
Nitrogen Fixation Activity and Genome Analysis of a Moderately Haloalkaliphilic Anoxygenic Phototrophic Bacterium Rhodovulum tesquicola

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Cultivation
2.2. Nitrogenase Activity Measurement
2.3. Genome Sequencing and Assembly
3. Results and Discussion
3.1. General Characteristics of Genome
3.2. Carbon Metabolism
3.3. Nitrogen Metabolism
3.4. Vitamin Requirements
3.5. Osmotic Adaptation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imhoff, J.F. Genus XIV Rhodovulum. In Bergey’s Manual of Systematic Bacteriology, Volume 2: The Proteobacteria, Part C: The Alpha-, Beta-, Delta-, and Epsilonproteobacteria; Garrity, G., Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Hiraishi, A.; Ueda, Y. Intrageneric structure of the genus Rhodobacter: Transfer of Rhodobacter sulfidophilus and related marine species to the genus Rhodovulum gen. nov. Int. J. Syst. Evol. Microbiol. 1994, 44, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Hiraishi, A.; Nagao, N.; Yonekawa, C.; Umekage, S.; Kikuchi, Y.; Eki, T.; Hirose, Y. Distribution of phototrophic purple nonsulfur bacteria in massive blooms in coastal and wastewater ditch environments. Microorganisms 2020, 8, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff, J.F. True marine and halophilic anoxygenic phototrophic bacteria. Arch. Microbiol. 2001, 176, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Kompantseva, E.I.; Komova, A.V.; Krauzova, V.I.; Kolganova, T.V.; Panteleeva, A.N. Purple nonsulfur bacteria in weakly and moderately mineralized soda lakes of the southern Transbaikal Region and northeastern Mongolia. Mikrobiologia 2009, 78, 246–253. [Google Scholar] [CrossRef]
- Fujimoto, H.; Wakabayashi, M.; Yamashiro, H.; Maeda, I.; Isoda, K.; Kondoh, M.; Kawase, M.; Miyasaka, H.; Yagi, K. Whole-cell arsenite biosensor using photosynthetic bacterium Rhodovulum sulfidophilum. Appl. Microbiol. Biot. 2006, 73, 332–338. [Google Scholar] [CrossRef]
