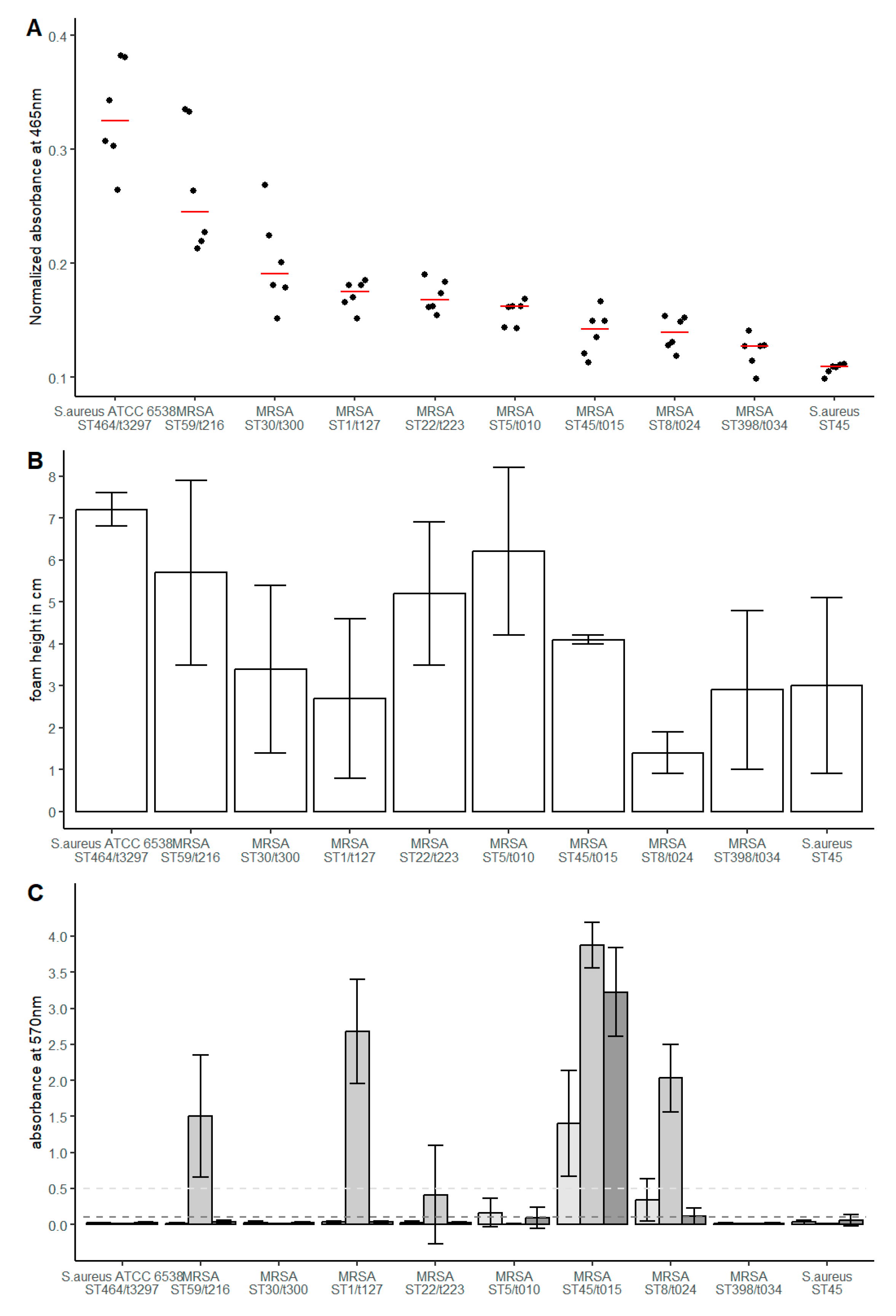
3.1. Genotypic and Phenotypic Characteristics of the Investigated Isolates
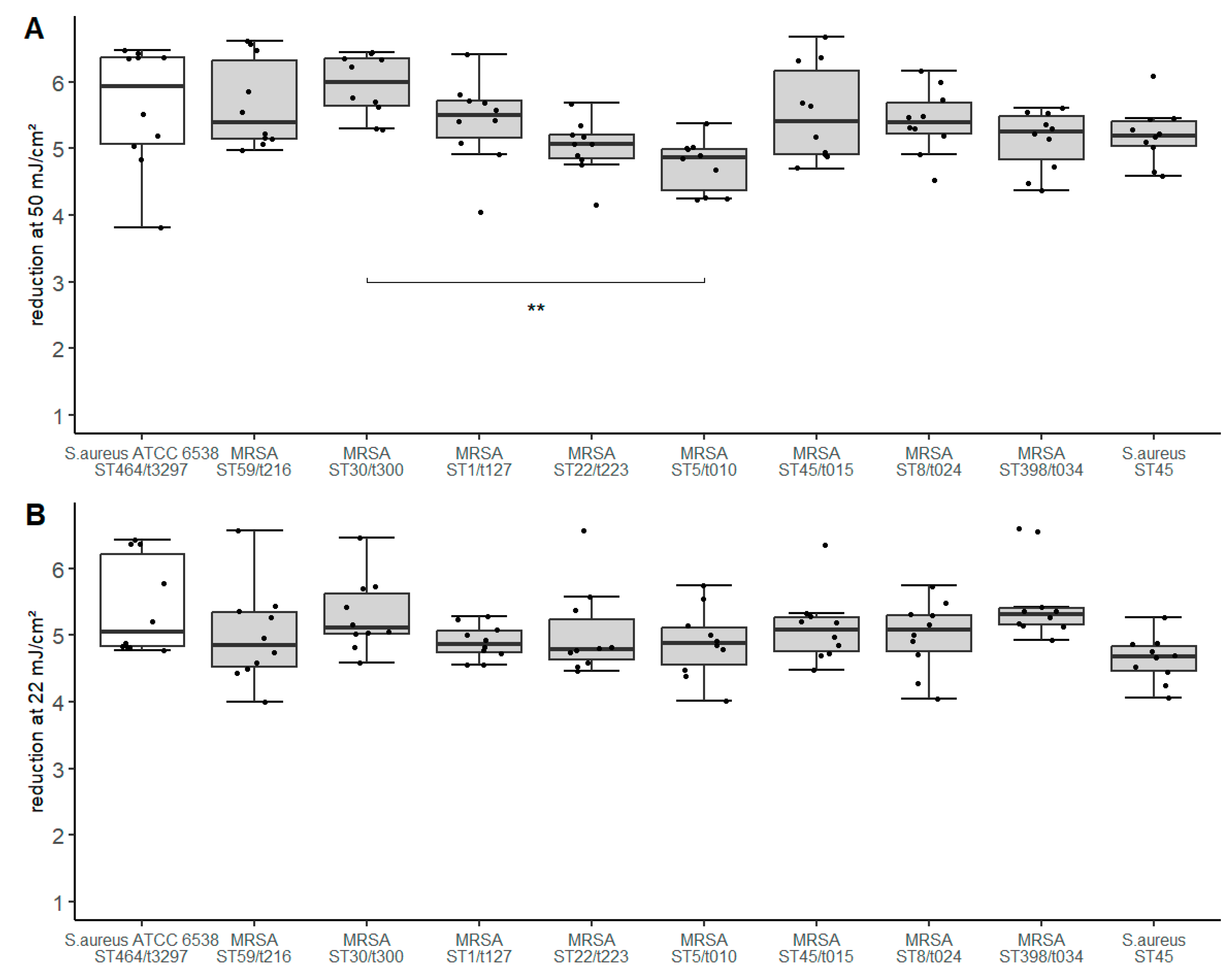
3.2. UV-C Disinfection of Investigated Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Algammal, A.M.; Abo Hashem, M.E.; Alfifi, K.J.; Al-Otaibi, A.S.; Alatawy, M.; ElTarabili, R.M.; Abd El-Ghany, W.A.; Hetta, H.F.; Hamouda, A.M.; Elewa, A.A.; et al. Sequence Analysis, Antibiogram Profile, Virulence and Antibiotic Resistance Genes of XDR and MDR Gallibacterium anatis Isolated from Layer Chickens in Egypt. Infect. Drug Resist. 2022, 15, 4321–4334. [Google Scholar] [CrossRef] [PubMed]
- Algammal, A.M.; Enany, M.E.; El-Tarabili, R.M.; Ghobashy, M.O.I.; Helmy, Y.A. Prevalence, Antimicrobial Resistance Profiles, Virulence and Enterotoxins-Determinant Genes of MRSA Isolated from Subclinical Bovine Mastitis in Egypt. Pathogens 2020, 9, 362. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.J.; Pfaller, M.A.; Shortridge, D.; Zervos, M.; Jones, R.N. Twenty-Year Trends in Antimicrobial Susceptibilities Among Staphylococcus aureus From the SENTRY Antimicrobial Surveillance Program. Open Forum Infect. Dis. 2019, 6, S47–S53. [Google Scholar] [CrossRef] [PubMed]
- Bal, A.M.; Coombs, G.W.; Holden, M.T.G.; Lindsay, J.A.; Nimmo, G.R.; Tattevin, P.; Skov, R.L. Genomic insights into the emergence and spread of international clones of healthcare-, community- and livestock-associated meticillin-resistant Staphylococcus aureus: Blurring of the traditional definitions. J. Glob. Antimicrob. Resist. 2016, 6, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Wißmann, J.E.; Kirchhoff, L.; Brüggemann, Y.; Todt, D.; Steinmann, J.; Steinmann, E. Persistence of Pathogens on Inanimate Surfaces: A Narrative Review. Microorganisms 2021, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Wolfensberger, A.; Mang, N.; Gibson, K.E.; Gontjes, K.; Cassone, M.; Brugger, S.D.; Mody, L.; Sax, H. Understanding short-term transmission dynamics of methicillin-resistant Staphylococcus aureus in the patient room. Infect. Control Hosp. Epidemiol. 2022, 43, 1147–1154. [Google Scholar] [CrossRef]
- Tanner, W.D.; Leecaster, M.K.; Zhang, Y.; Stratford, K.M.; Mayer, J.; Visnovsky, L.D.; Alhmidi, H.; Cadnum, J.L.; Jencson, A.L.; Koganti, S.; et al. Environmental Contamination of Contact Precaution and Non-Contact Precaution Patient Rooms in Six Acute Care Facilities. Clin. Infect. Dis. 2021, 72, S8–S16. [Google Scholar] [CrossRef]
