
Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Environmental Parameters
2.2. Primary Productivity Assay
2.3. Cultivation and Enumeration of Targeted Populations
2.4. Microbial Community eDNA Sequencing
3. Results
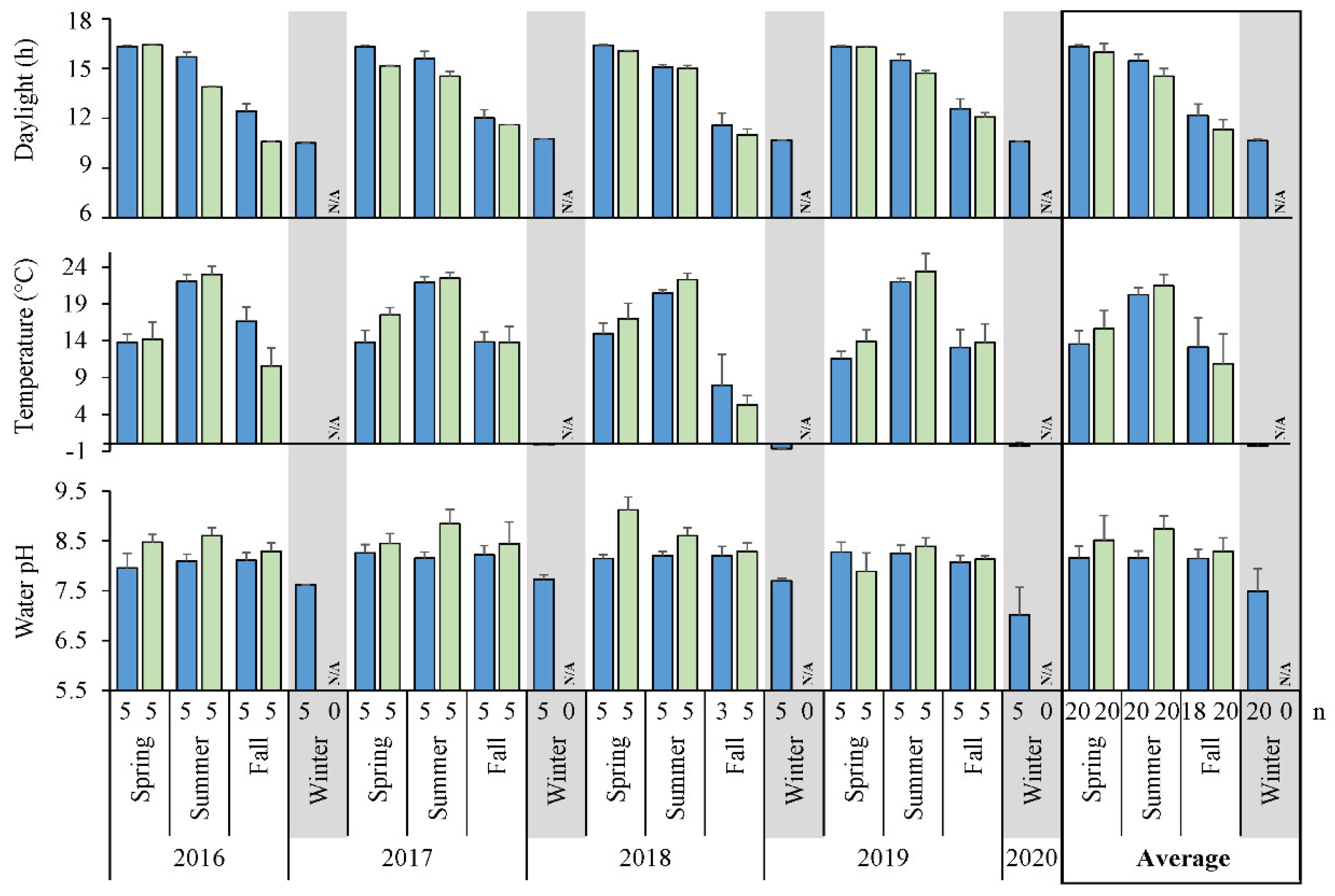
3.1. Environmental Characteristics
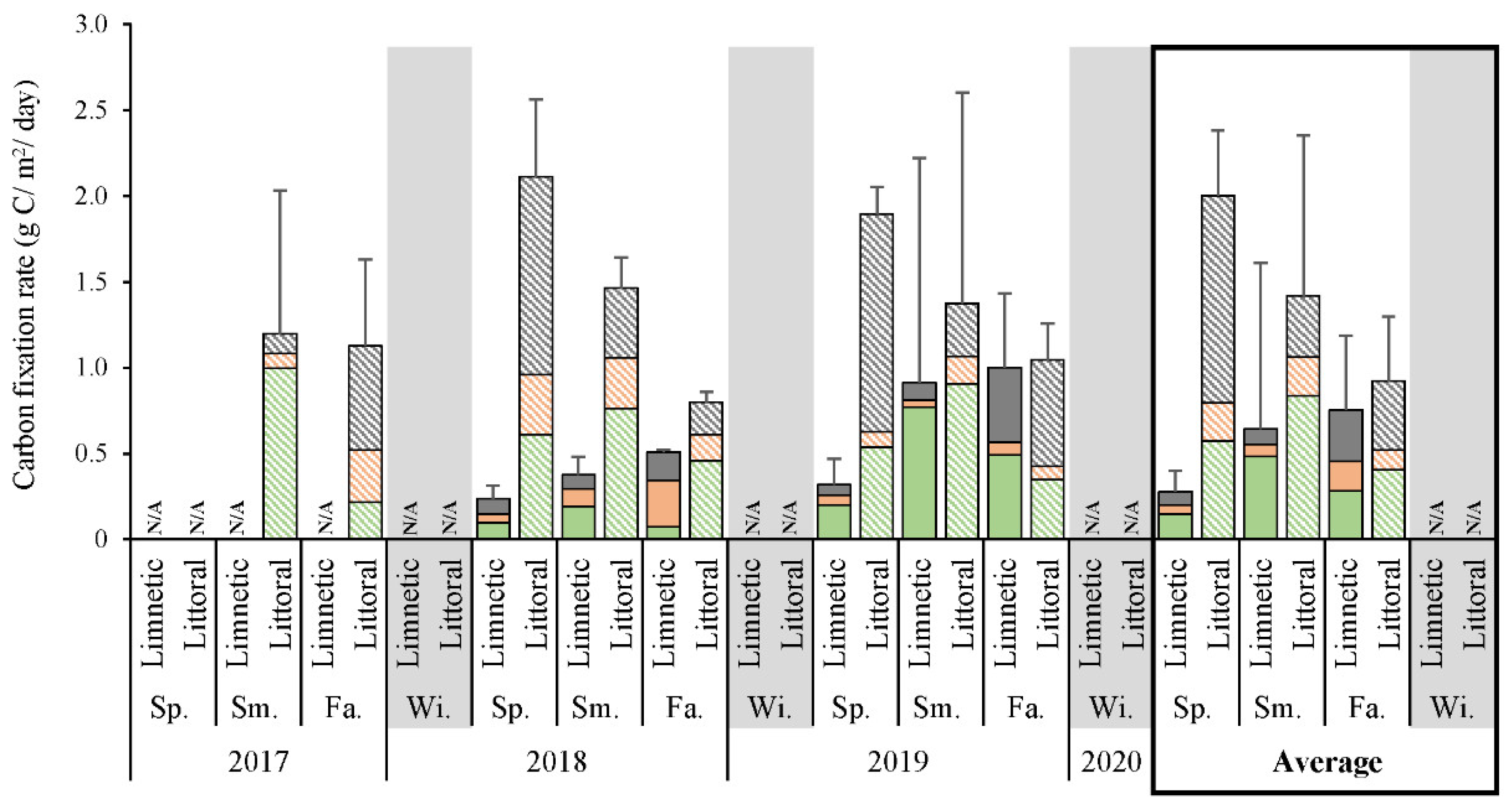
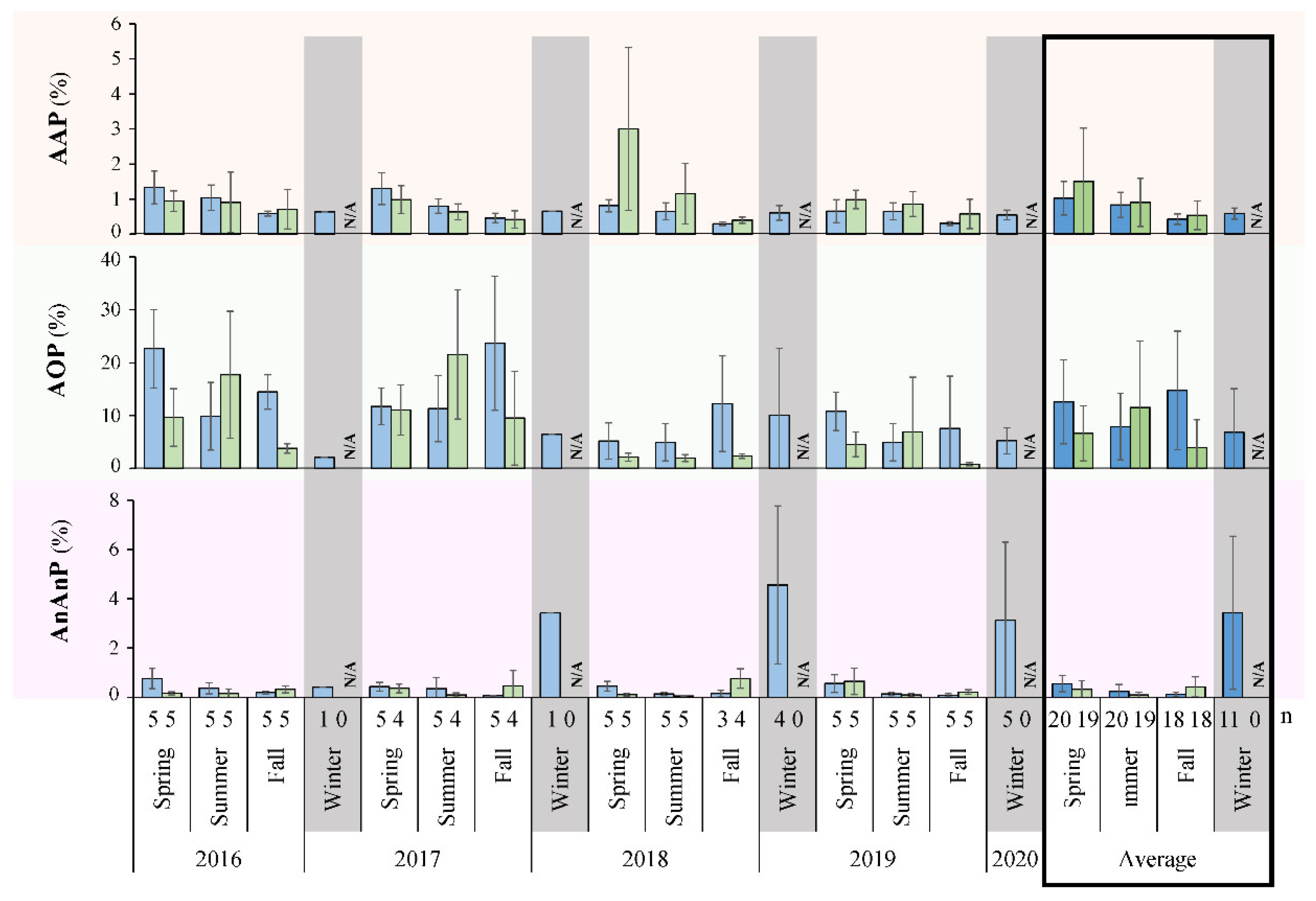
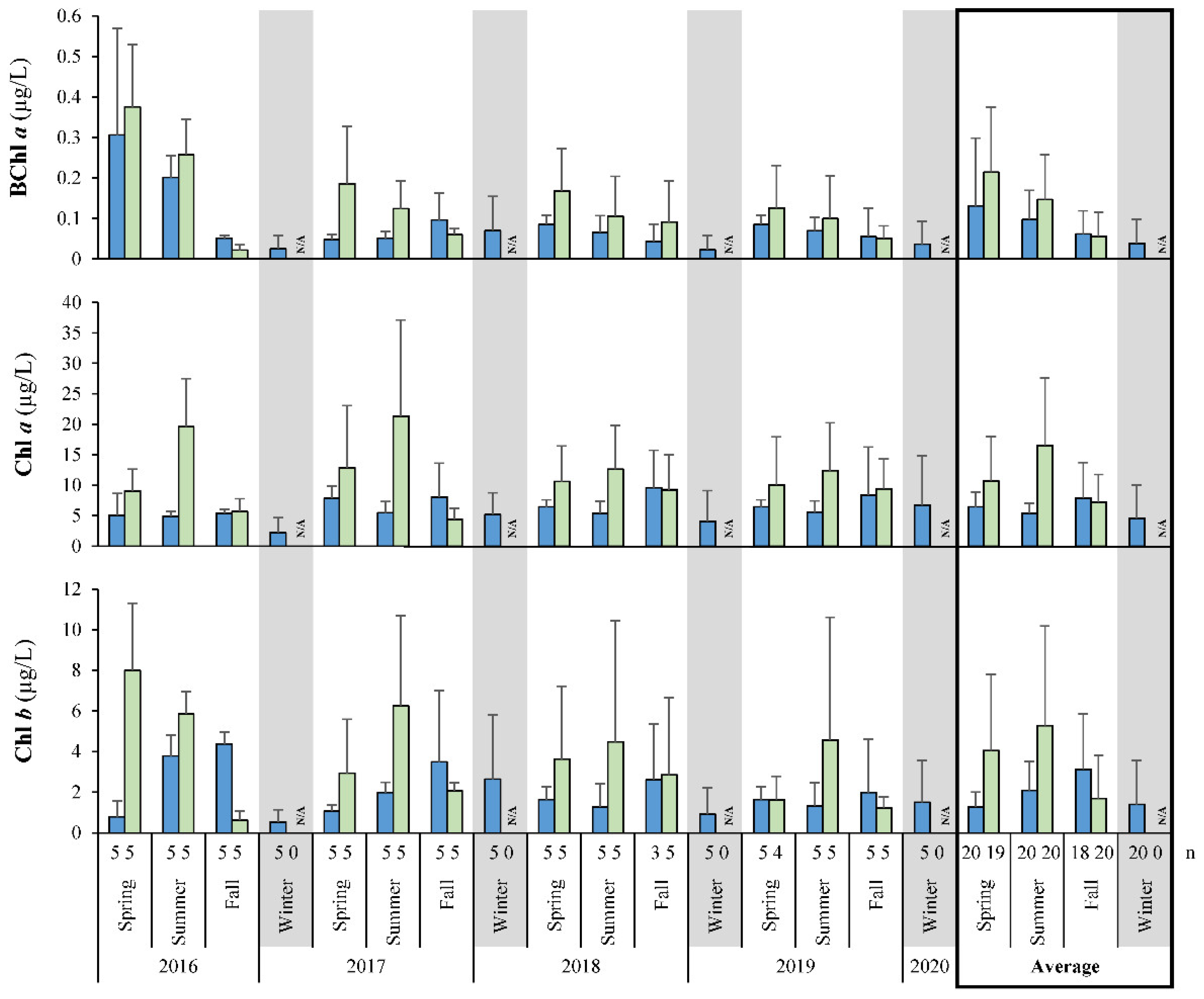
3.2. Primary Productivity Rates
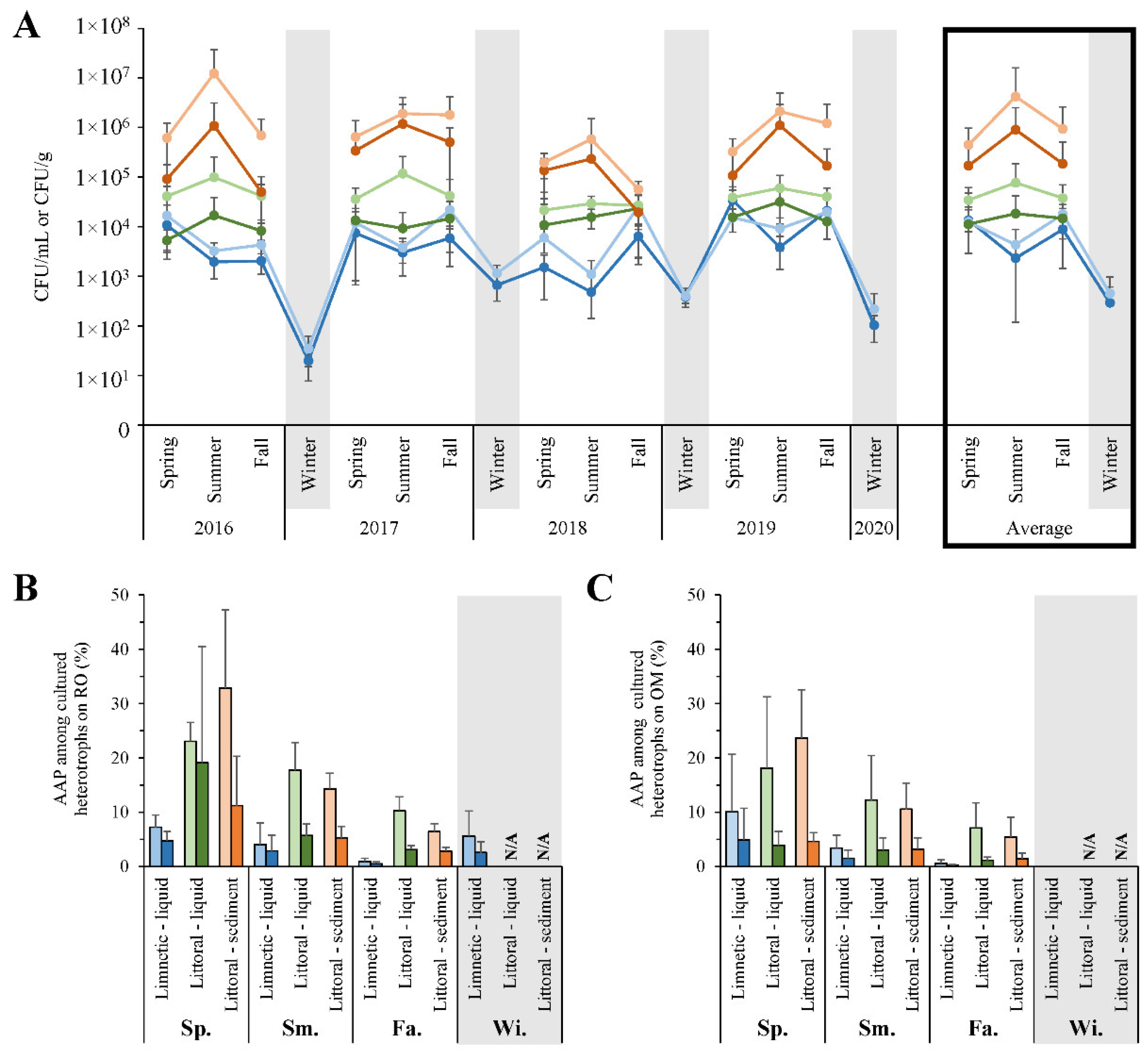
3.3. Seasonal Fluctuations in Cultivated Microbial Counts
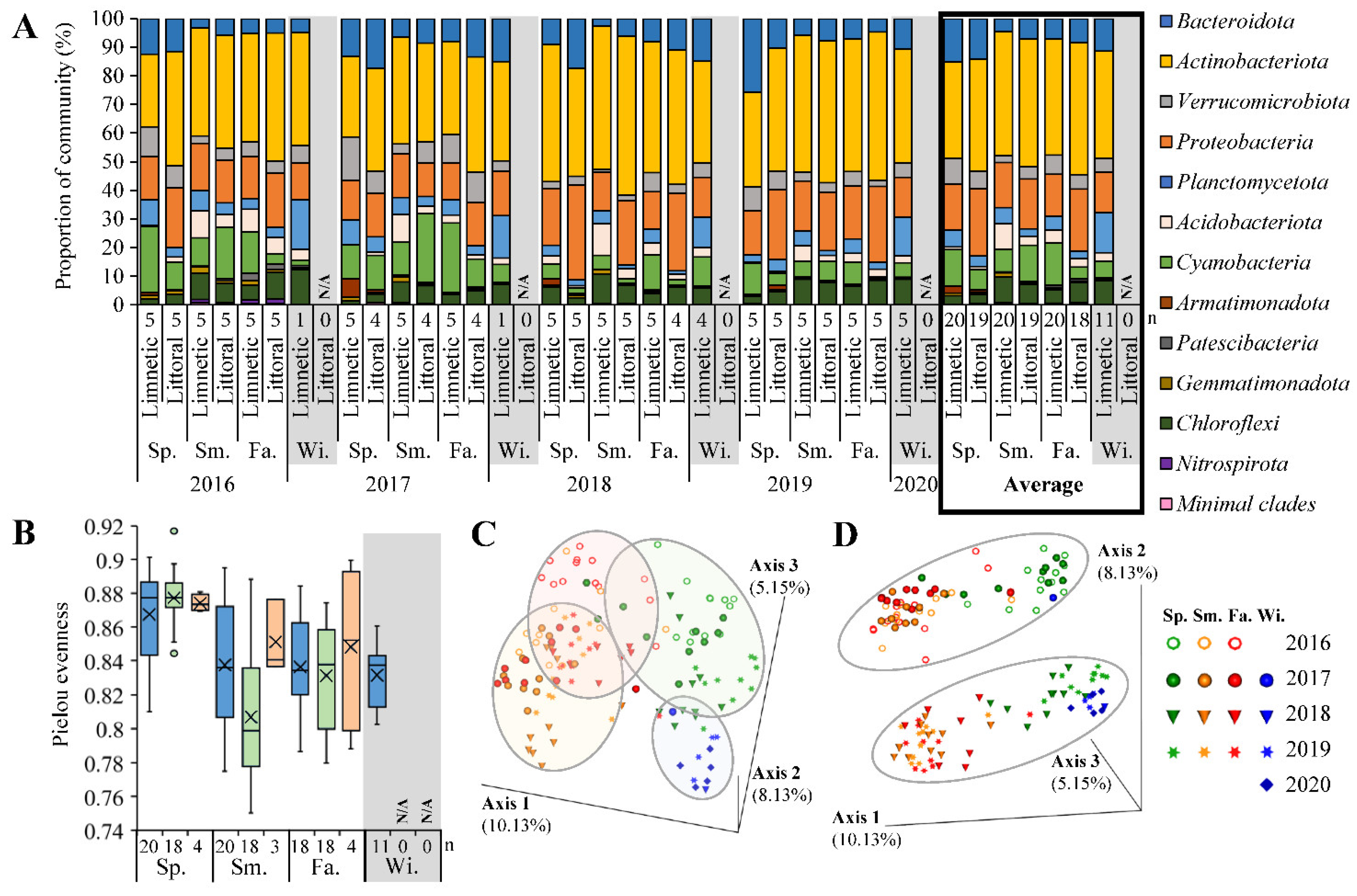
3.4. Sequenced Microbial Communities
4. Discussion
4.1. Habitat Features
4.2. Primary Productivity in Lake Winnipeg
4.3. Trends in Cultivated Microbial Numbers
4.4. Sequenced Community Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Reed, A.J.; Hicks, R.E. Microbial ecology of Lake Superior Bacteria and Archaea: An overview. Aquat. Ecosyst. Health Manag. 2011, 14, 386–395. [Google Scholar] [CrossRef]
- Mou, X.; Jacob, J.; Lu, X.; Robbins, S.; Sun, S.; Ortiz, J.D. Diversity and distribution of free-living and particle-associated bacterioplankton in Sandusky Bay and adjacent waters of Lake Erie Western Basin. J. Great Lakes Res. 2013, 39, 352–357. [Google Scholar] [CrossRef]
- Nakatsu, C.H.; Byappanahalli, M.N.; Nevers, M.B. Bacterial Community 16S rRNA Gene Sequencing Characterizes Riverine Microbial Impact on Lake Michigan. Front. Microbiol. 2019, 10, 996. [Google Scholar] [CrossRef]
- Paver, S.F.; Newton, R.J.; Coleman, M.L. Microbial communities of the Laurentian Great Lakes reflect connectivity and local biogeochemistry. Environ. Microbiol. 2020, 22, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Evans, M. The large lake ecosystems of northern Canada. Aquat. Ecosyst. Health Manag. 2000, 3, 65–79. [Google Scholar] [CrossRef]
- Boguski, D.A. Annotated Bibliography of Aquatic Research in the Great Slave Lake Region. Biodive Sci. Manuscr. Rep. 2015, 1502, 1–79. [Google Scholar]
- Koehler, G.; Brua, R.B.; Enders, E.C. Foreword to the second Lake Winnipeg special issue. J. Great Lakes Res. 2021, 47, 553–555. [Google Scholar] [CrossRef]
- Matisoff, G.; Watson, S.B.; Guo, J.; Duewiger, A.; Steely, R. Sediment and nutrient distribution and resuspension in Lake Winnipeg. Sci. Total Environ. 2017, 575, 173–186. [Google Scholar] [CrossRef]
- (MWSD) Manitoba Water Stewardship Division. State of Lake Winnipeg: 1999 to 2007. Manit. Steward. Board Environ. Can.; 2011; pp. 1–222. Available online: https://publications.gc.ca/site/eng/407829/publication.html (accessed on 18 August 2022).