- Cai, J.; Wang, G. Hydrogen production by a marine photosynthetic bacterium, Rhodovulum sulfidophilum P5, isolated from a shrimp pond. Int. J. Hydrogen. Energy 2012, 37, 15070–15080. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Morisaki, K.; Toyooka, K.; Numata, K. Synthesis of high-molecular-weight polyhydroxyalkanoates by marine photosynthetic purple bacteria. PLoS ONE 2016, 11, e0160981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Guzman, M.S.; Rengasamy, K.; Stoica, A.; Singh, R.; Ranaivoarisoa, T.O.; Davenport, E.J.; Bai, W.; McGinley, B.; Meacham, J.M.; et al. Photoferrotrophy and phototrophic extracellular electron uptake is common in the marine anoxygenic phototroph Rhodovulum sulfidophilum. ISME 2021, 15, 3384–3398. [Google Scholar] [CrossRef] [PubMed]
- Melack, J.M.; Kilham, P. Photosynthetic rates of phytoplankton in East African alkaline, saline lakes 1. Limnol. Oceanogr. 1974, 19, 743–755. [Google Scholar] [CrossRef]
- Cloern, J.E.; Cole, B.E.; Oremland, R.S. Autotrophic processes in meromictic Big Soda Lake, Nevada. Limnol. Oceanogr. 1983, 28, 1049–1061. [Google Scholar] [CrossRef]
- Jellison, R.; Miller, L.G.; Melack, J.M.; Dana, G.L. Meromixis in hypersaline Mono Lake, California. 2. Nitrogen fluxes. Limnol. Oceanogr. 1993, 38, 1020–1039. [Google Scholar] [CrossRef]
- Oremland, R.S. Nitrogen fixation dynamics of two diazotrophic communities in Mono Lake, California. Appl. Environ. Microbiol. 1990, 56, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Dubinin, A.V.; Gerasimenko, L.M.; Zavarzin, G.A. Nitrogen-fixation by cyanobacterium Microcoleus chthonoplastes from hypersaline lagoons od Lake Sivash. Mikrobiologia 1992, 61, 593–597. [Google Scholar]
