
Chromatic Acclimation Processes and Their Relationships with Phycobiliprotein Complexes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Selected Cyanobacteria Genomes and Predicated Photosensory Proteins
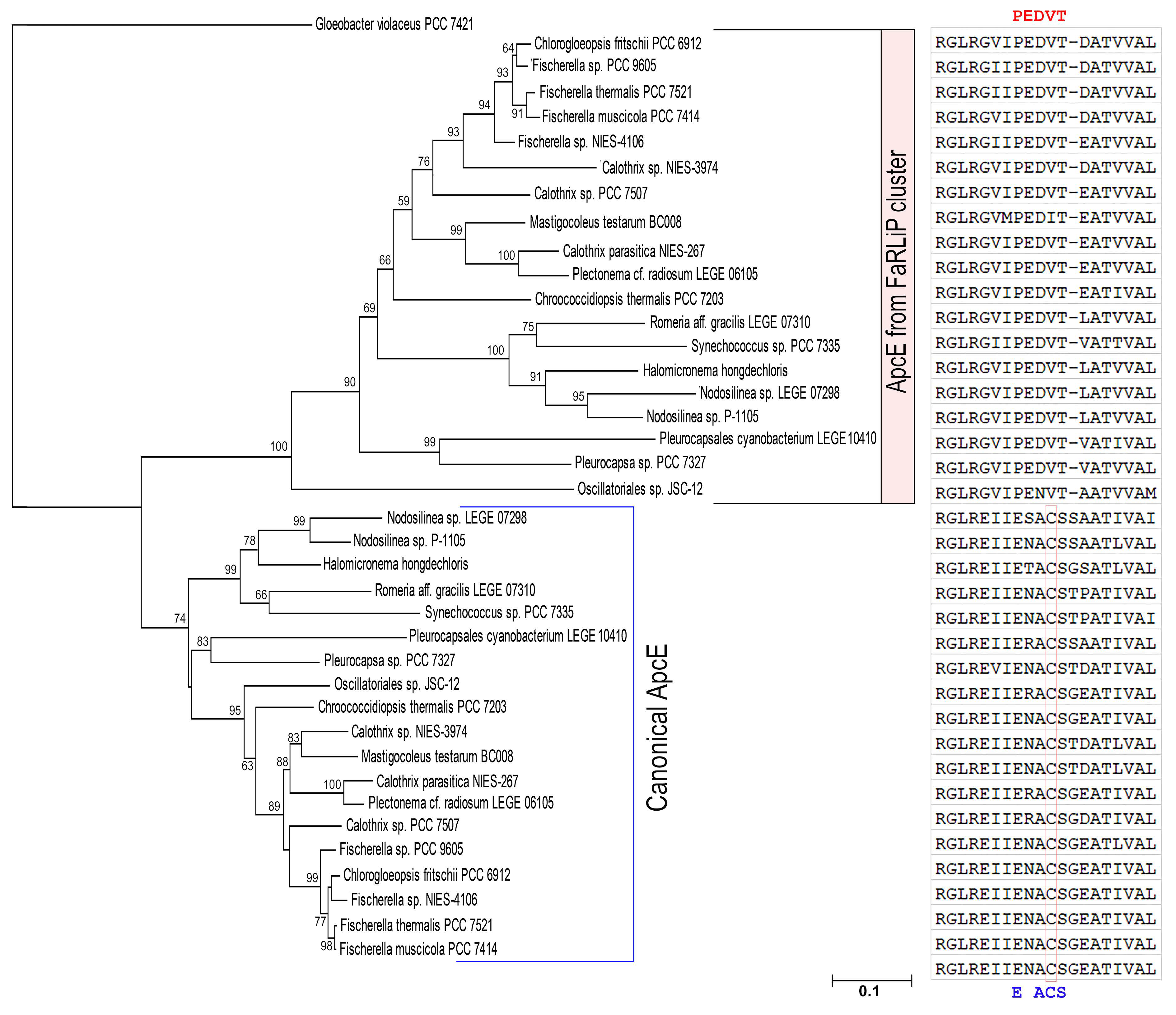
2.2. Sequence Homology and Phylogenetic Analysis
3. Results and Discussion
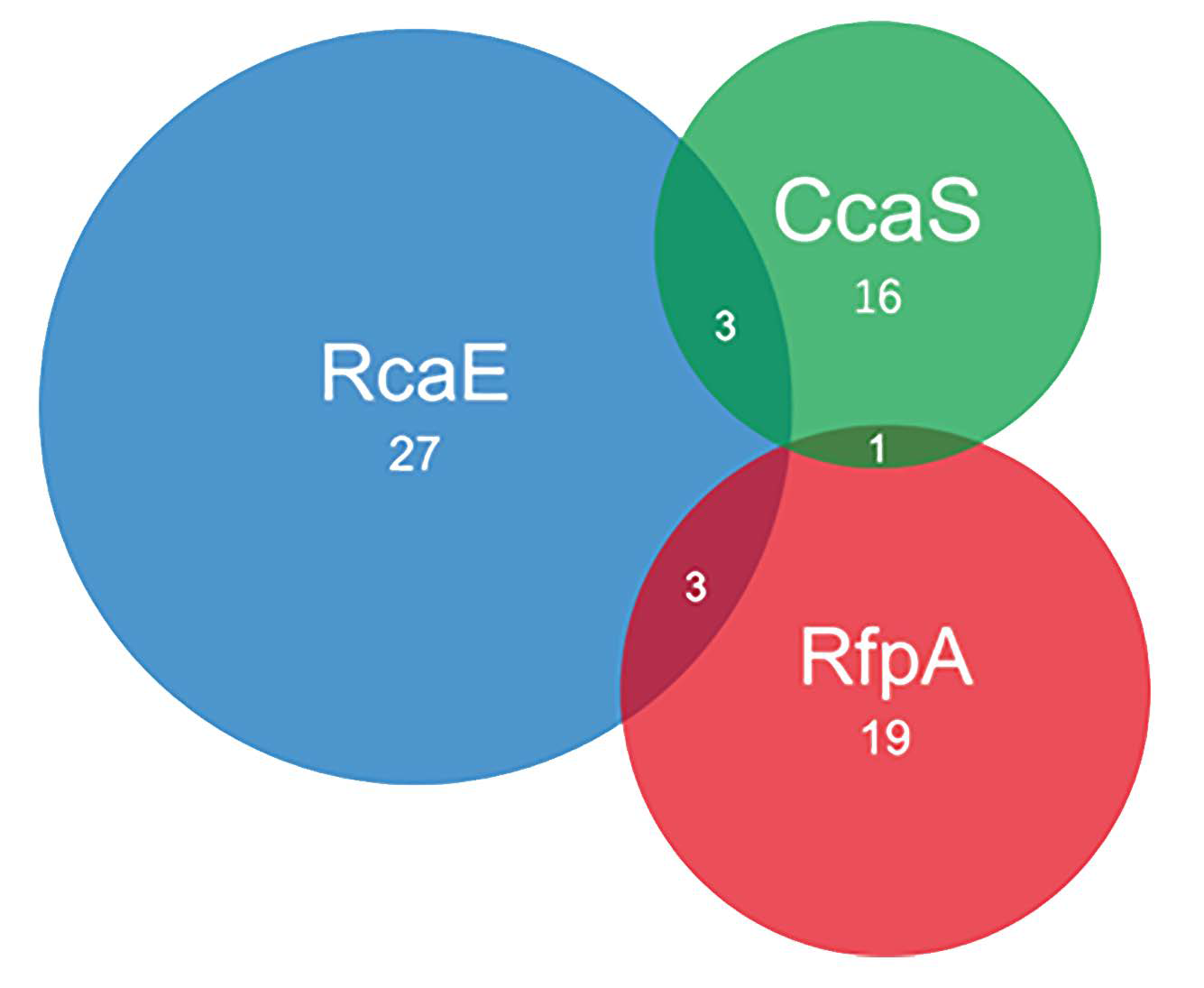
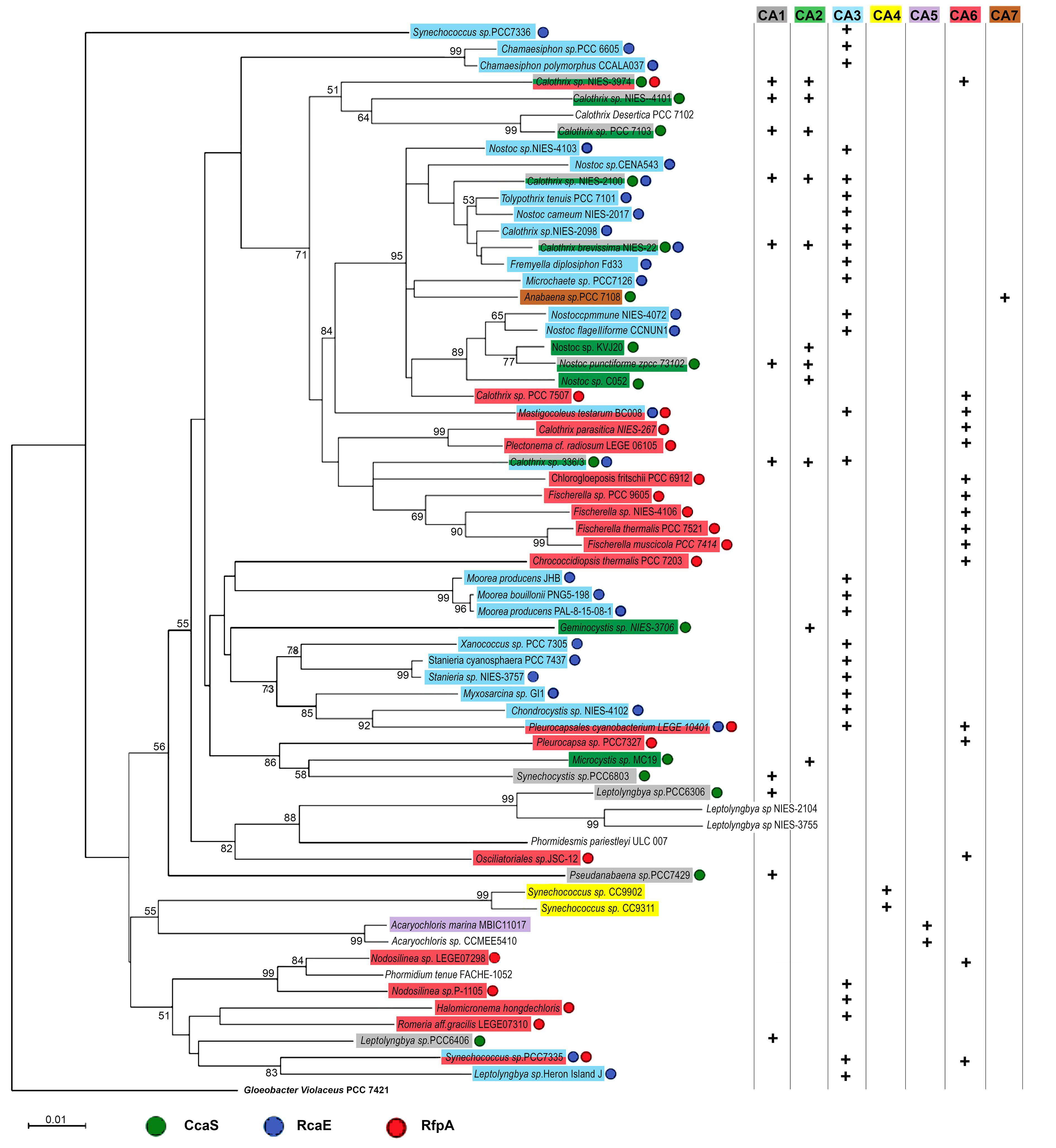
3.1. Photoreceptors and CA-Capable Cyanobacteria
3.1.1. Predication of Photoreceptors
3.1.2. Characterization of CA-Capabilities
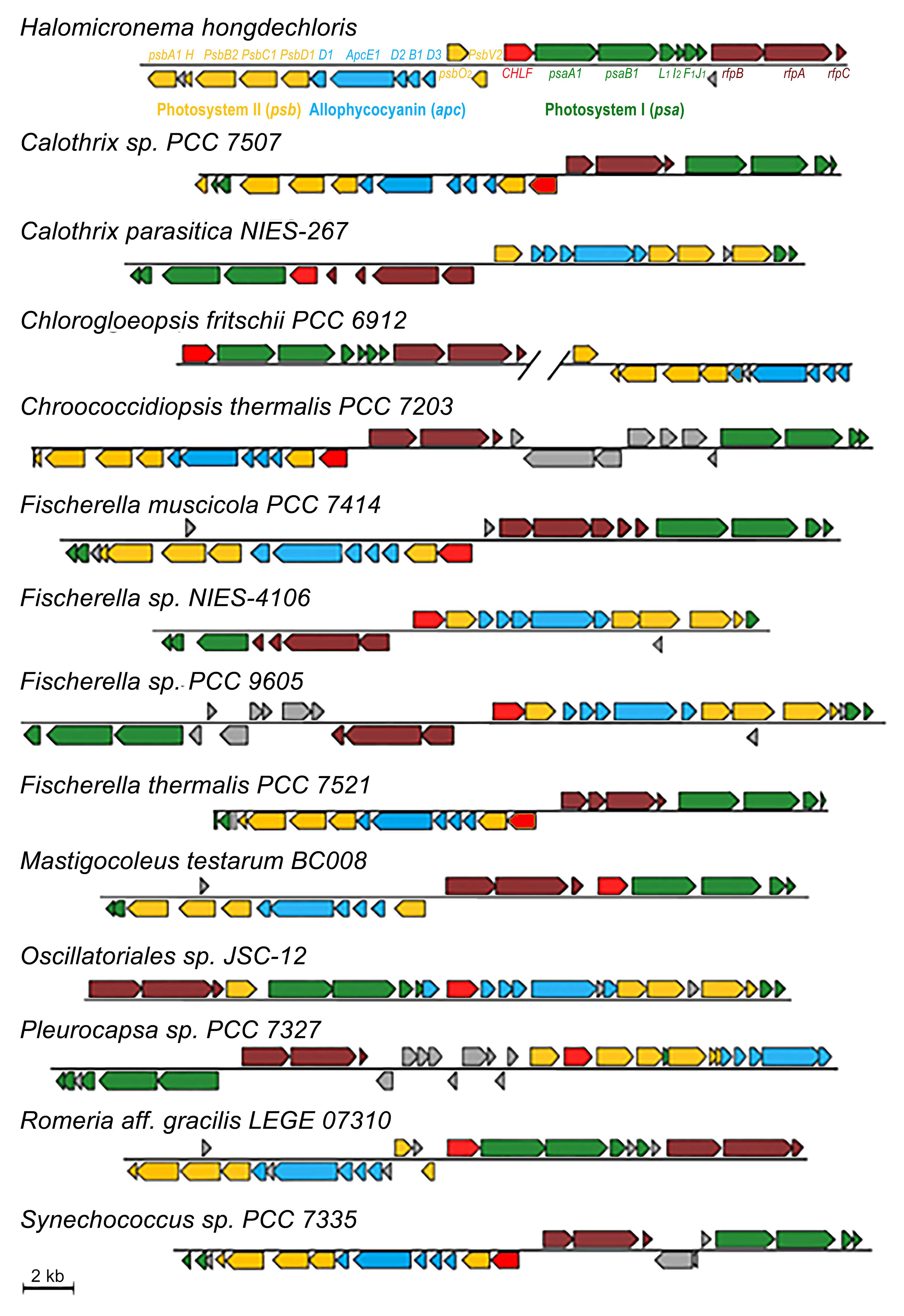
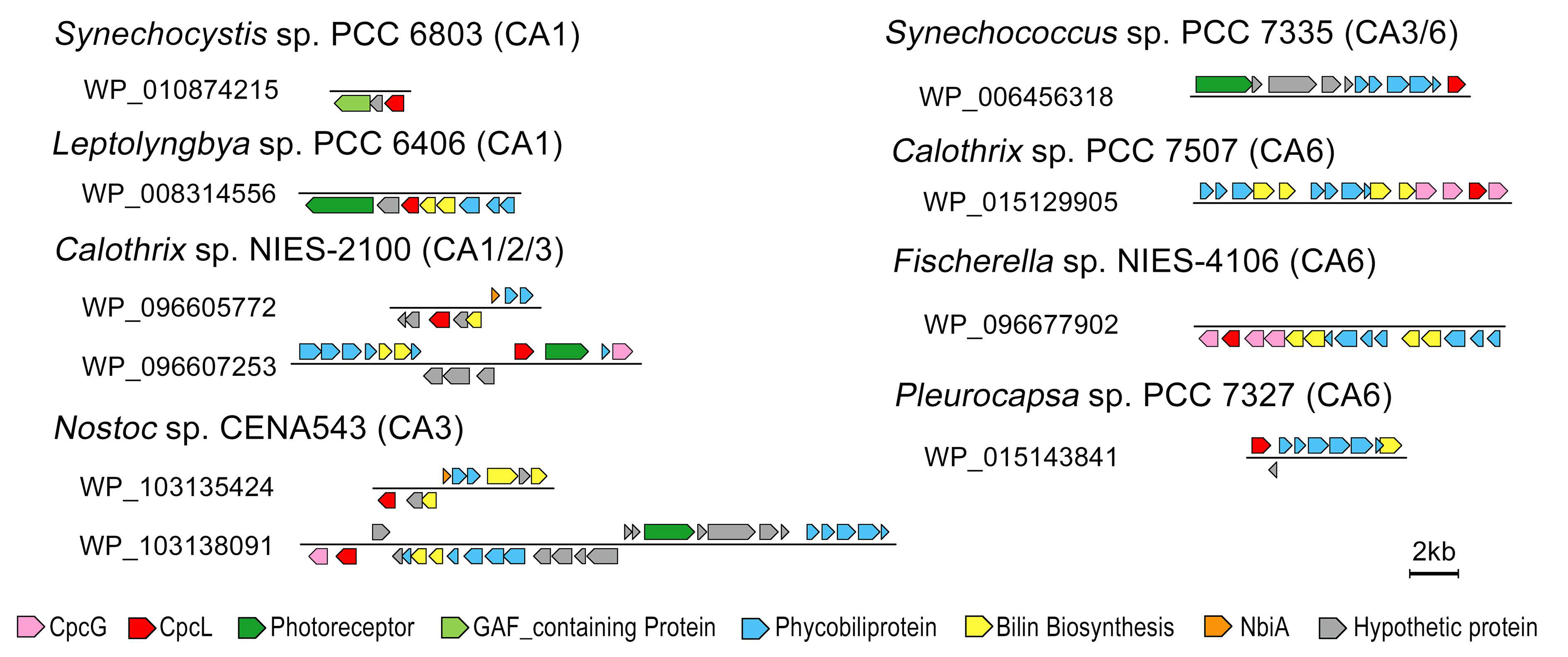
3.2. Cyanobacterial PBS Structure and CA Responses
3.2.1. Changeable PE Rods
3.2.2. Assembling CpcL-Rod PBS Structure
3.2.3. PBS Having Red-Shifted Absorption Characteristics
3.3. Photoacclimation and Alternative PBS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitton, B.A.; Potts, M. Introduction to the Cyanobacteria. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 1–11. [Google Scholar]
- Gutu, A.; Kehoe, D.M. Emerging perspectives on the mechanisms, regulation, and distribution of light color acclimation in cyanobacteria. Mol. Plant 2012, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacColl, R. Cyanobacterial phycobilisomes. J. Struct. Biol. 1998, 124, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Six, C.; Thomas, J.C.; Thion, L.; Lemoine, Y.; Zal, F.; Partensky, F. Two novel phycoerythrin-associated linker proteins in the marine cyanobacterium Synechococcus sp. strain WH8102. J. Bacteriol. 