- (MARD) Manitoba Agriculture and Resource Department. State of Lake Winnipeg 2nd edition. Environ. Clim. Chang. Can.; 2020; pp. 1–195. Available online: https://www.gov.mb.ca/water/pubs/water/lakes-beaches-rivers/state_lake_wpg_report_tech.pdf (accessed on 18 August 2022).
- Kling, H.J.; Watson, S.B.; McCullough, G.K.; Stainton, M.P. Bloom development and phytoplankton succession in Lake Winnipeg: A comparison of historical records with recent data. Aquat. Ecosyst. Health Manag. 2011, 14, 219–224. [Google Scholar] [CrossRef]
- Pick, F.R. Blooming algae: A Canadian perspective on the rise of toxic cyanobacteria. Can. J. Fish. Aquat. Sci. 2016, 73, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzyk, S.B.; Pritchard, A.O.; Plouffe, J.; Sorensen, J.L.; Yurkov, V. Psychrotrophic violacein-producing bacteria isolated from Lake Winnipeg, Canada. J. Great Lakes Res. 2021, 47, 715–724. [Google Scholar] [CrossRef]
- Kuzyk, S.B.; Wiens, K.; Ma, X.; Yurkov, V. Association of aerobic anoxygenic phototrophs and zebra mussels, Dreissena polymorpha, within the littoral zone of Lake Winnipeg. J. Great Lakes Res. 2021, 47, 567–582. [Google Scholar] [CrossRef]
- Vila, X.; Abella, C.A. Effects of light quality on the physiology and the ecology of planktonic green sulfur bacteria in lakes. Photosynth. Res. 1994, 41, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, V.; Beatty, J.T. Aerobic Anoxygenic Phototrophic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 695–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurkov, V.; Hughes, E. Aerobic Anoxygenic Phototrophs: Four Decades of Mystery. In Modern Topics in the Phototrophic Prokaryotes; Springer International Publishing: Cham, Switzerland, 2017; pp. 193–214. [Google Scholar]
- Vila, X.; Abella, C.; Figueras, J.; Hurley, J. Vertical models of phototrophic bacterial distribution in the metalimnetic microbial communities of several freshwater North-American kettle lakes. FEMS Microbiol. Ecol. 1998, 25, 287–299. [Google Scholar] [CrossRef]
- Koblížek, M. Ecology of aerobic anoxygenic phototrophs in aquatic environments. FEMS Microbiol. Rev. 2015, 39, 854–870. [Google Scholar] [CrossRef] [Green Version]
- Bilyj, M.; Lepitzki, D.; Hughes, E.; Swiderski, J.; Stackebrandt, E.; Pacas, C.; Yurkov, V. Abundance and Diversity of the Phototrophic Microbial Mat Communities of Sulphur Mountain Banff Springs and Their Significance to the Endangered Snail, Physella johnsoni. Open J. Ecol. 2014, 4, 488–516. [Google Scholar] [CrossRef] [Green Version]
