
Translational Frameshifting in the chlD Gene Gives a Clue to the Coevolution of the Chlorophyll and Cobalamin Biosyntheses



Abstract
:1. Introduction
2. Materials and Methods
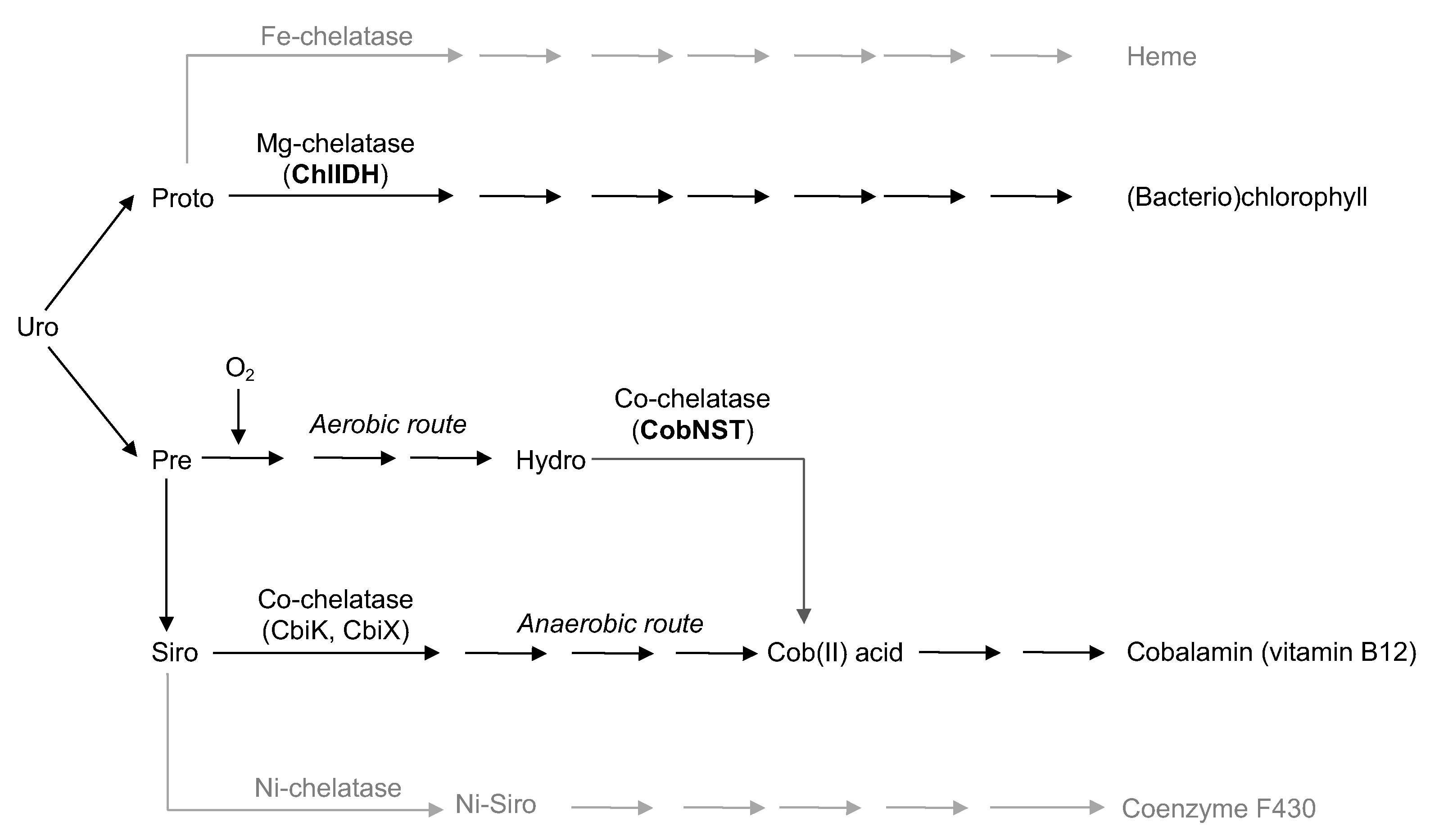
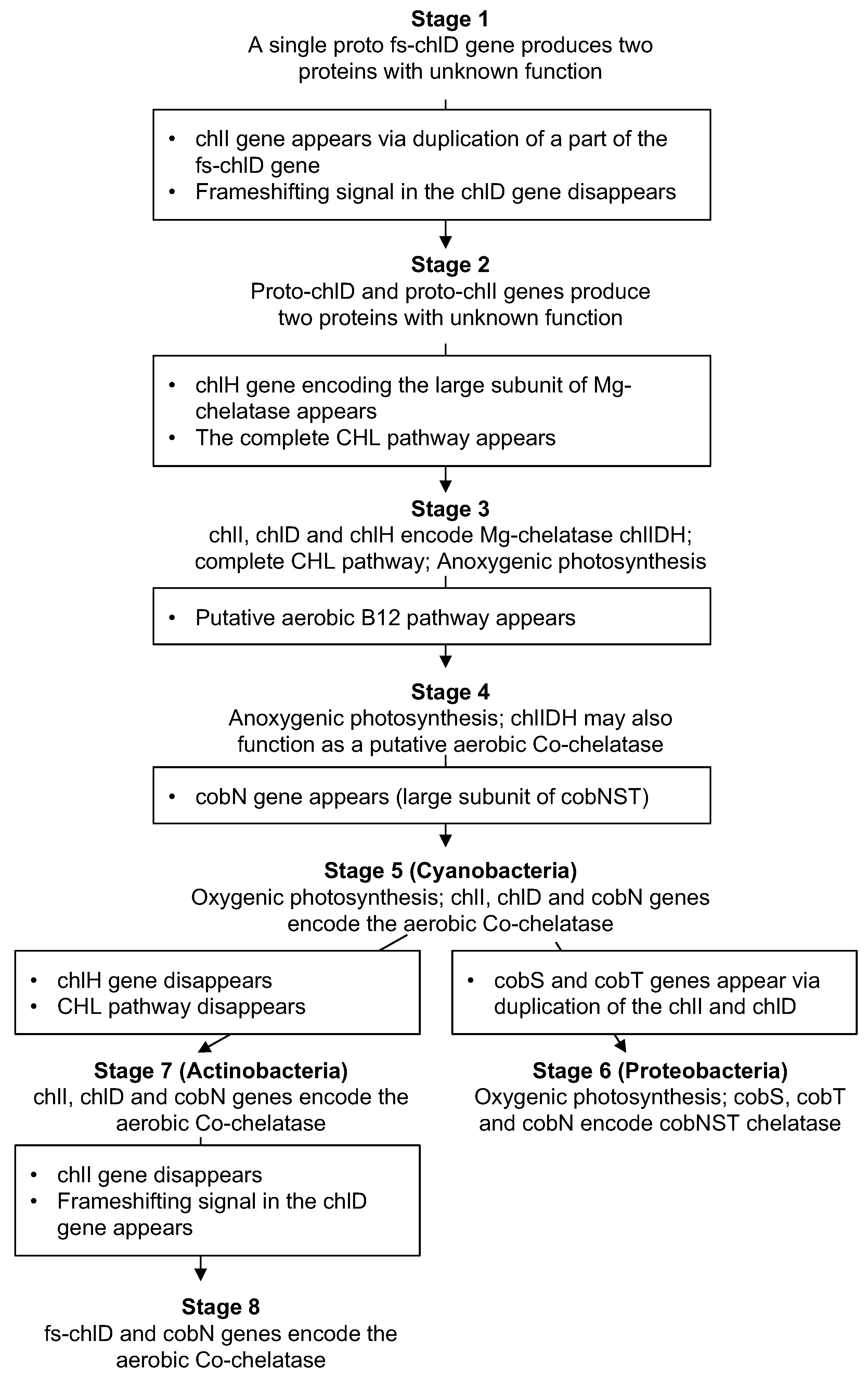
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| B12 | vitamin B12 or cobalamine (contains Co2+) |
| BCHL | bacteriochlorophyll (contains Mg2+) |
| CHL | chlorophyll (contains Mg2+) |
| F430 | coenzyme F430 (contains Ni2+) |
| fs-chlD gene | frameshifted chlD gene |
| Proto | Protoporphyrin IX |
| Uro | Uroporphyrinogen III |
References
- Blankenship, R.E.; Hartman, H. The origin and evolution of oxygenic photosynthesis. Trends Biochem. Sci. 1998, 23, 94–97. [Google Scholar] [CrossRef]
- Blankenship, R.E. Early evolution of photosynthesis. Plant Physiol. 2010, 154, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Willows, R.; Hansson, M. Mechanism, structure, and regulation of magnesium chelatase. Porphyr. Handb. II 2003, 113, 1–48. [Google Scholar]
- Thweatt, J.L.; Canniffe, D.P.; Bryant, D.A. Biosynthesis of chlorophylls and bacteriochlorophylls in green bacteria. Adv. Bot. Res. 2019, 90, 35–89. [Google Scholar]
- Guilard, R.; Kadish, K.M.; Smith, K.M.; Guilard, R. The Porphyrin Handbook; Academic Press: New York, NY, USA, 2003; Volume 18. [Google Scholar]
- Walker, C.J.; Willows, R.D. Mechanism and regulation of Mg-chelatase. Biochem. J. 1997, 327 Pt 2, 321–333. [Google Scholar] [CrossRef]
- Bollivar, D.W.; Suzuki, J.Y.; Beatty, J.T.; Dobrowolski, J.M.; Bauer, C.E. Directed mutational analysis of bacteriochlorophyll a biosynthesis in Rhodobacter capsulatus. J. Mol. Biol. 1994, 237, 622–640. [Google Scholar] [CrossRef]
- Gibson, L.; Willows, R.D.; Kannangara, C.G.; von Wettstein, D.; Hunter, C.N. Magnesium-protoporphyrin chelatase of Rhodobacter sphaeroides: Reconstitution of activity by combining the products of the bchH,-I, and-D genes expressed in Escherichia coli. Proc. Natl. Acad. Sci. USA 1995, 92, 1941–1944. [Google Scholar] [CrossRef]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B12: A review and future perspectives. Microb. Cell Factories 2017, 16, 1–14. [Google Scholar] [CrossRef]