- Meyer, J.; Nippak, P.; Cumming, A. An evaluation of cleaning practices at a teaching hospital. Am. J. Infect. Control 2021, 49, 40–43. [Google Scholar] [CrossRef]
- Chen, L.F.; Knelson, L.P.; Gergen, M.F.; Better, O.M.; Nicholson, B.P.; Woods, C.W.; Rutala, W.A.; Weber, D.J.; Sexton, D.J.; Anderson, D.J. A prospective study of transmission of Multidrug-Resistant Organisms (MDROs) between environmental sites and hospitalized patients-the TransFER study. Infect. Control Hosp. Epidemiol. 2019, 40, 47–52. [Google Scholar] [CrossRef]
- Wu, Y.-L.; Yang, X.-Y.; Ding, X.-X.; Li, R.-J.; Pan, M.-S.; Zhao, X.; Hu, X.-Q.; Zhang, J.-J.; Yang, L.-Q. Exposure to infected/colonized roommates and prior room occupants increases the risks of healthcare-associated infections with the same organism. J. Hosp. Infect. 2019, 101, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Cohen, C.C.; Løyland, B.; Larson, E.L. Transmission of health care-associated infections from roommates and prior room occupants: A systematic review. Clin. Epidemiol. 2017, 9, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Masjoudi, M.; Mohseni, M.; Bolton, J.R. Sensitivity of Bacteria, Protozoa, Viruses, and Other Microorganisms to Ultraviolet Radiation. J. Res. Natl. Inst. Stan. 2021, 126, 1–77. [Google Scholar] [CrossRef]
- Mariita, R.M.; Randive, R.V. Disinfection of methicillin-resistant Staphylococcus aureus, vancomycin-resistant Enterococcus faecium and Acinetobacter baumannii using Klaran WD array system. Access Microbiol. 2021, 3, 194. [Google Scholar] [CrossRef]
- Christenson, E.C.; Cronk, R.; Atkinson, H.; Bhatt, A.; Berdiel, E.; Cawley, M.; Cho, G.; Coleman, C.K.; Harrington, C.; Heilferty, K.; et al. Evidence Map and Systematic Review of Disinfection Efficacy on Environmental Surfaces in Healthcare Facilities. Int. J. Environ. Res. Public Health 2021, 18, 11100. [Google Scholar] [CrossRef]
- EN 14885:2018; Chemical Disinfectants and Antiseptics—Application of European Standards for Chemical Disinfectants and Antiseptics. Deutsches Insitut für Normung e.V.: Berlin, Germany; Beuth Verlag GmbH: Berlin, Germany, 2018. (In German)