2005, 187, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Peng, P.; Dong, L.; Sun, Y.; Zeng, X.; Ding, W.; Scheer, H.; Yang, X.; Zhao, K.H. The structure of allophycocyanin B from Synechocystis PCC 6803 reveals the structural basis for the extreme redshift of the terminal emitter in phycobilisomes. Acta Crystallogr. D 2014, 70, 2558–2569. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, H.; Niedzwiedzki, D.M.; Prado, M.; He, G.; Gross, M.L.; Blankenship, R.E. Phycobilisomes supply excitations to both photosystems in a megacomplex in cyanobacteria. Science 2013, 342, 1104–1107. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Ikeuchi, M. Phycobilisome: Architecture of a light-harvesting supercomplex. Photosynth. Res. 2013, 116, 265–276. [Google Scholar] [CrossRef]
- Bryant, D.A.; Canniffe, D.P. How nature designs light-harvesting antenna systems: Design principles and functional realization in chlorophototrophic prokaryotes. J. Phys. B At. Mol. Opt. 2018, 51, 033001. [Google Scholar] [CrossRef]
- Federspiel, N.A.; Grossman, A.R. Characterization of the light-regulated operon encoding the phycoerythrin-associated linker proteins from the cyanobacterium Fremyella diplosiphon. J. Bacteriol. 1990, 172, 4072–4081. [Google Scholar] [CrossRef] [Green Version]