- Fauteux, L.; Cottrell, M.T.; Kirchman, D.L.; Borrego, C.M.; Garcia-Chaves, M.C.; del Giorgio, P.A. Patterns in Abundance, Cell Size and Pigment Content of Aerobic Anoxygenic Phototrophic Bacteria along Environmental Gradients in Northern Lakes. PLoS ONE 2015, 10, e0124035. [Google Scholar] [CrossRef]
- Depew, D.C.; Krutzelmann, E.; Watchorn, K.E.; Caskenette, A.; Enders, E.C. The distribution, density, and biomass of the zebra mussel (Dreissena polymorpha) on natural substrates in Lake Winnipeg 2017–2019. J. Great Lakes Res. 2021, 47, 556–566. [Google Scholar] [CrossRef]
- Ritchie, R.J. Measurement of chlorophylls a and b and bacteriochlorophyll a in organisms from hypereutrophic auxinic waters. J. Appl. Phycol. 2018, 30, 3075–3087. [Google Scholar] [CrossRef]
- Gregersen, L.H.; Habicht, K.S.; Peduzzi, S.; Tonolla, M.; Canfield, D.E.; Miller, M.; Cox, R.P.; Frigaard, N.-U. Dominance of a clonal green sulfur bacterial population in a stratified lake. FEMS Microbiol. Ecol. 2009, 70, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SCOR-UNESCO. Determination of photosynthetic pigments. Monogr. Oceanogr. Methodol. 1966, 1, 11–18. [Google Scholar]
- Steemann-Nielsen, E. The Use of Radioactive Carbon (14C) for Measuring Organic Production in the Sea. ICES J. Mar. Sci. 1952, 43, 117–140. [Google Scholar] [CrossRef]
- Franson, M. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Yurkov, V.; Van Gemerden, H. Abundance and salt tolerance of obligately aerobic, phototrophic bacteria in a marine microbial mat. Neth. J. Sea Res. 1993, 31, 57–62. [Google Scholar] [CrossRef]
- Béjà, O.; Suzuki, M.T.; Heidelberg, J.F.; Nelson, W.C.; Preston, C.M.; Hamada, T.; Eisen, J.A.; Fraser, C.M.; DeLong, E.F. Unsuspected diversity among marine aerobic anoxygenic phototrophs. Nature 2002, 415, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Zervas, A.; Zeng, Y.; Madsen, A.M.; Hansen, L.H. Genomics of Aerobic Photoheterotrophs in Wheat Phyllosphere Reveals Divergent Evolutionary Patterns of Photosynthetic Genes in Methylobacterium spp. Genome Biol. Evol. 2019, 11, 2895–2908. [Google Scholar] [CrossRef]
- Kopejtka, K.; Zeng, Y.; Kaftan, D.; Selyanin, V.; Gardian, Z.; Tomasch, J.; Sommaruga, R.; Koblížek, M. Characterization of the Aerobic Anoxygenic Phototrophic Bacterium Sphingomonas sp. AAP5. Microorganisms 2021, 9, 768. [Google Scholar] [CrossRef]
- Hughes, E.; Head, B.; Maltman, C.; Piercey-Normore, M.; Yurkov, V. Aerobic anoxygenic phototrophs in gold mine tailings in Nopiming Provincial Park, Manitoba, Canada. Can. J. Microbiol. 2017, 63, 212–218. [Google Scholar] [CrossRef]