- Zwicker, P.; Schleusener, J.; Lohan, S.B.; Busch, L.; Sicher, C.; Einfeldt, S.; Kneissl, M.; Kühl, A.A.; Keck, C.M.; Witzel, C.; et al. Application of 233 nm far-UVC LEDs for eradication of MRSA and MSSA and risk assessment on skin models. Sci. Rep. 2022, 12, 2587. [Google Scholar] [CrossRef]
- Pannu, M.K.; Hudman, D.A.; Sargentini, N.J.; Singh, V.K. Role of SigB and Staphyloxanthin in Radiation Survival of Staphylococcus aureus. Curr. Microbiol. 2019, 76, 70–77. [Google Scholar] [CrossRef]
- Sullivan, L.E.; Rice, K.C. Measurement of Staphylococcus aureus Pigmentation by Methanol Extraction. Methods Mol. Biol. 2021, 2341, 1–7. [Google Scholar] [CrossRef]
- Iwase, T.; Tajima, A.; Sugimoto, S.; Okuda, K.; Hironaka, I.; Kamata, Y.; Takada, K.; Mizunoe, Y. A simple assay for measuring catalase activity: A visual approach. Sci. Rep. 2013, 3, 3081. [Google Scholar] [CrossRef]
- Pottage, T.; Macken, S.; Walker, J.T.; Bennett, A.M. Meticillin-resistant Staphylococcus aureus is more resistant to vaporized hydrogen peroxide than commercial Geobacillus stearothermophilus biological indicators. J. Hosp. Infect. 2012, 80, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Bono, N.; Ponti, F.; Punta, C.; Candiani, G. Effect of UV Irradiation and TiO2-Photocatalysis on Airborne Bacteria and Viruses: An Overview. Materials 2021, 14, 1075. [Google Scholar] [CrossRef]
- Chaggar, G.K.; Nkemngong, C.A.; Li, X.; Teska, P.J.; Oliver, H.F. Hydrogen peroxide, sodium dichloro-s-triazinetriones and quaternary alcohols significantly inactivate the dry-surface biofilms of Staphylococcus aureus and Pseudomonas aeruginosa more than quaternary ammoniums. Microbiology 2022, 168, 001140. [Google Scholar] [CrossRef]
- Otter, J.A.; Vickery, K.; Walker, J.T.; deLancey Pulcini, E.; Stoodley, P.; Goldenberg, S.D.; Salkeld, J.A.G.; Chewins, J.; Yezli, S.; Edgeworth, J.D. Surface-attached cells, biofilms and biocide susceptibility: Implications for hospital cleaning and disinfection. J. Hosp. Infect. 2015, 89, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Elasri, M.O.; Miller, R.V. Study of the response of a biofilm bacterial community to UV radiation. Appl. Environ. Microbiol. 1999, 65, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Ledwoch, K.; Dancer, S.J.; Otter, J.A.; Kerr, K.; Roposte, D.; Rushton, L.; Weiser, R.; Mahenthiralingam, E.; Muir, D.D.; Maillard, J.-Y. Beware biofilm! Dry biofilms containing bacterial pathogens on multiple healthcare surfaces; a multi-centre study. J. Hosp. Infect. 2018, 100, e47–e56. [Google Scholar] [CrossRef]
