
Evidence for Electron Transfer from the Bidirectional Hydrogenase to the Photosynthetic Complex I (NDH-1) in the Cyanobacterium Synechocystis sp. PCC 6803

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Construction of Mutants
2.3. Oxygen Measurements
2.4. PhotoH2 Measurements
2.5. Hydrogen Uptake
2.6. Electron Flow Determination through Photosystem I
2.7. State Change Measurements
2.8. Confocal Laser Scanning Microscopy and Colocalization
3. Results
3.1. Dark Respiration
3.2. PhotoH2 Production
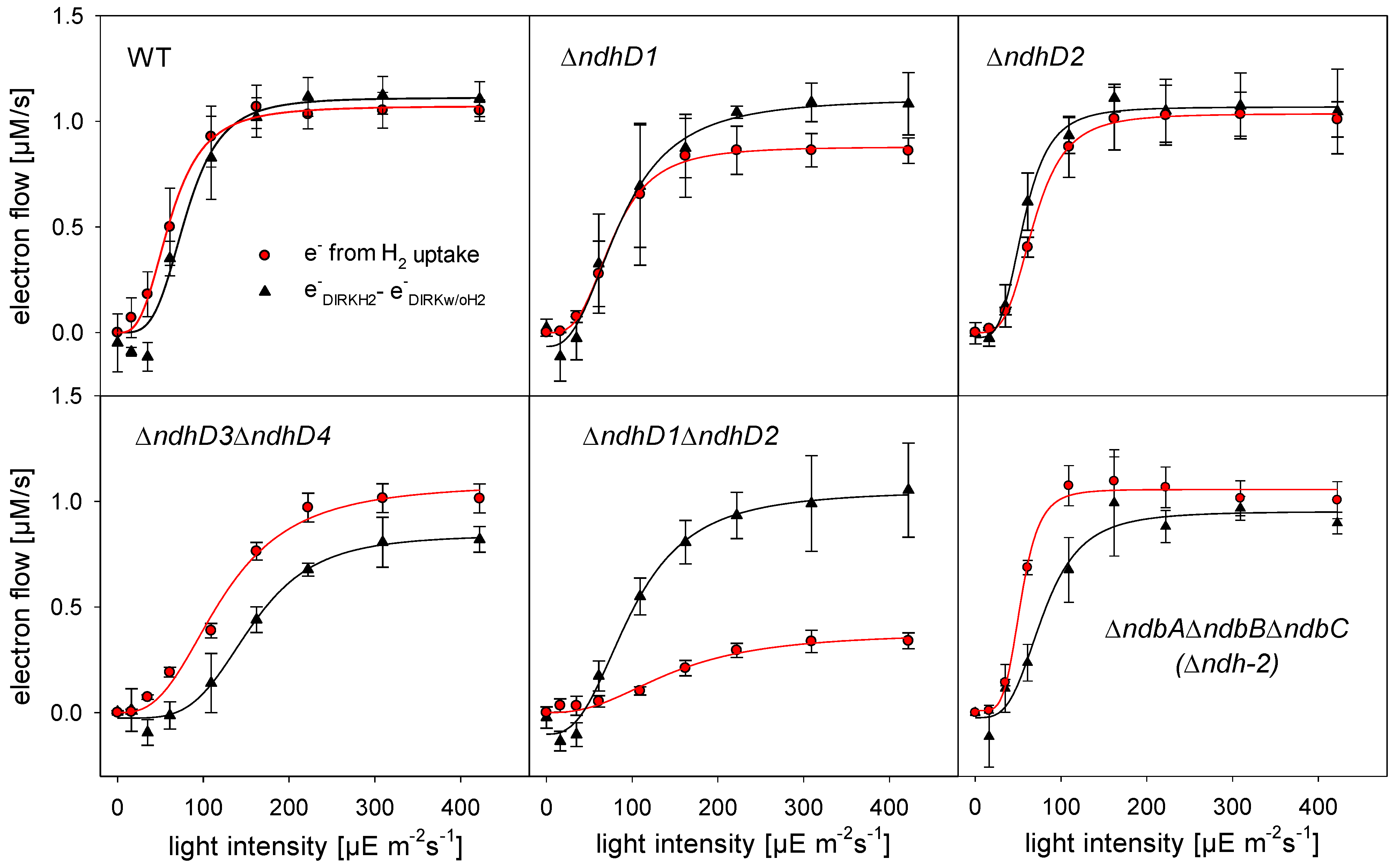
3.3. Electron Flow through PSI during H2 Uptake
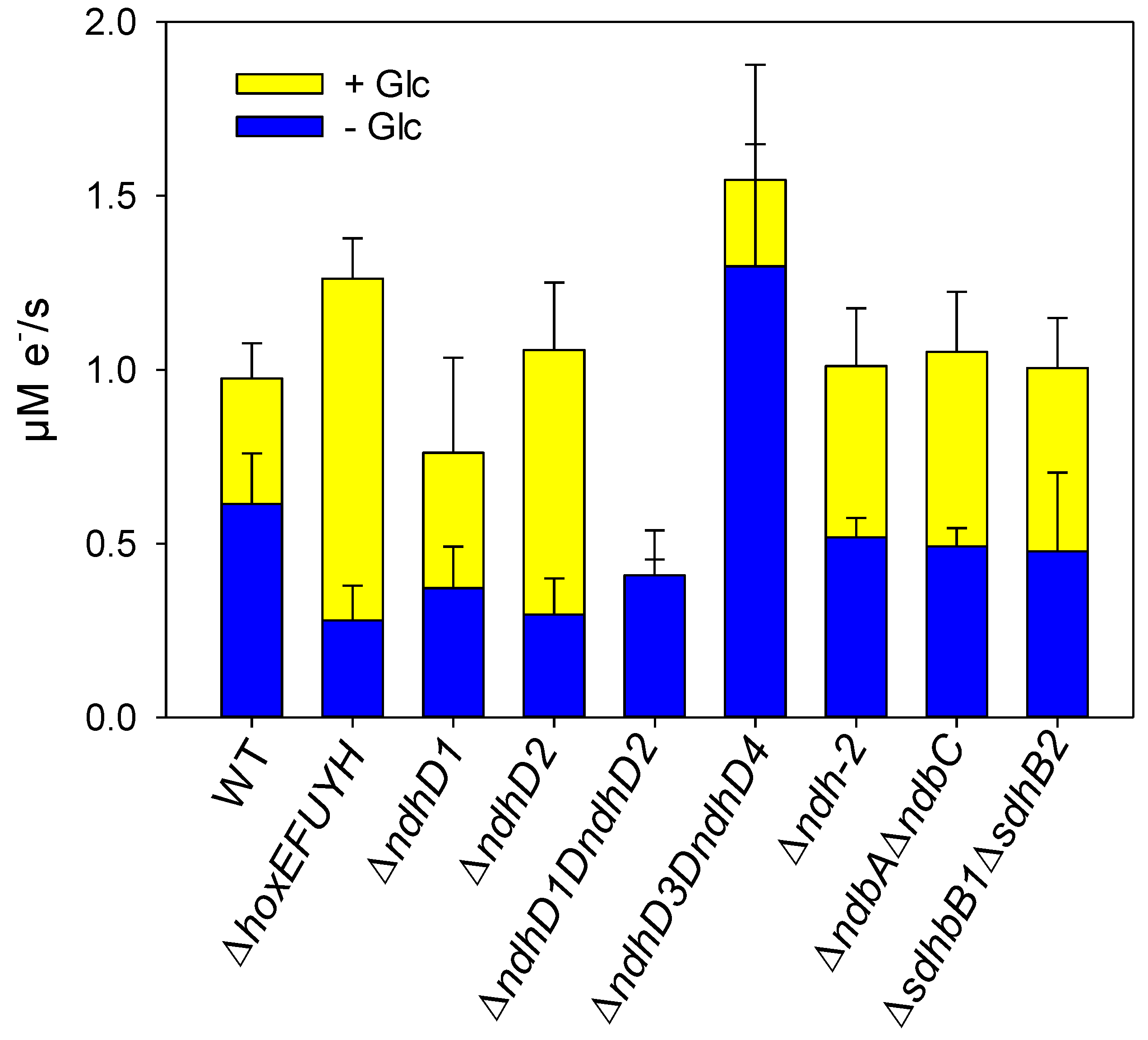
3.4. Glucose Utilization in the Light
3.5. State Change Due to Glucose Addition
3.6. Effect of Glucose Addition on Electron Transport When PSII Is Inhibited by DCMU
3.7. Colocalization of HoxF and NdhM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sazanov, L.A. A giant molecular proton pump: Structure and mechanism of respiratory complex I. Nat. Rev. Mol. Cell Biol. 2015, 16, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Schuller, J.M.; Birrell, J.A.; Tanaka, H.; Konuma, T.; Wulflhorst, H.; Cox, N.; Schuller, S.K.; Thiemann, J.; Lubitz, W.; Setif, P.; et al. Structural adaptations of photosynthetic complex I enable ferredoxin-dependent electron transfer. Science 2019, 363, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai, T. Chloroplast NDH: A different enzyme with a structure similar to that of respiratory NADH dehydrogenase. Biochim. Biophys. Acta-Bioenerg. 2016, 1857, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shuai, J.; Ran, Z.; Zhao, J.; Wu, Z.; Liao, R.; Wu, J.; Ma, W.; Lei, M. Structural insights into NDH-1 mediated cyclic electron transfer. Nat. Commun. 2020, 11, 888. [Google Scholar] [CrossRef] [Green Version]
- Appel, J.; Schulz, R. Sequence analysis of an operon of a NAD(P)-reducing nickel hydrogenase from the cyanobacterium Synechocystis sp PCC 6803 gives additional evidence for direct coupling of the enzyme to NAD(P)H-dehydrogenase (complex I). Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1996, 1298, 141–147. [Google Scholar] [CrossRef]