- Maltman, C.; Yurkov, V. The Effect of Tellurite on Highly Resistant Freshwater Aerobic Anoxygenic Phototrophs and Their Strategies for Reduction. Microorganisms 2015, 3, 826–838. [Google Scholar] [CrossRef] [Green Version]
- McKinney, W. Data Structures for Statistical Computing in Python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; pp. 56–61. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- McDonald, D.; Clemente, J.C.; Kuczynski, J.; Rideout, J.R.; Stombaugh, J.; Wendel, D.; Wilke, A.; Huse, S.; Hufnagle, J.; Meyer, F.; et al. The Biological Observation Matrix (BIOM) format or: How I learned to stop worrying and love the ome-ome. Gigascience 2012, 1, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. MSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E.; Goodfellow, M. Nucleic Acid Techniques in Bacterial Systematics; Wiley: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Gonzalez, A.; Smarr, L.; McDonald, D.; Morton, J.T.; Navas-Molina, J.A.; Knight, R. Bringing the Dynamic Microbiome to Life with Animations. Cell Host Microbe 2017, 21, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenaar, L.I. Dissolved oxygen status of Lake Winnipeg: Spatio-temporal and isotopic (δ18O–O2) patterns. J. Great Lakes Res. 2012, 38, 123–134. [Google Scholar] [CrossRef]
- Binding, C.E.; Greenberg, T.A.; McCullough, G.; Watson, S.B.; Page, E. An analysis of satellite-derived chlorophyll and algal bloom indices on Lake Winnipeg. J. Great Lakes Res. 2018, 44, 436–446. [Google Scholar] [CrossRef]
- Mašín, M.; Nedoma, J.; Pechar, L.; Koblížek, M. Distribution of aerobic anoxygenic phototrophs in temperate freshwater systems. Environ. Microbiol. 2008, 10, 1988–1996. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N. Z. J. Mar. Freshw. Res. 1987, 21, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Sayers, M.; Bosse, K.; Fahnenstiel, G.; Shuchman, R. Carbon Fixation Trends in Eleven of the World’s Largest Lakes: 2003–2018. Water 2020, 12, 3500. [Google Scholar] [CrossRef]
- Yezhova, Y.; Capelle, D.; Stainton, M.; Papakyriakou, T. Carbon fixation by the phytoplankton community across Lake Winnipeg. J. Great Lakes Res. 2021, 47, 703–714. [Google Scholar] [CrossRef]
- Vollenweider, R.A.; Munawar, M.; Stadelmann, P. A Comparative Review of Phytoplankton and Primary Production in the Laurentian Great Lakes. J. Fish. Res. Board Can. 1974, 31, 739–762. [Google Scholar] [CrossRef]
- Frischer, M.E.; Nierzwicki-Bauer, S.A.; Parsons, R.H.; Vathanodorn, K.; Waitkus, K.R. Interactions between zebra mussels (Dreissena polymorpha) and microbial communities. Can. J. Fish. Aquat. Sci. 2000, 57, 591–599. [Google Scholar] [CrossRef]