- Bhaya, D. In the limelight: Photoreceptors in cyanobacteria. MBio 2016, 7, e00741-16. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Chihong, S.; Watanabe, M.; Yonekawa, C.; Murata, K.; Ikeuchi, M.; Eki, T. Diverse chromatic acclimation processes regulating phycoerythrocyanin and rod-shaped phycobilisome in cyanobacteria. Mol. Plant 2019, 12, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, J.E.; Garczarek, L.; Partensky, F.; Kehoe, D.M. Chromatic acclimation in cyanobacteria: A diverse and widespread process for optimizing photosynthesis. Annu. Rev. Microbiol. 2019, 73, 407–433. [Google Scholar] [CrossRef] [PubMed]
- Kehoe, D.M.; Gutu, A. Responding to color: The regulation of complementary chromatic adaptation. Annu. Rev. Plant. Biol. 2006, 57, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Everroad, C.; Six, C.; Partensky, F.; Thomas, J.-C.; Holtzendorff, J.; Wood, A.M. Biochemical bases of type IV chromatic adaptation in marine Synechococcus spp. J. Bacteriol. 2006, 188, 3345–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, A.; Biswas, A.; Blot, N.; Partensky, F.; Karty, J.A.; Hammad, L.A.; Garczarek, L.; Gutu, A.; Schluchter, W.M.; Kehoe, D.M. Phycoerythrin-specific bilin lyase-isomerase controls blue-green chromatic acclimation in marine Synechococcus. Proc. Natl. Acad. Sci. USA 2012, 109, 20136–20141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duxbury, Z.; Schliep, M.; Ritchie, R.J.; Larkum, A.W.D.; Chen, M. Chromatic photoacclimation extends utilisable photosynthetically active radiation in the chlorophyll d-containing cyanobacterium, Acaryochloris marina. Photosynth. Res. 2009, 101, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, J.E.; Nguyen, A.A.; Karty, J.A.; Shukla, A.; Schluchter, W.M.; Garczarek, L.; Partensky, F.; Kehoe, D.M. Self-regulating genomic island encoding tandem regulators confers chromatic acclimation to marine Synechococcus. Proc. Natl. Acad. Sci. USA 2016, 113, 6077–6082. [Google Scholar] [CrossRef] [Green Version]
- Wiltbank, L.B.; Kehoe, D.M. Diverse light responses of cyanobacteria mediated by phytochrome superfamily photoreceptors. Nat. Rev. Microbiol. 2019, 17, 37–50. [Google Scholar] [CrossRef]
- Hirose, Y.; Narikawa, R.; Katayama, M.; Ikeuchi, M. Cyanobacteriochrome CcaS regulates phycoerythrin accumulation in Nostoc punctiforme, a group II chromatic adapter. Proc. Natl. Acad. Sci. USA 2010, 107, 8854–8859. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, D.M.; Grossman, A.R. New classes of mutants in complementary chromatic adaptation provide evidence for a novel four-step phosphorelay system. J. Bacteriol. 1997, 179, 3914–3921. [Google Scholar] [CrossRef] [Green Version]
- Terauchi, K.; Montgomery, B.L.; Grossman, A.R.; Lagarias, J.C.; Kehoe, D.M. RcaE is a complementary chromatic adaptation photoreceptor required for green and red light responsiveness. Mol. Microbiol. 2004, 51, 567–577. [Google Scholar] [CrossRef]
- Kondo, K.; Geng, X.X.; Katayama, M.; Ikeuchi, M. Distinct roles of CpcG1 and CpcG2 in phycobilisome assembly in the cyanobacterium Synechocystis sp. PCC 6803. Photosynth. Res. 2005, 84, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Ochiai, Y.; Katayama, M.; Ikeuchi, M. The Membrane-associated CpcG2-phycobilisome in Synechocystis: A new photosystem I antenna. Plant Physiol. 