- Schmitz, O.; Bothe, H. The diaphorase subunit HoxU of the bidirectional hydrogenase as electron transferring protein in cyanobacterial respiration? Naturwissenschaften 1996, 83, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Schut, G.J.; Zadvornyy, O.; Wu, C.H.; Peters, J.W.; Boyd, E.S.; Adams, M.W. The role of geochemistry and energetics in the evolution of modern respiratory complexes from a proton-reducing ancestor. Biochim. Biophys. Acta 2016, 1857, 958–970. [Google Scholar] [CrossRef]
- Yu, H.; Schut, G.J.; Haja, D.K.; Adams, M.W.W.; Li, H. Evolution of complex I–like respiratory complexes. J. Biol. Chem. 2021, 296, 100740. [Google Scholar] [CrossRef] [PubMed]
- Greening, C.; Biswas, A.; Carere, C.R.; Jackson, C.J.; Taylor, M.C.; Stott, M.B.; Cook, G.M.; Morales, S.E. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 2016, 10, 761–777. [Google Scholar] [CrossRef] [Green Version]
- Greening, C.; Grinter, R. Microbial oxidation of atmospheric trace gases. Nat. Rev. Microbiol. 2022. [Google Scholar] [CrossRef]
- Cournac, L.; Guedeney, G.; Peltier, G.; Vignais, P.M. Sustained photoevolution of molecular hydrogen in a mutant of Synechocystis sp. strain PCC 6803 deficient in the type I NADPH-dehydrogenase complex. J. Bacteriol. 2004, 186, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Gutekunst, K.; Schulz, R. CHAPTER 4 The Physiology of the Bidirectional NiFe-hydrogenase in Cyanobacteria and the Role of Hydrogen Throughout the Evolution of Life. In Microalgal Hydrogen Production: Achievements and Perspectives; The Royal Society of Chemistry: London, UK, 2018; pp. 107–138. [Google Scholar]
- Gutthann, F.; Egert, M.; Marques, A.; Appel, J. Inhibition of respiration and nitrate assimilation enhances photohydrogen evolution under low oxygen concentrations in Syneehocystis sp PCC 6803. Biochim. Biophys. Acta-Bioenerg. 2007, 1767, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Gutekunst, K.; Chen, X.; Schreiber, K.; Kaspar, U.; Makam, S.; Appel, J. The bidirectional NiFe-hydrogenase in Synechocystis sp. PCC 6803 is reduced by flavodoxin and ferredoxin and is essential under mixotrophic, nitrate-limiting conditions. J. Biol. Chem. 2014, 289, 1930–1937. [Google Scholar] [CrossRef] [Green Version]
- Artz, J.H.; Tokmina-Lukaszewska, M.; Mulder, D.W.; Lubner, C.E.; Gutekunst, K.; Appel, J.; Bothner, B.; Boehm, M.; King, P.W. The structure and reactivity of the HoxEFU complex from the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 2020, 295, 9445–9454. [Google Scholar] [CrossRef]