- Pinckney, J.L.; Paerl, H.W. Anoxygenic photosynthesis and nitrogen fixation by a microbial mat community in a Bahamian hypersaline lagoon. Appl. Environ. Microbiol. 1997, 63, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Herbst, D.B. Potential salinity limitations on nitrogen fixation in sediments from Mono Lake, California. Int. J. Salt Lake Res. 1998, 7, 261–274. [Google Scholar] [CrossRef]
- Namsaraev, Z.; Samylina, O.; Sukhacheva, M.; Borisenko, G.; Sorokin, D.Y.; Tourova, T. Effect of salinity on diazotrophic activity and microbial composition of phototrophic communities from Bitter-1 soda lake (Kulunda Steppe, Russia). Extremophiles 2018, 22, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Munson, T.O.; Burris, R.H. Nitrogen fixation by Rhodospirillum rubrum grown in nitrogen-limited continuous culture. J. Bacteriol. 1969, 97, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.; Cox, S.S.; Stegeman, R.A. Nitrogen fixation and nitrogenase activities in members of the family Rhodospirillaceae. J. Bacteriol. 1984, 157, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Oda, Y.; Samanta, S.K.; Rey, F.E.; Wu, L.; Liu, X.; Yan, T.; Zhou, J.; Harwood, C.S. Functional genomic analysis of three nitrogenase isozymes in the photosynthetic bacterium Rhodopseudomonas palustris. J. Bacteriol. 2005, 187, 7784–7794. [Google Scholar] [CrossRef] [Green Version]
- Kompantseva, E.I.; Komova, A.V.; Sorokin, D.Y. Communities of anoxygenic phototrophic bacteria in the saline soda lakes of the Kulunda Steppe (Altai Krai). Mikrobiologia 2010, 79, 89–95. [Google Scholar] [CrossRef]
- Kompantseva, E.I.; Komova, A.V.; Novikov, A.A.; Kostrikina, N.A. Rhodovulum tesquicola sp. nov., a haloalkaliphilic purple non-sulfur bacterium from brackish steppe soda lakes. Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 12, 2962–2966. [Google Scholar] [CrossRef] [Green Version]
- Hardy, R.W.; Holsten, R.D.; Jackson, E.K.; Burns, R. The acetylene-ethylene assay for N2 fixation: Laboratory and field evaluation. Plant Physiol. 1968, 43, 1185–1207. [Google Scholar] [CrossRef] [Green Version]