- Brooks, J.L.; Jefferson, K.K. Phase variation of poly-N-acetylglucosamine expression in Staphylococcus aureus. PLoS Pathog. 2014, 10, e1004292. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026-19. [Google Scholar] [CrossRef]
- Knobloch, J.K.-M.; Horstkotte, M.A.; Rohde, H.; Mack, D. Evaluation of different detection methods of biofilm formation in Staphylococcus aureus. Med. Microbiol. Immunol. 2002, 191, 101–106. [Google Scholar] [CrossRef]
- Scott, R.; Joshi, L.T.; McGinn, C. Hospital surface disinfection using ultraviolet germicidal irradiation technology: A review. Healthc. Technol. Lett. 2022, 9, 25–33. [Google Scholar] [CrossRef]
- Beswick, A.J.; Fry, C.; Bradley, C.R.; Pottage, T.; Sharpe, S.; Haill, C.F.; Mugglestone, M.A.; Bak, A.; Marsden, G.L.; Bennett, A.; et al. Automated room decontamination: Report of a Healthcare Infection Society Working Party. J. Hosp. Infect. 2022, 124, 97–120. [Google Scholar] [CrossRef]
- Carlsen, L.; Büttner, H.; Christner, M.; Franke, G.; Indenbirken, D.; Knobling, B.; Lütgehetmann, M.; Knobloch, J. High burden and diversity of carbapenemase-producing Enterobacterales observed in wastewater of a tertiary care hospital in Germany. Int. J. Hyg. Environ. Health 2022, 242, 113968. [Google Scholar] [CrossRef] [PubMed]
- Aires-de-Sousa, M. Methicillin-resistant Staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2017, 23, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Bartscht, K.; Fischer, C.; Rohde, H.; de Grahl, C.; Dobinsky, S.; Horstkotte, M.A.; Kiel, K.; Knobloch, J.K. Genetic and biochemical analysis of Staphylococcus epidermidis biofilm accumulation. Methods Enzymol. 2001, 336, 215–239. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, J.K.; Bartscht, K.; Sabottke, A.; Rohde, H.; Feucht, H.H.; Mack, D. Biofilm formation by Staphylococcus epidermidis depends on functional RsbU, an activator of the sigB operon: Differential activation mechanisms due to ethanol and salt stress. J. Bacteriol. 2001, 183, 2624–2633. [Google Scholar] [CrossRef]
- EN 17272:2020; Chemical Disinfectants and Antiseptics—Methods of Airborne Room Disinfection by Automated Process—Determination of Bactericidal, Mycobactericidal, Sporicidal, Fungicidal, Yeasticidal, Virucidal and Phagocidal Activities. Deutsches Insitut für Normung e.V.: Berlin, Germany; Beuth Verlag GmbH: Berlin, Germany, 2020. (In German)