- Aubert-Jousset, E.; Cano, M.; Guedeney, G.; Richaud, P.; Cournac, L. Role of HoxE subunit in Synechocystis PCC6803 hydrogenase. FEBS J. 2011, 278, 4035–4043. [Google Scholar] [CrossRef]
- Appel, J.; Phunpruch, S.; Steinmuller, K.; Schulz, R. The bidirectional hydrogenase of Synechocystis sp PCC 6803 works as an electron valve during photosynthesis. Arch. Microbiol. 2000, 173, 333–338. [Google Scholar] [CrossRef]
- Burgstaller, H.; Wang, Y.; Caliebe, J.; Hueren, V.; Appel, J.; Boehm, M.; Leitzke, S.; Theune, M.; King, P.W.; Gutekunst, K. Synechocystis sp. PCC 6803 Requires the Bidirectional Hydrogenase to Metabolize Glucose and Arginine Under Oxic Conditions. Front. Microbiol. 2022, 13, 896190. [Google Scholar] [CrossRef]
- Ohkawa, H.; Pakrasi, H.B.; Ogawa, T. Two types of functionally distinct NAD(P)H dehydrogenases in Synechocystis sp. strain PCC6803. J. Biol. Chem. 2000, 275, 31630–31634. [Google Scholar] [CrossRef] [Green Version]
- Shibata, M.; Ohkawa, H.; Kaneko, T.; Fukuzawa, H.; Tabata, S.; Kaplan, A.; Ogawa, T. Distinct constitutive and low-CO2-induced CO2 uptake systems in cyanobacteria: Genes involved and their phylogenetic relationship with homologous genes in other organisms. Proc. Natl. Acad. Sci. USA 2001, 98, 11789–11794. [Google Scholar] [CrossRef] [Green Version]
- Burroughs, N.J.; Boehm, M.; Eckert, C.; Mastroianni, G.; Spence, E.M.; Yu, J.; Nixon, P.J.; Appel, J.; Mullineaux, C.W.; Bryan, S.J. Solar powered biohydrogen production requires specific localization of the hydrogenase. Energy Environ. Sci. 2014, 7, 3791–3800. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.-N.; Bryan, S.J.; Huang, F.; Yu, J.; Nixon, P.J.; Rich, P.R.; Mullineaux, C.W. Control of electron transport routes through redox-regulated redistribution of respiratory complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 11431–11436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, X.; Spengler, K.; Terberger, K.; Boehm, M.; Appel, J.; Barske, T.; Timm, S.; Battchikova, N.; Hagemann, M.; et al. Pyruvate:ferredoxin oxidoreductase and low abundant ferredoxins support aerobic photomixotrophic growth in cyanobacteria. eLife 2022, 11, e71339. [Google Scholar] [CrossRef] [PubMed]
- Makowka, A.; Nichelmann, L.; Schulze, D.; Spengler, K.; Wittmann, C.; Forchhammer, K.; Gutekunst, K. Glycolytic Shunts Replenish the Calvin-Benson-Bassham Cycle as Anaplerotic Reactions in Cyanobacteria. Mol. Plant 2020, 13, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343-U341. [Google Scholar] [CrossRef]