2007, 144, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marsac, N.T.; Cohen-bazire, G. Molecular composition of cyanobacterial phycobilisomes. Proc. Natl. Acad. Sci. USA 1977, 74, 1635–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Lin, Y.; Garvey, C.J.; Birch, D.; Corkery, R.W.; Loughlin, P.C.; Scheer, H.; Willows, R.D.; Chen, M. Characterization of red-shifted phycobilisomes isolated from the chlorophyll f-containing cyanobacterium Halomicronema hongdechloris. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, F.; Shen, G.; Bryant, D. Occurrence of Far-Red Light Photoacclimation (FaRLiP) in diverse cyanobacteria. Life 2014, 5, 4–24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Gan, F.; Shen, G.; Bryant, D.A. RfpA, RfpB, and RfpC are the master control elements of far-red light photoacclimation (FaRLiP). Front. Microbiol. 2015, 6, 1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Hernandez-Prieto, M.A.; Loughlin, P.C.; Li, Y.; Willows, R.D. Genome and proteome of the chlorophyll f-producing cyanobacterium Halomicronema hongdechloris: Adaptative proteomic shifts under different light conditions. BMC Genom. 2019, 20, 207. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Zhang, S.; Rockwell, N.; Martin, S.; Lagarias, J.; Bryant, D. Extensive remodeling of a cyanobacterial photosynthetic apparatus in far-red light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Rockwell, N.C.; Nishiyama, K.; Narikawa, R.; Ukaji, Y.; Inomata, K.; Lagarias, J.C.; Ikeuchi, M. Green/red cyanobacteriochromes regulate complementary chromatic acclimation via a protochromic photocycle. Proc. Natl. Acad. Sci. USA 2013, 110, 4974–4979. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Kikukawa, T.; Miyoshi, R.; Kajimoto, K.; Yonekawa, C.; Fujisawa, T.; Unno, M.; Eki, T.; Hirose, Y. Protochromic absorption changes in the two-cysteine photocycle of a blue/orange cyanobacteriochrome. J. Biol. Chem. 2019, 294, 18909–18922. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.-Y.; Gan, F.; Shen, G.; Bryant, D.A. Far-red light photoacclimation (FaRLiP) in Synechococcus sp. PCC 7335. II. Characterization of phycobiliproteins produced during acclimation to far-red light. Photosynth. Res. 2017, 131, 187–202. [Google Scholar] [CrossRef]
- Hirose, Y.; Shimada, T.; Narikawa, R.; Katayama, M.; Ikeuchi, M. Cyanobacteriochrome CcaS is the green light receptor that induces the expression of phycobilisome linker protein. Proc. Natl. Acad. Sci. USA 2008, 105, 9528–9533. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Semchonok, D.A.; Webber-Birungi, M.T.; Ehira, S.; Kondo, K.; Narikawa, R.; Ohmori, M.; Boekema, E.J.; Ikeuchi, M. Attachment of phycobilisomes in an antenna–photosystem I supercomplex of cyanobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, 