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Posit Team. RStudio: Integrated Development Environment for R. Posit Software; Posit Software, PBC: Boston, MA, USA, 2022. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. 2020. Available online: https://cran.r-project.org/web/packages/ggpubr/index.html (accessed on 20 December 2022).
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2021. Available online: https://cloud.r-project.org/web/packages/rstatix/index.html (accessed on 20 December 2022).
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal response to oxidative stress. Front. Cell. Infect. Microbiol. 2012, 2, 33. [Google Scholar] [CrossRef]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef]
- Inose, Y.; Takeshita, S.L.; Hidaka, T.; Higashide, M.; Maruyama, A.; Hayashi, H.; Morikawa, K.; Ohta, T. Genetic characterization of the natural SigB variants found in clinical isolates of Staphylococcus aureus. J. Gen. Appl. Microbiol. 2006, 52, 259–271. [Google Scholar] [CrossRef]
- Painter, K.L.; Strange, E.; Parkhill, J.; Bamford, K.B.; Armstrong-James, D.; Edwards, A.M. Staphylococcus aureus adapts to oxidative stress by producing H2O2-resistant small-colony variants via the SOS response. Infect. Immun. 2015, 83, 1830–1844. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knobling, B.; Franke, G.; Carlsen, L.; Belmar Campos, C.; Büttner, H.; Klupp, E.M.; Maurer, P.M.; Knobloch, J.K. Phenotypic Variation in Clinical S. aureus Isolates Did Not Affect Disinfection Efficacy Using Short-Term UV-C Radiation. Microorganisms 2023, 11, 1332. https://doi.org/10.3390/microorganisms11051332
Knobling B, Franke G, Carlsen L, Belmar Campos C, Büttner H, Klupp EM, Maurer PM, Knobloch JK. Phenotypic Variation in Clinical S. aureus Isolates Did Not Affect Disinfection Efficacy Using Short-Term UV-C Radiation. Microorganisms. 2023; 11(5):1332. https://doi.org/10.3390/microorganisms11051332
Chicago/Turabian StyleKnobling, Birte, Gefion Franke, Laura Carlsen, Cristina Belmar Campos, Henning Büttner, Eva M. Klupp, Philipp Maximilian Maurer, and Johannes K. Knobloch. 2023. "Phenotypic Variation in Clinical S. aureus Isolates Did Not Affect Disinfection Efficacy Using Short-Term UV-C Radiation" Microorganisms 11, no. 5: 1332. https://doi.org/10.3390/microorganisms11051332