- Williams, J.G.K. [85] Construction of specific mutations in photosystem II photosynthetic reaction center by genetic engineering methods in Synechocystis 6803. In Methods Enzymol; Academic Press: Cambridge, MA, USA, 1988; Volume 167, pp. 766–778. [Google Scholar]
- Fatihi, A.; Latimer, S.; Schmollinger, S.; Block, A.; Dussault, P.H.; Vermaas, W.F.; Merchant, S.S.; Basset, G.J. A Dedicated Type II NADPH Dehydrogenase Performs the Penultimate Step in the Biosynthesis of Vitamin K1 in Synechocystis and Arabidopsis. Plant Cell 2015, 27, 1730–1741. [Google Scholar] [CrossRef] [Green Version]
- Theune, M.L.; Hildebrandt, S.; Steffen-Heins, A.; Bilger, W.; Gutekunst, K.; Appel, J. In-vivo quantification of electron flow through photosystem I—Cyclic electron transport makes up about 35% in a cyanobacterium. Biochim. Biophys. Acta (BBA)-Bioenerg. 2021, 1862, 148353. [Google Scholar] [CrossRef] [PubMed]
- Spence, E.; Sarcina, M.; Ray, N.; Moller, S.G.; Mullineaux, C.W.; Robinson, C. Membrane-specific targeting of green fluorescent protein by the Tat pathway in the cyanobacterium Synechocystis PCC6803. Mol. Microbiol. 2003, 48, 1481–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcdonald, J.H.; Dunn, K.W. Statistical tests for measures of colocalization in biological microscopy. J. Microsc. 2013, 252, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, N.; Ceccarelli, E.A. Open questions in ferredoxin-NADP+ reductase catalytic mechanism. Eur. J. Biochem. 2003, 270, 1900–1915. [Google Scholar] [CrossRef]
- Kramer, D.M.; Evans, J.R. The Importance of Energy Balance in Improving Photosynthetic Productivity. Plant Physiol. 2011, 155, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Knoop, H.; Steuer, R. A Computational Analysis of Stoichiometric Constraints and Trade-Offs in Cyanobacterial Biofuel Production. Front. Bioeng. Biotechnol. 2015, 3, 47. [Google Scholar] [CrossRef] [PubMed]
- Belkin, S.; Packer, L. Determination of pH gradients in intact cyanobacteria by electron spin resonance spectroscopy. Methods Enzymol. 1988, 167, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Cooley, J.W.; Vermaas, W.F.J. Succinate dehydrogenase and other respiratory pathways in thylakoid membranes of Synechocystis sp. strain PCC 6803: Capacity comparisons and physiological function. J. Bacteriol. 