- Korzhenkov, A. ZGA: A flexible pipeline for read processing, de novo assembly and annotation of prokaryotic genomes. bioRxiv 2021. [Google Scholar] [CrossRef]
- BBMap—Bushnell B. Available online: Sourceforge.net/projects/bbmap/ (accessed on 12 April 2022).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, A.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Lomsadze, A.; Gemayel, K.; Tang, S.; Borodovsky, M. Modeling leaderless transcription and atypical genes results in more accurate gene prediction in prokaryotes. Genome Res. 2018, 29, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2019, 36, 2251–2252. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]
- Eren, A.M.; Kiefl, E.; Shaiber, A.; Veseli, I.; Miller, S.E.; Schechter, M.S.; Fink, I.; Pan, J.N.; Yousef, M.; Fogarty, E.C.; et al. Community-led, integrated, reproducible multi-omics with anvi’o. Nat. Microbiol. 2021, 6, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Lechner, M.; Findeiß, S.; Steiner, L.; Marz, M.; Stadler, P.F.; Prohaska, S.J. Proteinortho: Detection of (Co-)orthologs in large-scale analysis. BMC Bioinform. 2011, 12, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.H.; Tang, Y.J.; Blankenship, R.E. Carbon metabolic pathways in phototrophic bacteria and their broader evolutionary implications. Front. Microbiol. 2011, 2, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petushkova, E.; Mayorova, E.; Tsygankov, A. TCA Cycle Replenishing Pathways in Photosynthetic Purple Non-Sulfur Bacteria Growing with Acetate. Life 2021, 11, 711. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Falaleeva, M.I.; Lyashchenko, M.S.; Toropygin, I.Y.; Igamberdiev, A.U. Oligomeric forms of bacterial malate dehydrogenase: A study of the enzyme from the phototrophic non-sulfur bacterium Rhodovulum steppense A-20s. Biosci. Biotechnol. Biochem. 