2512–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Floetenmeyer, M.; Bibby, T.S. Supramolecular organization of phycobiliproteins in the chlorophyll d-containing cyanobacterium Acaryochloris marina. FEBS Lett. 2009, 583, 2535–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.-Y.; Gan, F.; Shen, G.; Zhao, C.; Bryant, D.A. Far-red light photoacclimation (FaRLiP) in Synechococcus sp. PCC 7335: I. Regulation of FaRLiP gene expression. Photosynth. Res. 2017, 131, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, M. The specificity of the bilin lyase CpcS for chromophore attachment to allophycocyanin in the chlorophyll f-containing cyanobacterium Halomicronima hongdechloris. Photosynth. Res. 2021, 151, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Soulier, N.; Laremore, T.N.; Bryant, D.A. Characterization of cyanobacterial allophycocyanins absorbing far-red light. Photosynth. Res. 2020, 145, 189–207. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ishizuka, T. Cyanobacteriochromes: A new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria. Photochem. Photobiol. Sci. 2008, 7, 1159. [Google Scholar] [CrossRef]
- Wiltbank, L.B.; Kehoe, D.M. Two cyanobacterial photoreceptors regulate photosynthetic light harvesting by sensing teal, green, yellow, and red light. MBio 2016, 7, e02130-15. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A. Light signals, phytochromes and crosstalk with other environmental cues. J. Exp. Bot. 2003, 55, 271–276. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyanobacteria | Photoreceptors | CA Types | Phycobiliproteins | Linker Proteins | |||