- Crespo, A.; Blanco-Cabra, N.; Torrents, E. Aerobic vitamin B12 biosynthesis is essential for Pseudomonas aeruginosa class II ribonucleotide reductase activity during planktonic and biofilm growth. Front. Microbiol. 2018, 9, 986. [Google Scholar] [CrossRef]
- Fodje, M.; Hansson, A.; Hansson, M.; Olsen, J.; Gough, S.; Willows, R.; Al-Karadaghi, S. Interplay between an AAA module and an integrin I domain may regulate the function of magnesium chelatase. J. Mol. Biol. 2001, 311, 111–122. [Google Scholar] [CrossRef]
- Escalante-Semerena, J.C.; Warren, M.J. Biosynthesis and use of cobalamin (B12). EcoSal Plus 2008, 3. [Google Scholar] [CrossRef]
- Lundqvist, J.; Elmlund, D.; Heldt, D.; Deery, E.; Söderberg, C.A.; Hansson, M.; Warren, M.; Al-Karadaghi, S. The AAA+ motor complex of subunits CobS and CobT of cobaltochelatase visualized by single particle electron microscopy. J. Struct. Biol. 2009, 167, 227–234. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. J. Biol. Chem. 2003, 278, 41148–41159. [Google Scholar] [CrossRef] [PubMed]
- Antonov, I.; Coakley, A.; Atkins, J.F.; Baranov, P.V.; Borodovsky, M. Identification of the nature of reading frame transitions observed in prokaryotic genomes. Nucleic. Acids Res. 2013, 41, 6514–6530. [Google Scholar] [CrossRef] [PubMed]
- Antonov, I.V. Two Cobalt Chelatase Subunits Can Be Generated from a Single chlD Gene via Programed Frameshifting. Mol. Biol. Evol. 2020, 37, 2268–2278. [Google Scholar] [CrossRef] [PubMed]
- Dailey, H.A. Illuminating the black box of B12 biosynthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 14823–14824. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.H.; Zu Siederdissen, C.H.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Debussche, L.; Couder, M.; Thibaut, D.; Cameron, B.; Crouzet, J.; Blanche, F. Assay, purification, and characterization of cobaltochelatase, a unique complex enzyme catalyzing cobalt insertion in hydrogenobyrinic acid a, c-diamide during coenzyme B12 biosynthesis in Pseudomonas denitrificans. J. Bacteriol. 1992, 174, 7445–7451. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef] [PubMed]