2018, 82, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Priefert, H.O.R.S.T.; Steinbüchel, A. Identification and molecular characterization of the acetyl coenzyme a synthetase gene (acoE) of Alcaligenes eutrophus. J. Bacteriol. 1992, 174, 6590–6599. [Google Scholar] [CrossRef] [Green Version]
- Crosby, H.A.; Rank, K.C.; Rayment, I.; Escalante-Semerena, J.C. Structure-guided expansion of the substrate range of methylmalonyl coenzyme a synthetase (MatB) of Rhodopseudomonas palustris. Appl. Environ. Microbiol. 2012, 78, 6619–6629. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wen, Q.; Harwood, C.S.; Liang, B.; Yang, J. A disjointed pathway for malonate degradation by Rhodopseudomonas palustris. Appl. Environ. Microbiol. 2020, 86, e00631-20. [Google Scholar] [CrossRef]
- Samylina, O.S.; Namsaraev, Z.B.; Grouzdev, D.S.; Slobodova, N.V.; Zelenev, V.V.; Borisenko, G.V.; Sorokin, D.Y. The patterns of nitrogen fixation in haloalkaliphilic phototrophic communities of Kulunda Steppe soda lakes (Altai, Russia). FEMS Microbiol. Ecol. 2019, 95, fiz174. [Google Scholar] [CrossRef]
- Tourova, T.P.; Slobodova, N.V.; Bumazhkin, B.K.; Sukhacheva, M.V.; Sorokin, D.Y. Diversity of diazotrophs in the sediments of saline and soda lakes analyzed with the use of the nifH gene as a molecular marker. Mikrobiologia 2014, 83, 634–647. [Google Scholar] [CrossRef]
- Roberts, G.P.; MacNeil, T.; MacNeil, D.; Brill, W.J. Regulation and characterization of protein products coded by the nif (nitrogen fixation) genes of Klebsiella pneumoniae. J. Bacteriol. 1978, 136, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Rangaraj, P.; Ryle, M.J.; Lanzilotta, W.N.; Ludden, P.W.; Shah, V.K. In vitro biosynthesis of iron-molybdenum cofactor and maturation of the nif-encoded apodinitrogenase: Effect of substitution for NifH with site-specifically altered forms of NifH. J. Biol. Chem. 1999, 274, 19778–19784. [Google Scholar] [CrossRef] [Green Version]