2001, 183, 4251–4258. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, C.W.; Emlyn-Jones, D. State transitions: An example of acclimation to low-light stress. J. Exp. Bot. 2004, 56, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Santana Sánchez, A.; Ermakova, M.; Rögner, M.; Cournac, L.; Allahverdiyeva, Y. Functional redundancy between flavodiiron proteins and NDH-1 in Synechocystis sp. PCC 6803. Plant J. 2020, 11, 14812. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.W.; Kamocka, M.M.; McDonald, J.H. A practical guide to evaluating colocalization in biological microscopy. Am. J. Physiol.-Cell Physiol. 2011, 300, C723–C742. [Google Scholar] [CrossRef] [Green Version]
- Herce, H.D.; Casas-Delucchi, C.S.; Cardoso, M.C. New image colocalization coefficient for fluorescence microscopy to quantify (bio-)molecular interactions. J. Microsc. 2013, 249, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, A.; Gaffron, H.; Battley, E.H. Photosynthesis and photoreduction by the blue green alga, synechococcus-elongatus, nag. Biol. Bull. 1950, 99, 157–162. [Google Scholar] [CrossRef]
- Houchins, J.P. The physiology and biochemistry of hydrogen metabolism in cyanobacteria. Biochim. Biophys. Acta 1984, 768, 227–255. [Google Scholar] [CrossRef]
- Howitt, C.A.; Udall, P.K.; Vermaas, W.F.J. Type 2 NADH dehydrogenases in the cyanobacterium Synechocystis sp strain PCC 6803 are involved in regulation rather than respiration. J. Bacteriol. 1999, 181, 3994–4003. [Google Scholar] [CrossRef] [Green Version]
- Schneider, K.; Schlegel, H.G. Purification and properties of soluble hydrogenase from Alcaligenes eutrophus H 16. Biochim. Biophys. Acta 1976, 452, 66–80. [Google Scholar] [CrossRef]
- Lupacchini, S.; Appel, J.; Stauder, R.; Bolay, P.; Klähn, S.; Lettau, E.; Adrian, L.; Lauterbach, L.; Bühler, B.; Schmid, A.; et al. Rewiring cyanobacterial photosynthesis by the implementation of an oxygen-tolerant hydrogenase. Metab. Eng. 2021, 68, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Schulze, D.; Kohlstedt, M.; Becker, J.; Cahoreau, E.; Peyriga, L.; Makowka, A.; Hildebrandt, S.; Gutekunst, K.; Portais, J.-C.; Wittmann, C. GC/MS-based 13C metabolic flux analysis resolves the parallel and cyclic photomixotrophic metabolism of Synechocystis sp. PCC 6803 and selected deletion mutants including the Entner-Doudoroff and phosphoketolase pathways. Microb. Cell Factories 2022, 21, 69. [Google Scholar] [CrossRef] [PubMed]