- Yurkov, V.; Jappe, J.; Vermeglio, A. Tellurite resistance and reduction by obligately aerobic photosynthetic bacteria. Appl. Environ. Microbiol. 1996, 62, 4195–4198. [Google Scholar] [CrossRef] [Green Version]
- Jankowiak, J.; Hattenrath-Lehmann, T.; Kramer, B.J.; Ladds, M.; Gobler, C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensification, diversity, and toxicity in western Lake Erie. Limnol. Oceanogr. 2019, 64, 1347–1370. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Stegen, J.C.; Yu, Y.; Deng, Y.; Li, X.; Wu, S.; Dai, L.; Zhang, X.; Li, J.; Wang, C.; et al. Nearly a decade-long repeatable seasonal diversity patterns of bacterioplankton communities in the eutrophic Lake Donghu (Wuhan, China). Mol. Ecol. 2017, 26, 3839–3850. [Google Scholar] [CrossRef] [PubMed]
- Kolářová, E.; Medová, H.; Piwosz, K.; Koblížek, M. Seasonal dynamics of aerobic anoxygenic phototrophs in freshwater lake Vlkov. Folia Microbiol. 2019, 64, 705–710. [Google Scholar] [CrossRef]
- Čuperová, Z.; Holzer, E.; Salka, I.; Sommaruga, R.; Koblížek, M. Temporal Changes and Altitudinal Distribution of Aerobic Anoxygenic Phototrophs in Mountain Lakes. Appl. Environ. Microbiol. 2013, 79, 6439–6446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fecskeová, L.K.; Piwosz, K.; Hanusová, M.; Nedoma, J.; Znachor, P.; Koblížek, M. Diel changes and diversity of pufM expression in freshwater communities of anoxygenic phototrophic bacteria. Sci. Rep. 2019, 9, 18766. [Google Scholar] [CrossRef] [PubMed]
- Csotonyi, J.T.; Swiderski, J.; Stackebrandt, E.; Yurkov, V. Novel halophilic aerobic anoxygenic phototrophs from a Canadian hypersaline spring system. Extremophiles 2008, 12, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Kasalický, V.; Zeng, Y.; Piwosz, K.; Šimek, K.; Kratochvilová, H.; Koblížek, M. Aerobic Anoxygenic Photosynthesis Is Commonly Present within the Genus Limnohabitans. Appl. Environ. Microbiol. 2018, 84, e02116-17. [Google Scholar] [CrossRef] [Green Version]
- Suyama, T.; Shigematsu, T.; Takaichi, S.; Nodasaka, Y.; Fujikawa, S.; Hosoya, H.; Tokiwa, Y.; Kanagawa, T.; Hanada, S. Roseateles depolymerans gen. nov., sp. nov., a new bacteriochlorophyll a-containing obligate aerobe belonging to the β-subclass of the Proteobacteria. Int. J. Syst. Evol. Microbiol. 1999, 49, 449–457. [Google Scholar] [CrossRef]
- Shiba, T. Roseobacter litoralis gen. nov., sp. nov., and Roseobacter denitrificans sp. nov., Aerobic Pink-Pigmented Bacteria Which Contain Bacteriochlorophyll a. Syst. Appl. Microbiol. 1991, 14, 140–145. [Google Scholar]