- Ribbe, M.W.; Bursey, E.H.; Burgess, B.K. Identification of an Fe protein residue (Glu146) of Azotobacter vinelandii nitrogenase that is specifically involved in FeMo cofactor insertion. J. Biol. Chem. 2000, 275, 17631–17638. [Google Scholar] [CrossRef] [Green Version]
- Avtges, P.; Kranz, R.G.; Haselkorn, R. Isolation and organization of genes for nitrogen fixation in Rhodopseudomonas capsulata. Mol. Gen. Genet. 1985, 201, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Schüddekopf, K.; Hennecke, S.; Liese, U.; Kutsche, M.; Klipp, W. Characterization of anf genes specific for the alternative nitrogenase and identification of nif genes required for both nitrogenases in Rhodobacter capsulatus. Mol. Microbiol. 1993, 8, 673–684. [Google Scholar] [CrossRef]
- Mus, F.; Alleman, A.B.; Pence, N.; Seefeldt, L.C.; Peters, J.W. Exploring the alternatives of biological nitrogen fixation. Metallomics 2018, 10, 523–538. [Google Scholar] [CrossRef]
- Scolnik, P.A.; Haselkorn, R. Activation of extra copies of genes coding for nitrogenase in Rhodopseudomonas capsulata. Nature 1984, 307, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.; Siefert, J.L.; Staples, C.R.; Blankenship, R.E. The natural history of nitrogen fixation. Mol. Biol. Evol. 2004, 21, 541–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koirala, A.; Brözel, V.S. Phylogeny of Nitrogenase Structural and Assembly Components Reveals New Insights into the Origin and Distribution of Nitrogen Fixation across Bacteria and Archaea. Microorganisms 2021, 9, 1662. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Pratte, B.S. Regulation of three nitrogenase gene clusters in the cyanobacterium Anabaena variabilis ATCC 29413. Life 2014, 4, 944–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraishi, A.; Ueda, Y. Isolation and characterization of Rhodovulum strictum sp. nov. and some other purple nonsulfur bacteria from colored blooms in tidal and seawater pools. Int. J. Syst. Evol. Microbiol. 1995, 45, 319–326. [Google Scholar]
