
Genome Sequence and Characterization of a Xanthorhodopsin-Containing, Aerobic Anoxygenic Phototrophic Rhodobacter Species, Isolated from Mesophilic Conditions at Yellowstone National Park

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation
2.2. Cultivation
2.3. Genome Sequencing
2.4. Whole Genome Comparison
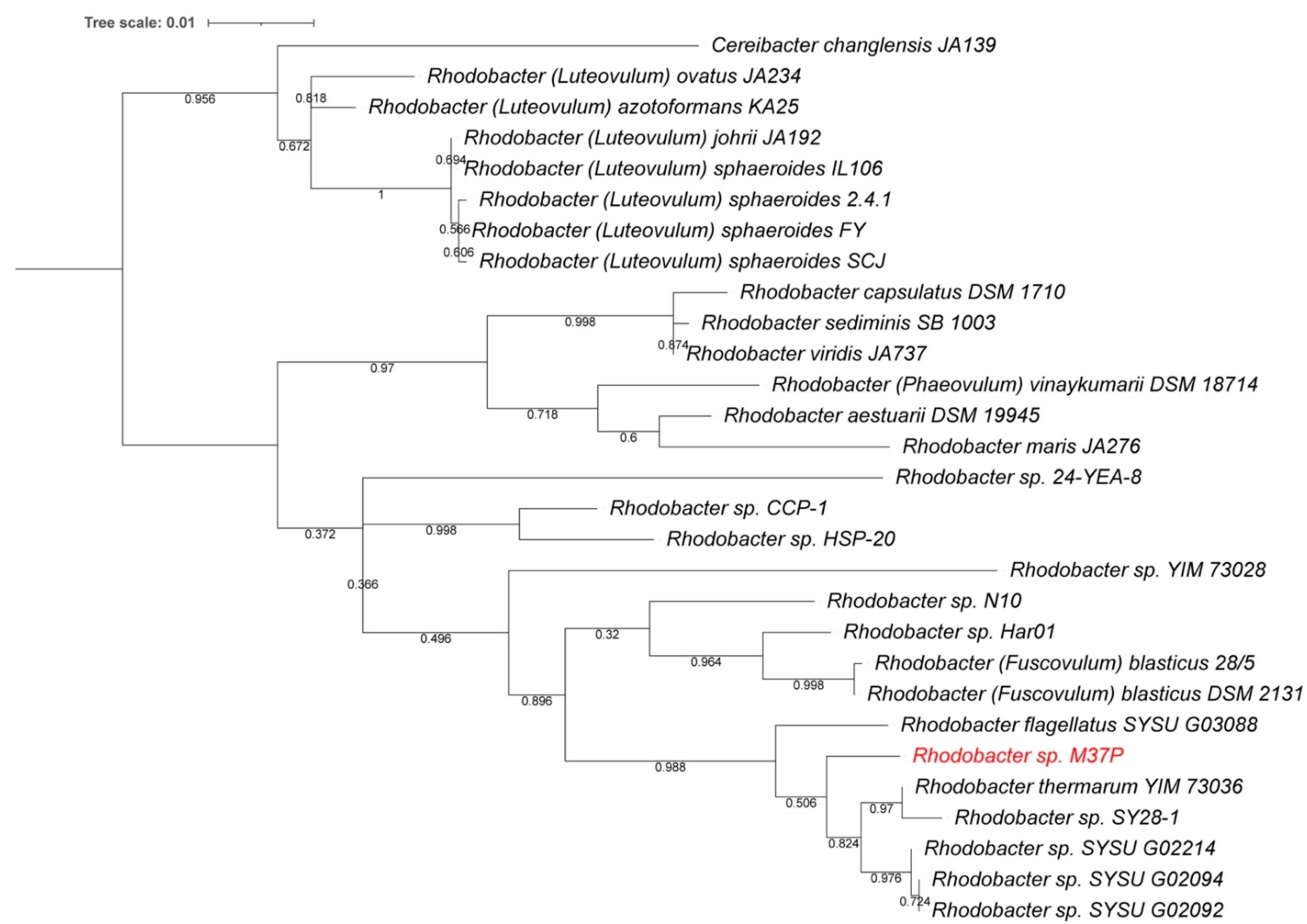
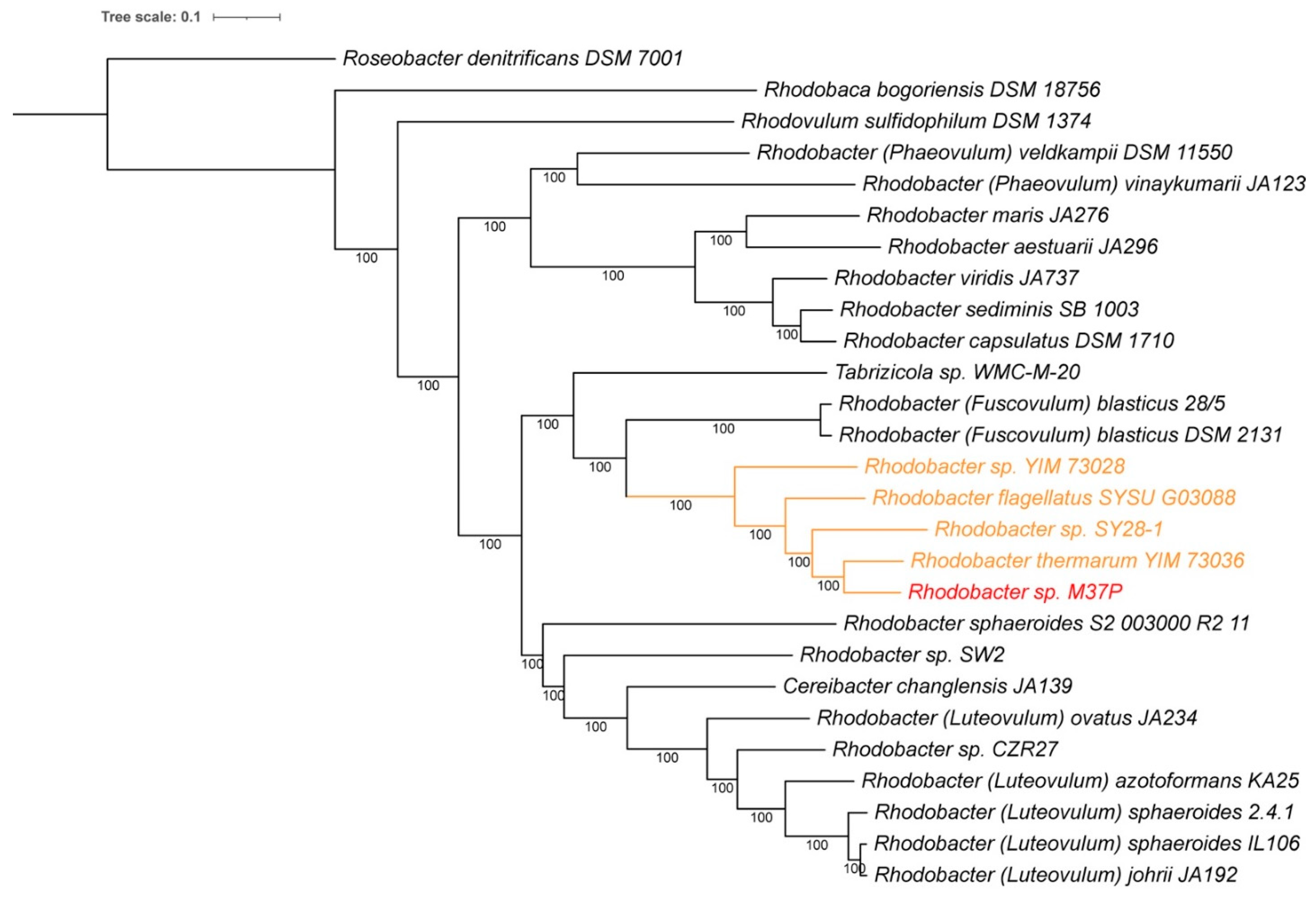
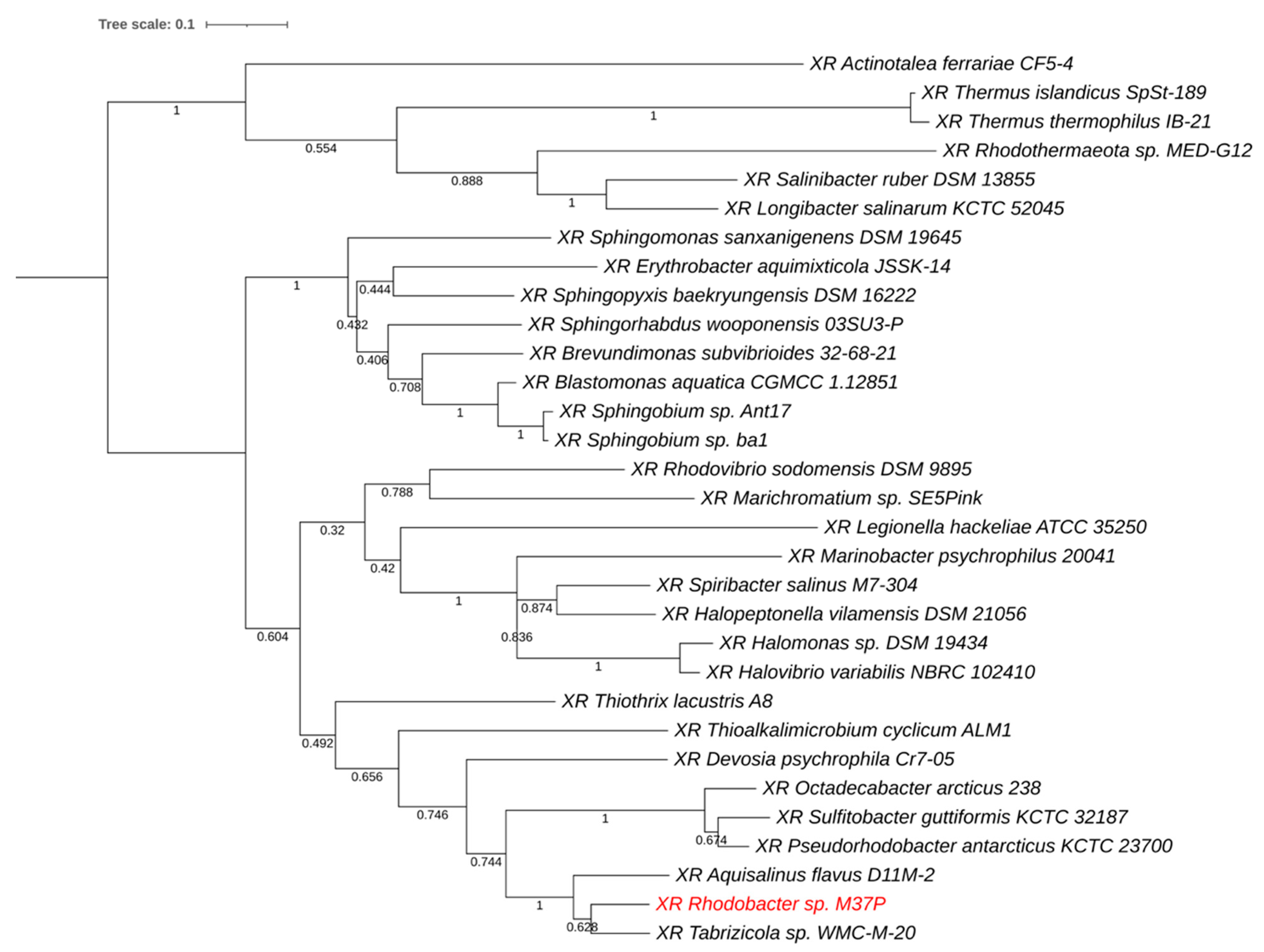
2.5. Single-Gene Phylogenetic Comparisons