- Rathgeber, C.; Yurkova, N.; Stackebrandt, E.; Schumann, P.; Beatty, J.T.; Yurkov, V. Roseicyclus mahoneyensis gen. nov., sp. nov., an aerobic phototrophic bacterium isolated from a meromictic lake. Int. J. Syst. Evol. Microbiol. 2005, 55, 1597–1603. [Google Scholar] [CrossRef] [Green Version]
- Csotonyi, J.T.; Stackebrandt, E.; Swiderski, J.; Schumann, P.; Yurkov, V. An alphaproteobacterium capable of both aerobic and anaerobic anoxygenic photosynthesis but incapable of photoautotrophy: Charonomicrobium ambiphototrophicum, gen. nov., sp. nov. Photosynth. Res. 2011, 107, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Kuzyk, S.B.; Jafri, M.; Humphrey, E.; Maltman, C.; Kyndt, J.A.; Yurkov, V. Prosthecate aerobic anoxygenic phototrophs Photocaulis sulfatitolerans gen. nov. sp. nov. and Photocaulis rubescens sp. nov. isolated from alpine meromictic lakes in British Columbia, Canada. Arch. Microbiol. 2022, 204, 444. [Google Scholar] [CrossRef] [PubMed]
- Tank, M.; Bryant, D.A. Nutrient requirements and growth physiology of the photoheterotrophic Acidobacterium, Chloracidobacterium thermophilum. Front. Microbiol. 2015, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathgeber, C.; Beatty, J.T.; Yurkov, V. Aerobic phototrophic bacteria: New evidence for the diversity, ecological importance and applied potential of this previously overlooked group. Review. Photosynth. Res. 2004, 81, 113–128. [Google Scholar] [CrossRef]
- Yurkov, V.; Csotonyi, J. New light on aerobic anoxygenic phototrophs. Book chapter. In The Purple Photosynthetic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 31–55. [Google Scholar]
- Yurkov, V.; Hughes, E. Genes associated with the peculiar phenotypes of the aerobic anoxygenic phototrophs. In Genome Evolution of Photosynthetic Bacteria; Beatty, J.T., Jacquot, J.P., Gadal, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 66, pp. 327–358. [Google Scholar]
- Peyraud, R.; Schneider, K.; Kiefer, P.; Massou, S.; Vorholt, J.A.; Portais, J.C. Genome-scale reconstruction and system level investigation of the metabolic network of Methylobacterium extorquens AM1. BMC Syst. Biol. 2011, 5, 189. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.; Yang, L.H.; Chen, Y.P.; Tao, Y.; Feng, F.Y.; Meng, J.Y. Aerophototrophica crusticola gen. nov., sp. nov., isolated from desert biocrusts. Int. J. Syst. Evol. Microbiol. 2021, 71, 004677. [Google Scholar] [CrossRef]