- Grammann, K.; Volke, A.; Kunte, H.J. New type of osmoregulated solute transporter identified in halophilic members of the bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581T. J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzuki, M.; Moskvin, O.V.; Kuribayashi, M.; Sato, K.; Retamal, S.; Abo, M.; Zeilstra-Ryalls, J.; Gomelsky, M. Salt stress-induced changes in the transcriptome, compatible solutes, and membrane lipids in the facultatively phototrophic bacterium Rhodobacter sphaeroides. Appl. Environ. Microbiol. 2011, 77, 7551–7559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Keller, A.; Neulinger, S.C. Osmotic adaptation and compatible solute biosynthesis of phototrophic bacteria as revealed from genome analyses. Microorganisms 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed]







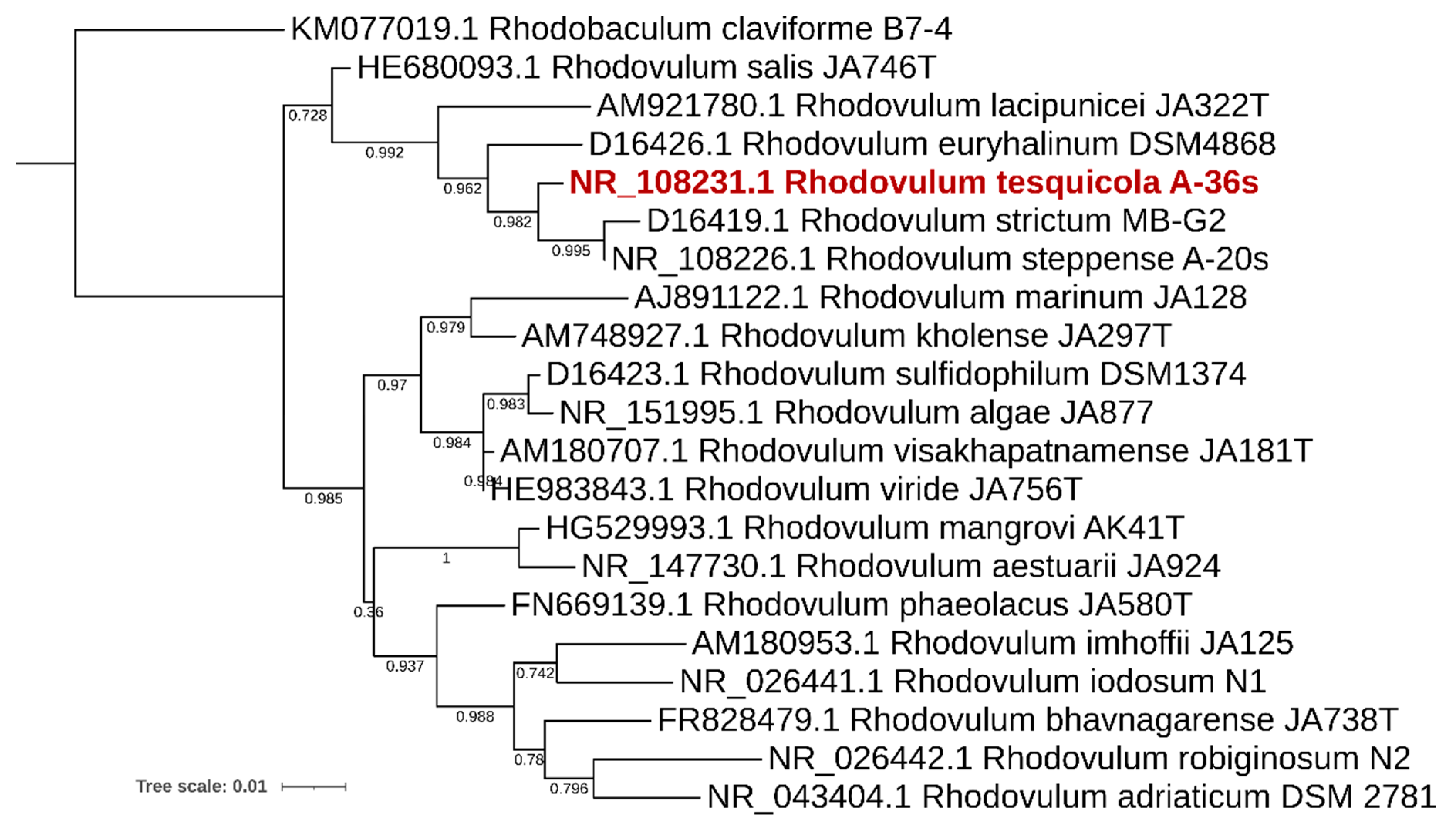{kind=link}
{kind=link}
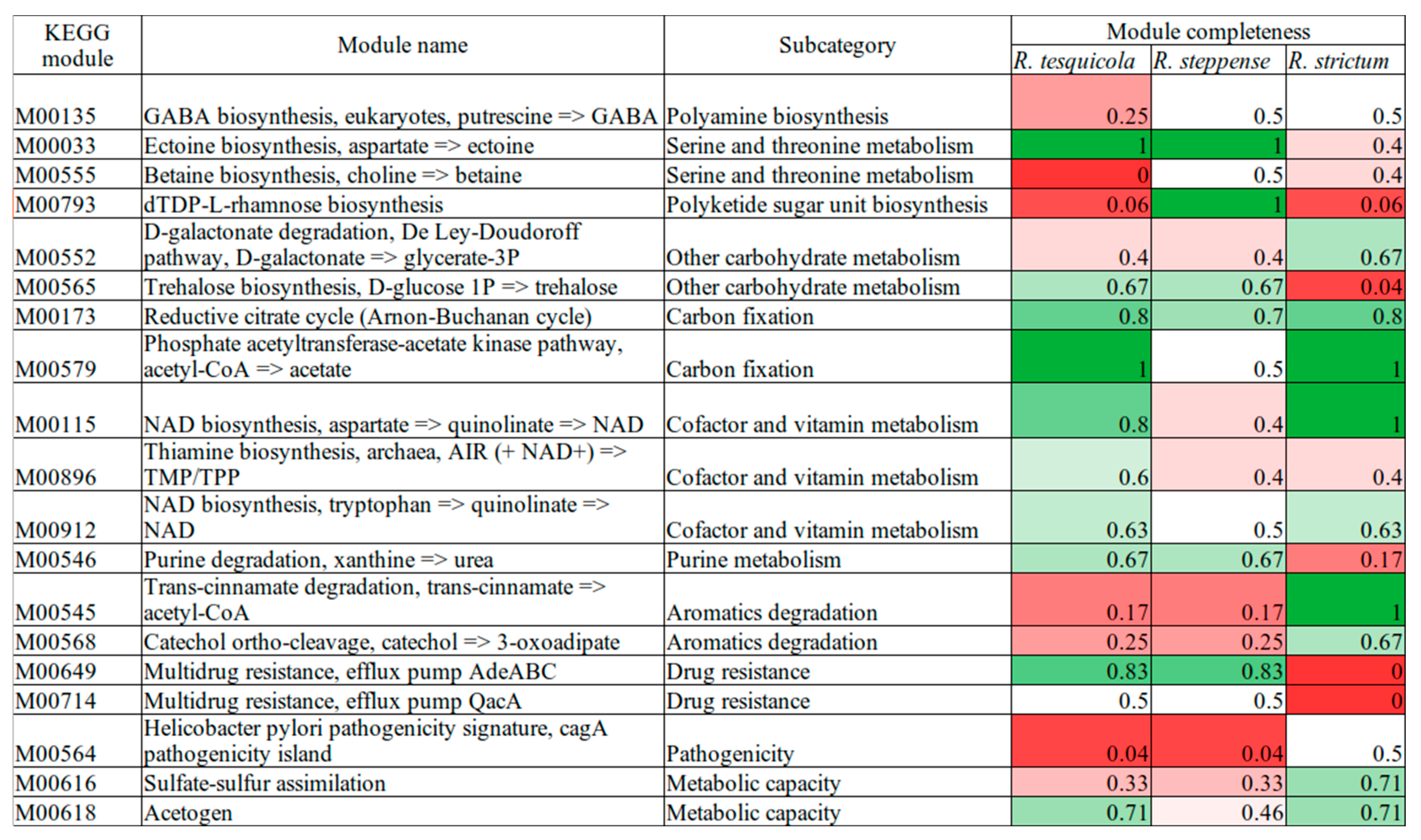{kind=link}
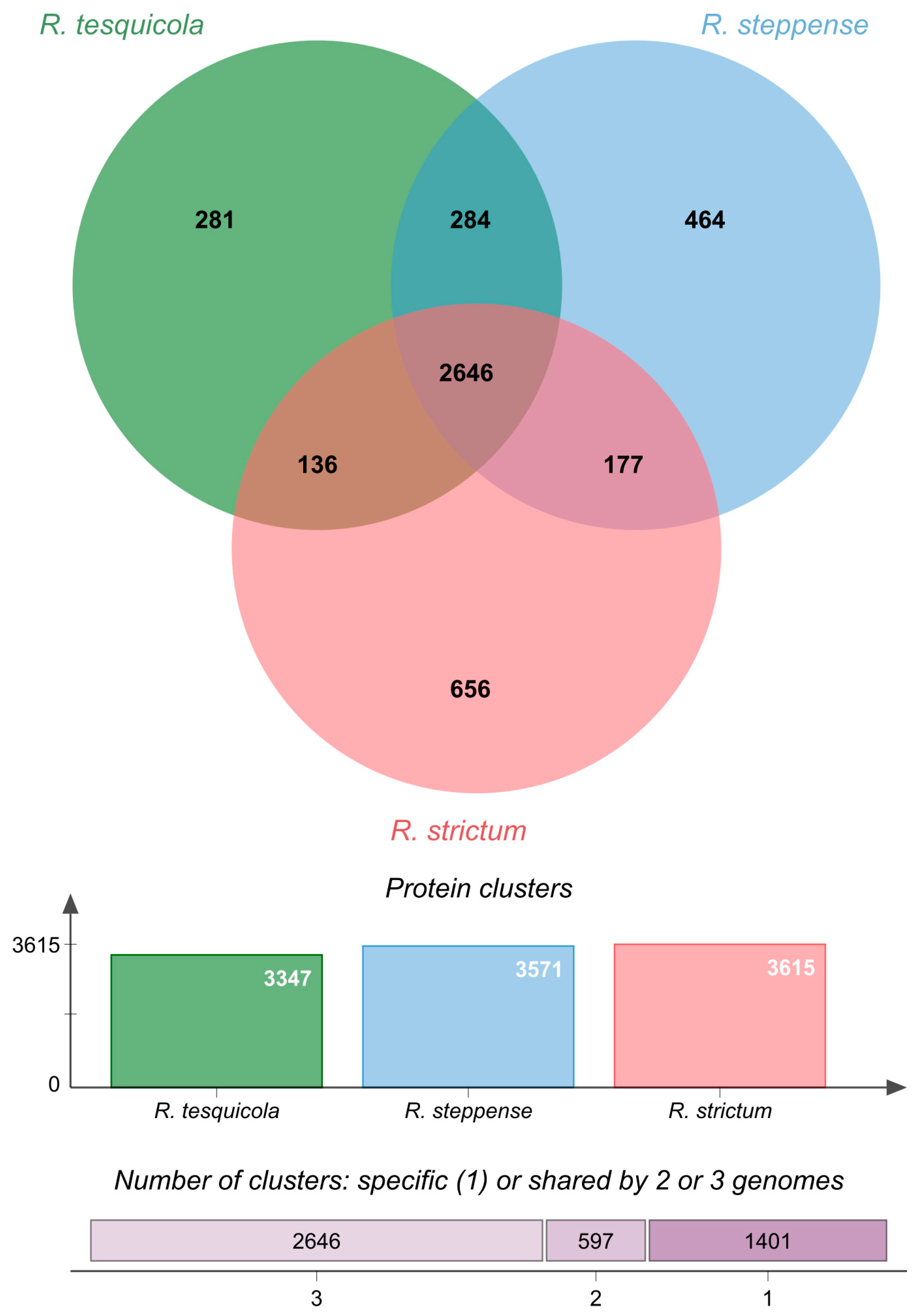{kind=link}
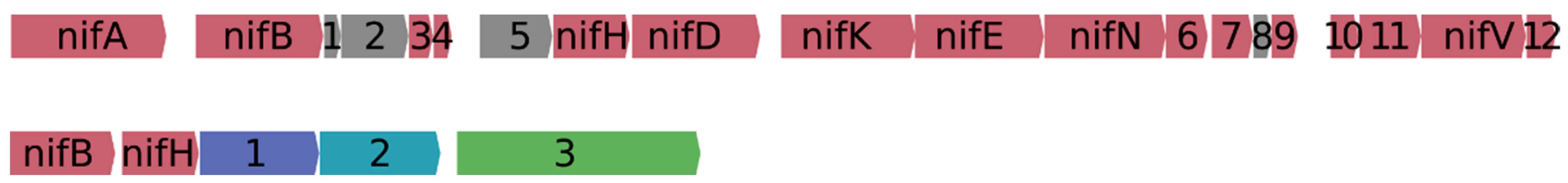{kind=link}
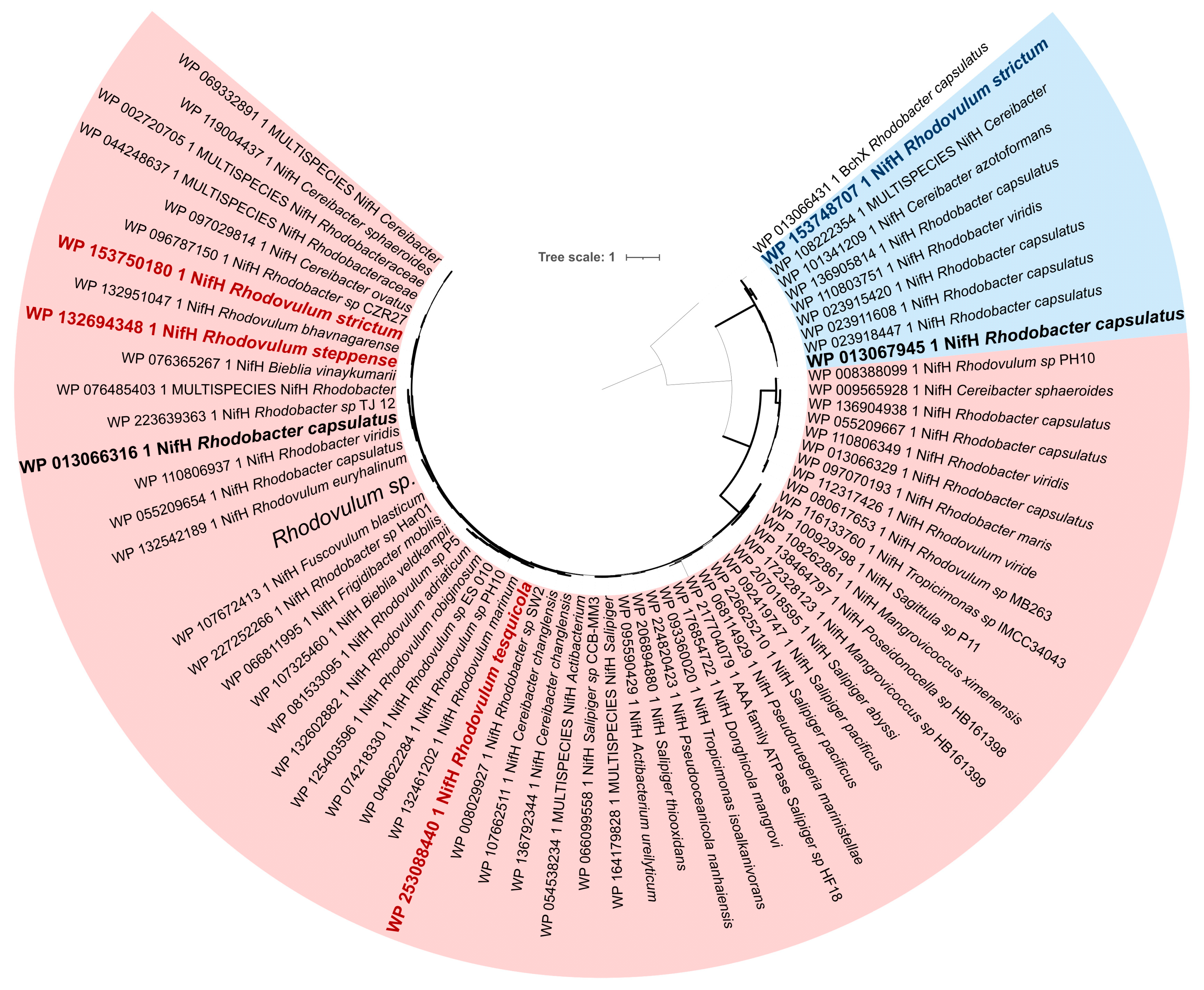{kind=link}
| Rhodovulum tesquicola A-36sT | Rhodovulum steppense A-20sT | Rhodovulum strictum DSM 11289T | |
|---|---|---|---|
| Genome size | 3,521,822 | 3,665,949 | 3,825,097 |
| Total number of genes | 3490 | 3634 | 3756 |
| Proteins | 3375 | 3582 | 3636 |
| rRNA genes | 1, 1, 1 (5S, 16S, 23S) | 1, 1, 2 (5S, 16S, 23S partial) | 1, 1, 1 (5S, 16S, 23S) |
| tRNA genes | 45 | 51 | 42 |
| Genome completeness, % | 99.41 | 99.12 | 98.83 |
| Genome contamination, % | 0 | 0.15 | 0.59 |
| R. tesquicola A-36sT | R. steppense A-20sT | R. strictum DSM 11289T | |
|---|---|---|---|
| pH range (optimum) | 7.5–10.0 (8.5–9.0) | 7.5–10.0 (8.5) | 7.5–9.0 (8.0) |
| NaCl range (optimum) (%) | 0.3–10.0 (1–3) | 0.5–10.0 (1–5) | 0.25–3.00 (0.8) |