|---|---|---|---|---|---|---|---|
| Calothrix sp. NIES-3974 | CcaS/RfpA | CA1 | CA2 | CA6 | APC/PC/PE | ApcE×2/CpcG×3/CpcL×2 | |
| Calothrix sp. NIES-2100 | CcaS/RcaE | CA1 | CA2 | CA3 | APC/PC/PE | ApcE×1/CpcG×1/CpcL×2 | |
| Calothrix brevissima NIES-22 | CcaS/RcaE | CA1 | CA2 | CA3 | APC/PC/PE | ApcE×1/CpcG×2/CpcL×2 | |
| Calothrix sp. 336/3 | CcaS/RcaE | CA1 | CA2 | CA3 | APC/PC/PE | ApcE×1/CpcG×2/CpcL×2 | |
| Calothrix sp. 336/3 | CcaS/RcaE | CA1 | CA2 | CA3 | APC/PC/PE | ApcE×1/CpcG×2/CpcL×2 | |
| Mastigocoleus testarum BC008 | RcaE/RfpA | CA3 | CA6 | APC/PC/PE | ApcE×3/CpcG×1/CpcL×2 | ||
| Pleurocapsales cyanobacterium LEGE 10410 | RcaE/RfpA | CA3 | CA6 | APC/PC/PE | ApcE×2/CpcG×1 | ||
| Synechococcus sp. PCC 7335 | RcaE/RfpA | CA3 | CA6 | APC/PC/PE | ApcE×2/CpcG×1/CpcL×1 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Chen, M. Chromatic Acclimation Processes and Their Relationships with Phycobiliprotein Complexes. Microorganisms 2022, 10, 1562. https://doi.org/10.3390/microorganisms10081562
Wang F, Chen M. Chromatic Acclimation Processes and Their Relationships with Phycobiliprotein Complexes. Microorganisms. 2022; 10(8):1562. https://doi.org/10.3390/microorganisms10081562
Chicago/Turabian StyleWang, Fanyue, and Min Chen. 2022. "Chromatic Acclimation Processes and Their Relationships with Phycobiliprotein Complexes" Microorganisms 10, no. 8: 1562. https://doi.org/10.3390/microorganisms10081562