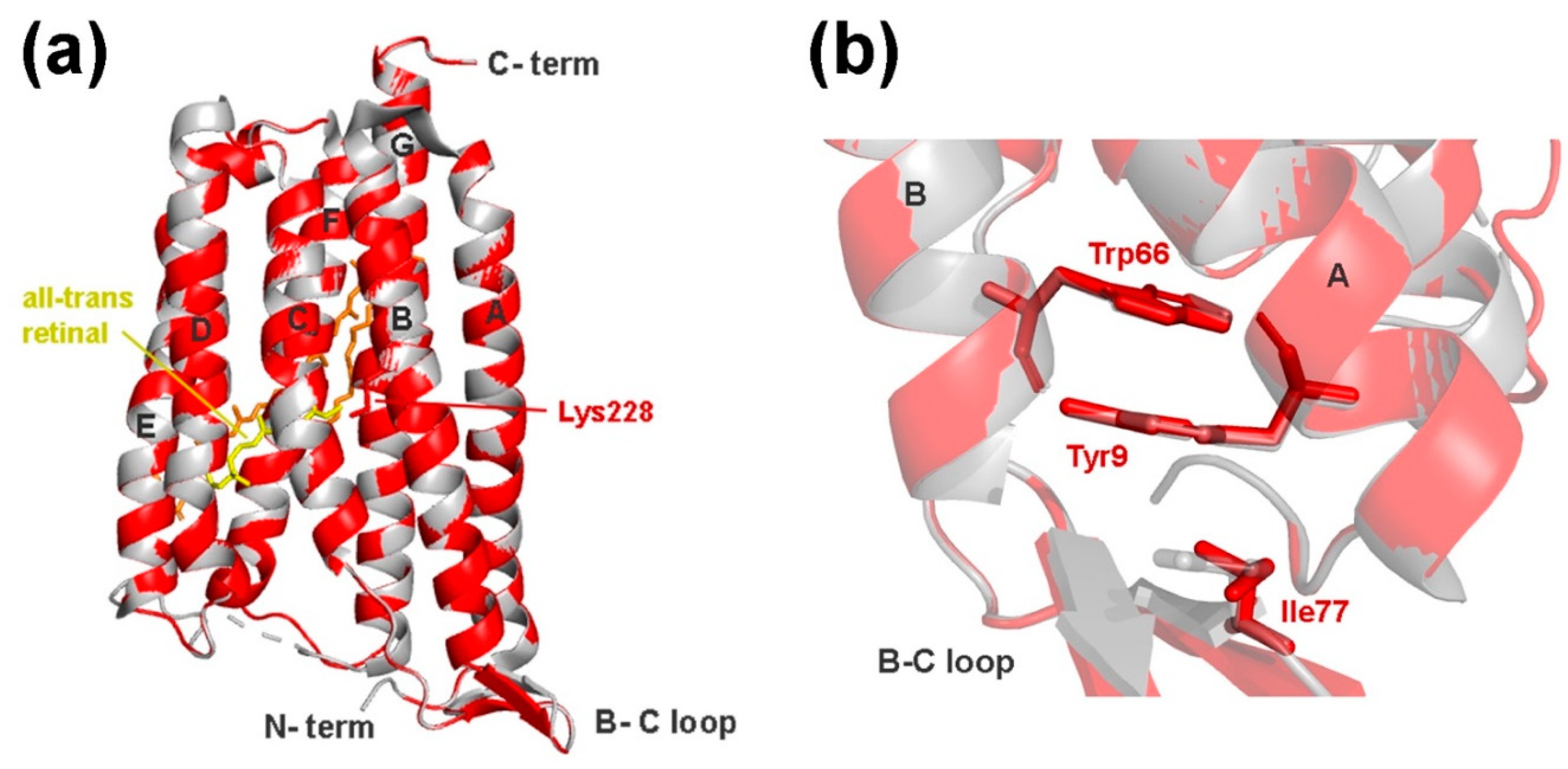
2.6. Structure Modeling
3. Results and Discussion
3.1. Genome Analysis
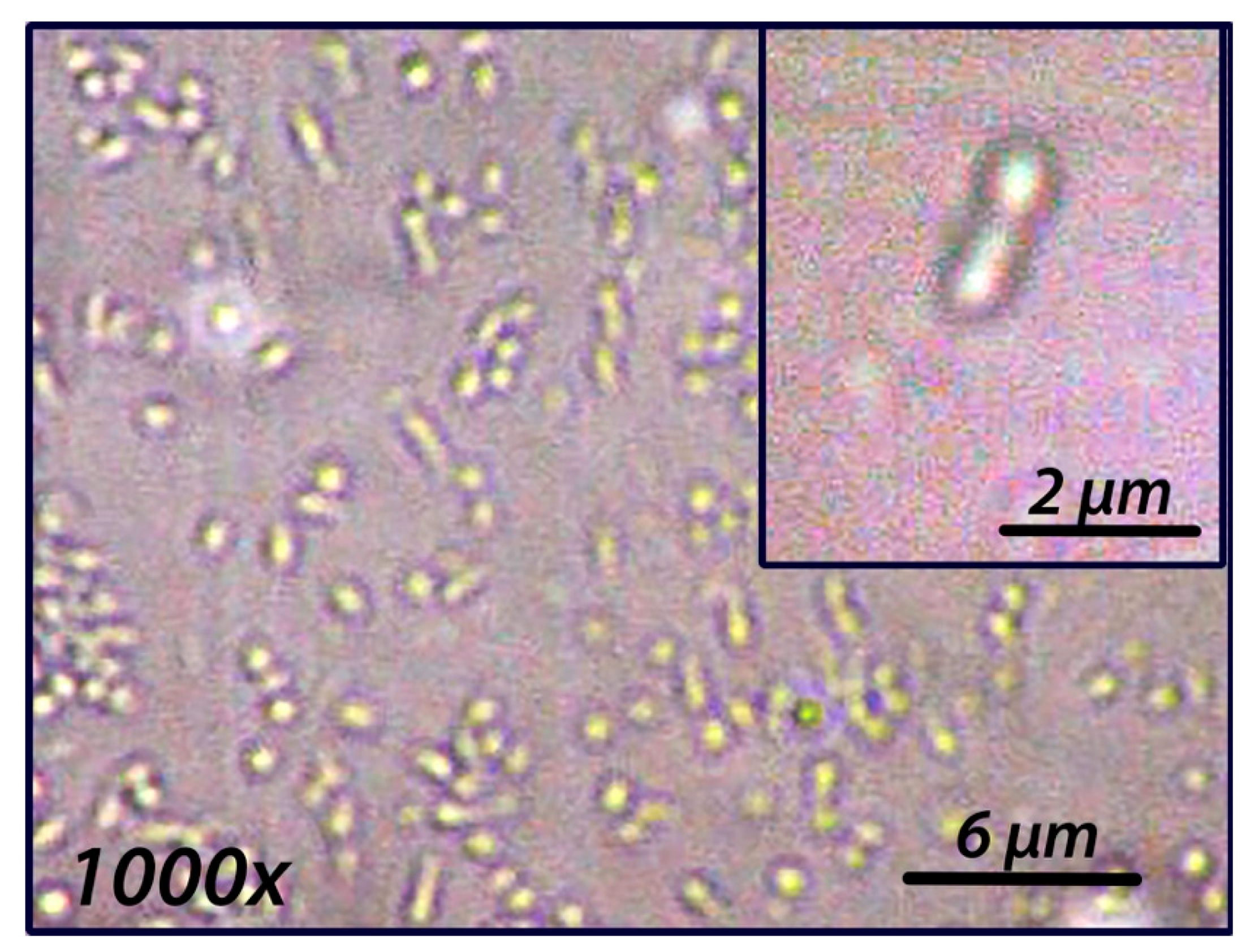