- Mehrshad, M.; Salcher, M.M.; Okazaki, Y.; Nakano, S.-i.; Šimek, K.; Andrei, A.-S.; Ghai, R. Hidden in plain sight—highly abundant and diverse planktonic freshwater Chloroflexi. Microbiome 2018, 6, 176. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Kyndt, J.A.; Meyer, T.E. Genomic Comparison, Phylogeny and Taxonomic Reevaluation of the Ectothiorhodospiraceae and Description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov. Microorganisms 2022, 10, 295. [Google Scholar] [CrossRef]
- Kyndt, J.A.; Salama, D.M.; Meyer, T.E.; Imhoff, J.F. Phylogenetic relationship of phototrophic heliobacteria and systematic reconsideration of species and genus assignments based on genome sequences of eight species. Int. J. Syst. Evol. Microbiol. 2021, 71, 004729. [Google Scholar] [CrossRef]
- Imhoff, J.F. Phylogenetic taxonomy of the family Chlorobiaceae on the basis of 16S rRNA and fmo (Fenna– Matthews–Olson protein) gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 941–951. [Google Scholar] [CrossRef] [Green Version]
- Saini, M.K.; ChihChe, W.; Soulier, N.; Sebastian, A.; Albert, I.; Thiel, V.; Bryant, D.A.; Hanada, S.; Tank, M. Caldichromatium japonicum gen. nov., sp. nov., a novel thermophilic phototrophic purple sulphur bacterium of the Chromatiaceae isolated from Nakabusa hot springs, Japan. Int. J. Syst. Evol. Microbiol. 2020, 70, 5701–5710. [Google Scholar] [CrossRef] [PubMed]
- Nupur, N.; Saini, M.K.; Singh, P.K.; Korpole, S.; Tanuku, N.R.S.; Takaichi, S.; Pinnaka, A.K. Imhoffiella gen. nov., a marine phototrophic member of the family Chromatiaceae including the description of Imhoffiella purpurea sp. nov. and the reclassification of Thiorhodococcus bheemlicus Anil Kumar et al. 2007 as Imhoffiella bheemlica comb. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Suling, J.; Petri, R. Phylogenetic relationships among the Chromatiaceae, their taxonomic reclassification and description of the new genera Allochromatium, Halochromatium, Isochromatium, Marichromatium, Thiococcus, Thiohalocapsa and Thermochromatium. Int. J. Syst. Bacteriol. 1998, 48, 1129–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlenko, V.M.; Krassilnikova, E.N.; Kikina, O.G.; Tatarinova, N.Y. The new motile purple sulfur bacteria Lamprobacter modestohalophilus nov. gen., nov. sp. with gas vacuoles. Izv. Akad. Nauk SSSR Ser. Biol. 1979, 5, 755–767. [Google Scholar]
- Watson, S.W. Characteristics of a marine nitrifying bacterium, Nitrosocystis oceanus sp. n. Limnol. Oceanogr. 1965, 10, R274–R289. [Google Scholar] [CrossRef]
- Tanuku, N.R.; Shinichi, T.; Pinnaka, A.K. Phaeobacterium nitratireducens gen. nov., sp. nov., a phototrophic gammaproteobacterium isolated from a mangrove forest sediment sample. Int. J. Syst. Evol. Microbiol. 2015, 65, 2357–2364. [Google Scholar]