- Sukkasam, N.; Incharoensakdi, A.; Monshupanee, T. Disruption of Hydrogen Gas Synthesis Enhances the Cellular Levels of NAD(P)H, Glycogen, Poly(3-hydroxybutyrate) and Photosynthetic Pigments Under Specific Nutrient Condition(s) in Cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2021, 63, 135–147. [Google Scholar] [CrossRef]
- Hedderich, R.; Forzi, L. Energy-converting NiFe hydrogenases: More than just H-2 activation. J. Mol. Microbiol. Biotechnol. 2005, 10, 92–104. [Google Scholar] [CrossRef]
- Gutekunst, K. Hypothesis on the Synchronistic Evolution of Autotrophy and Heterotrophy. Trends Biochem. Sci. 2018, 43, 402–411. [Google Scholar] [CrossRef]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Evolution of Photosynthesis. Annu. Rev. Plant Biol. 2011, 62, 515–548. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Vmax + H2 e−/PSI/s | Vmax H2-Uptake e−/PSI/s | ATP/NADPH Ratio 1 |
|---|---|---|---|
| WT | 12.4 ± 0.5 | 6.0 ± 0.9 | 3.5 ± 0.5 |
| ΔndhD1 | 10.2 ± 1.1 | 5.4 ± 0.9 | 3.3 ± 0.7 |
| ΔndhD2 | 10.0 ± 0.4 | 6.4 ± 0.8 | 2.7 ± 0.4 |
| ΔndhD1ΔndhD2 | 9.2 ± 1.6 | 1.9 ± 0.5 | 4.0 ± 1.2 2 |
| ΔndhD3ΔndhD4 | 15.8 ± 1.7 | 6.4 ± 0.9 | 4.2 ± 0.8 |
| Δndh-2 | 20.6 ± 0.7 | 14.1 ± 1.4 | 2.6 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Appel, J.; Craig, S.; Theune, M.; Hüren, V.; Künzel, S.; Forberich, B.; Bryan, S.; Gutekunst, K. Evidence for Electron Transfer from the Bidirectional Hydrogenase to the Photosynthetic Complex I (NDH-1) in the Cyanobacterium Synechocystis sp. PCC 6803. Microorganisms 2022, 10, 1617. https://doi.org/10.3390/microorganisms10081617
Appel J, Craig S, Theune M, Hüren V, Künzel S, Forberich B, Bryan S, Gutekunst K. Evidence for Electron Transfer from the Bidirectional Hydrogenase to the Photosynthetic Complex I (NDH-1) in the Cyanobacterium Synechocystis sp. PCC 6803. Microorganisms. 2022; 10(8):1617. https://doi.org/10.3390/microorganisms10081617
Chicago/Turabian StyleAppel, Jens, Sean Craig, Marius Theune, Vanessa Hüren, Sven Künzel, Björn Forberich, Samantha Bryan, and Kirstin Gutekunst. 2022. "Evidence for Electron Transfer from the Bidirectional Hydrogenase to the Photosynthetic Complex I (NDH-1) in the Cyanobacterium Synechocystis sp. PCC 6803" Microorganisms 10, no. 8: 1617. https://doi.org/10.3390/microorganisms10081617