- Angelov, A.; Liebl, S.; Ballschmiter, M.; Bomeke, M.; Lehmann, R.; Liesegang, H.; Daniel, R.; Liebl, W. Genome sequence of the polysaccharide-degrading, thermophilic anaerobe Spirochaeta thermophila DSM 6192. J. Bacteriol. 2010, 192, 6492–6493. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Mai, U.; Pfeiffer, W.; Janssen, S.; Asnicar, F.; Sanders, J.G.; Belda-Ferre, P.; Al-Ghalith, G.A.; Kopylova, E.; McDonald, D.; et al. Phylogenomics of 10,575 genomes reveals evolutionary proximity between domains Bacteria and Archaea. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Atkins, J.F.; Loughran, G.; Bhatt, P.R.; Firth, A.E.; Baranov, P.V. Ribosomal frameshifting and transcriptional slippage: From genetic steganography and cryptography to adventitious use. Nucleic Acids Res. 2016, 44, 7007–7078. [Google Scholar] [CrossRef]
- Cameron, B.; Guilhot, C.; Blanche, F.; Cauchois, L.; Rouyez, M.; Rigault, S.; Levy-Schil, S.; Crouzet, J. Genetic and sequence analyses of a Pseudomonas denitrificans DNA fragment containing two cob genes. J. Bacteriol. 1991, 173, 6058–6065. [Google Scholar] [CrossRef]
- Sousa, F.L.; Shavit-Grievink, L.; Allen, J.F.; Martin, W.F. Chlorophyll biosynthesis gene evolution indicates photosystem gene duplication, not photosystem merger, at the origin of oxygenic photosynthesis. Genome Biol. Evol. 2013, 5, 200–216. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Yamada, T.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int. J. Syst. Evol. Microbiol. 2003, 53, 1843–1851. [Google Scholar] [CrossRef]
- Hahn, M.W.; Schmidt, J.; Pitt, A.; Taipale, S.J.; Lang, E. Reclassification of four Polynucleobacter necessarius strains as representatives of Polynucleobacter asymbioticus comb. nov., Polynucleobacter duraquae sp. nov., Polynucleobacter yangtzensis sp. nov. and Polynucleobacter sinensis sp. nov., and emended description of Polynucleobacter necessarius. Int. J. Syst. Evol. Microbiol. 2016, 66, 2883. [Google Scholar]
- Challacombe, J.F.; Majid, S.; Deole, R.; Brettin, T.S.; Bruce, D.; Delano, S.F.; Detter, J.C.; Gleasner, C.D.; Han, C.S.; Misra, M.; et al. Complete genome sequence of Halorhodospira halophila SL1. Stand. Genom. Sci. 2013, 8, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Weaver, P. Fermentation and anaerobic respiration by Rhodospirillum rubrum and Rhodopseudomonas capsulata. J. Bacteriol. 1982, 149, 181–190. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Stage | Example Species (Phila) | Synthesis of the Medium and Small Proteins | Function of the Medium and Small Proteins | B12 Pathway (Anaerobic) | Large Subunit (chlH/cobN) | CHL Pathway | B12 Pathway (Aerobic) |