- Shivali, K.; Sasikala, C.; Ramana, C.V. MLSA barcoding of Marichromatium spp. and reclassification of Marichromatium fluminis (Sucharita et al., 2010) as Phaeochromatium fluminis gen. nov. comb. nov. Syst. Appl. Microbiol. 2012, 35, 221–225. [Google Scholar] [CrossRef]
- Dilling, W.; Liesack, W.; Pfennig, N. Rhabdochromatium marinum gen. nom. rev., sp. nov., a purple sulfur bacterium from a salt marsh microbial mat. Arch. Microbiol. 1995, 164, 125–131. [Google Scholar] [CrossRef]
- Bryantseva, I.A.; Gorlenko, V.M.; Kompantseva, E.I.; Imhoff, J.F. Thioalkalicoccus limnaeus gen. nov., sp. nov., a new alkaliphilic purple sulfur bacterium with bacteriochlorophyll b. Int. J. Syst. Evol. Microbiol. 2000, 50, 2157–2163. [Google Scholar] [CrossRef] [Green Version]
- Rees, G.N.; Harfoot, C.G.; Janssen, P.H.; Schoenborn, L.; Kuever, J.; Lunsdorf, H. Thiobaca trueperi gen. nov., sp. nov., a phototrophic purple sulfur bacterium isolated from freshwater lake sediment. Int. J. Syst. Evol. Microbiol. 2002, 52, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Luedin, S.M.; Pothier, J.F.; Danza, F.; Storelli, N.; Frigaard, N.-U.; Wittwer, M.; Tonolla, M. Complete genome sequence of “Thiodictyon syntrophicum” sp. nov. strain Cad16T, a photolithoautotrophic purple sulfur bacterium isolated from the alpine meromictic Lake Cadagno. Genomic Sci. 2018, 13, 14. [Google Scholar] [CrossRef] [Green Version]
- Eichler, B.; Pfennig, N. Isolation and characteristics of Thiopedia rosea (neotype)*. Arch. Microbiol. 1991, 155, 210–216. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Meyer, T.E.; Kyndt, J.A. The genome sequence of the giant phototrophic gammaproteobacterium Thiospirillum jenense gives insight into its physiological properties and phylogenetic relationships. Arch. Microbiol. 2021, 203, 97–105. [Google Scholar] [CrossRef] [PubMed]
- van Niel, C.B. The culture, general physiology, morphology, and classification of the non-sulfur purple and brown bacteria. Bacteriol. Rev. 1944, 8, 1–118. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuzyk, S.B.; Ma, X.; Yurkov, V. Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability. Microorganisms 2022, 10, 1690. https://doi.org/10.3390/microorganisms10091690
Kuzyk SB, Ma X, Yurkov V. Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability. Microorganisms. 2022; 10(9):1690. https://doi.org/10.3390/microorganisms10091690
Chicago/Turabian StyleKuzyk, Steven B., Xiao Ma, and Vladimir Yurkov. 2022. "Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability" Microorganisms 10, no. 9: 1690. https://doi.org/10.3390/microorganisms10091690