| Vitamin requirement | b, paba, t, n | b, n, t | b, paba, t |
| Nitrogenase activity | +− | +− | + |
| Species | Nitrogenase Activity (nmoles C2H2/mg Dry Weight per Hour) | Reference |
|---|---|---|
| Rhodobacter capsulatus | 1430–4900 | [19] |
| Rhodobacter capsulatus B10 | 3650 | [19] |
| Cereibacter sphaeroides | 1110–2755 | [19] |
| Rhodovulum sulfidophilum W4 | 640 | [19] |
| Rhodothalassium salexigens DSM 2172 | 630 | [19] |
| Rhodomicrobium vannielii | 660–1570 | [19] |
| Rhodopseudomonas palustris | 480–700 | [19] |
| Rhodoblastus acidophilus | 500–900 | [19] |
| Rhodovulum tesquicola A-36sT | 278 | This work |
| Rhodovulum steppense A-20sT | 523 | This work |
| Rhodovulum strictum DSM 11289T | 4066 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komova, A.V.; Bakhmutova, E.D.; Izotova, A.O.; Kochetova, E.S.; Toshchakov, S.V.; Namsaraev, Z.B.; Golichenkov, M.V.; Korzhenkov, A.A. Nitrogen Fixation Activity and Genome Analysis of a Moderately Haloalkaliphilic Anoxygenic Phototrophic Bacterium Rhodovulum tesquicola. Microorganisms 2022, 10, 1615. https://doi.org/10.3390/microorganisms10081615
Komova AV, Bakhmutova ED, Izotova AO, Kochetova ES, Toshchakov SV, Namsaraev ZB, Golichenkov MV, Korzhenkov AA. Nitrogen Fixation Activity and Genome Analysis of a Moderately Haloalkaliphilic Anoxygenic Phototrophic Bacterium Rhodovulum tesquicola. Microorganisms. 2022; 10(8):1615. https://doi.org/10.3390/microorganisms10081615
Chicago/Turabian StyleKomova, Anastasia V., Elizaveta D. Bakhmutova, Anna O. Izotova, Evelina S. Kochetova, Stepan V. Toshchakov, Zorigto B. Namsaraev, Maxim V. Golichenkov, and Aleksei A. Korzhenkov. 2022. "Nitrogen Fixation Activity and Genome Analysis of a Moderately Haloalkaliphilic Anoxygenic Phototrophic Bacterium Rhodovulum tesquicola" Microorganisms 10, no. 8: 1615. https://doi.org/10.3390/microorganisms10081615