|---|---|---|---|---|---|---|---|
| 1 | Spirochaeta thermophila (Spirochaetes) Brevefilum fermentans (Chloroflexi) |  | Unknown | - | - | - | - |
| 2 | Thermodesulfobacterium commune (Thermodesulfobacteria) Thermodesulfovibrio yellowstonii (Nitrospirae) |  | Unknown | - | - | - | - |
| 3 | Polynucleobacter duraquae (Proteobacteria) |  | Mg-chelatase (small and medium subunits) | - |  | Yes | - |
| 4 | Halorhodospira halophila (Proteobacteria) Thiocystis violascens (Proteobacteria) |  | Mg-(Co-?)chelatase (small and medium subunits) | Yes |  | Yes | Yes? |
| 5 | Synechococcus elongatus (Cyanobacteria) Prochlorococcus marinus (Cyanobacteria) |  | Mg- and Co- chelatases (small and medium subunits) | Yes |  | Yes | Yes |
| 6 | Rhodospirillum rubrum (Proteobacteria) Dinoroseobacter shibae (Proteobacteria) Rhodovulum sulfidophilum (Proteobacteria) |  | Mg- and Co- chelatases (small and medium subunits) | Yes |  | Yes | Yes |
| 7 | Mycobacterium tuberculosis (Actinobacteria) Prauserella marina (Actinobacteria) |  | Small and medium subunits of Co- chelatases | Yes |  | - | Yes |
| 8 | Pseudomonas aeruginosa (Proteobacteria) Methanocaldococcus fervens (Archaea) |  | Small and medium subunits of Co- chelatases | Yes |  | - | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuznetsov, S.; Milenkin, A.; Antonov, I. Translational Frameshifting in the chlD Gene Gives a Clue to the Coevolution of the Chlorophyll and Cobalamin Biosyntheses. Microorganisms 2022, 10, 1200. https://doi.org/10.3390/microorganisms10061200
Kuznetsov S, Milenkin A, Antonov I. Translational Frameshifting in the chlD Gene Gives a Clue to the Coevolution of the Chlorophyll and Cobalamin Biosyntheses. Microorganisms. 2022; 10(6):1200. https://doi.org/10.3390/microorganisms10061200
Chicago/Turabian StyleKuznetsov, Stepan, Alexander Milenkin, and Ivan Antonov. 2022. "Translational Frameshifting in the chlD Gene Gives a Clue to the Coevolution of the Chlorophyll and Cobalamin Biosyntheses" Microorganisms 10, no. 6: 1200. https://doi.org/10.3390/microorganisms10061200