3.2. Species and Growth Characteristics
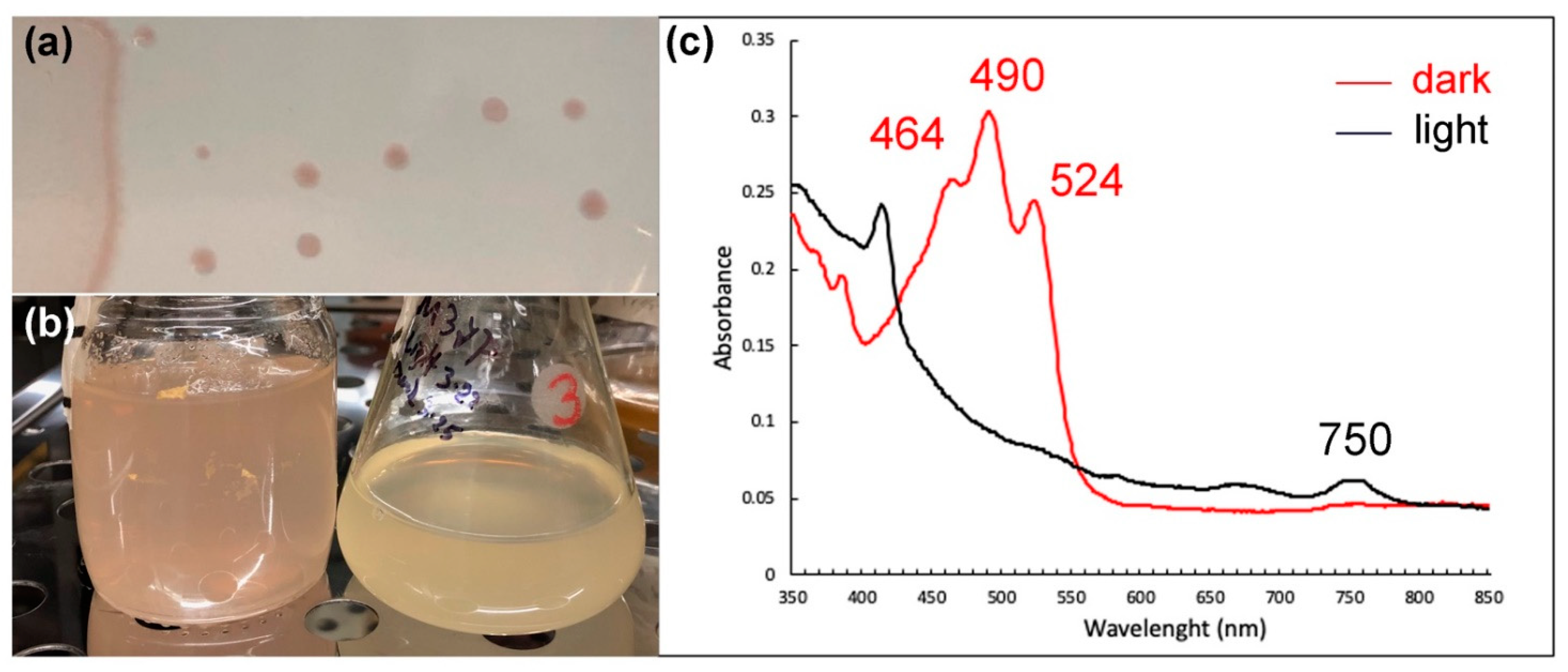
3.3. Xanthorhodopsin
3.4. Xanthorhodopsin in Mushroom Spring Microbial Mats
3.5. Description of Rhodobacter calidifons sp. nov.
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imhoff, J.F.; Truper, H.G.; Pfennig, N. Rearrangement of the species and genera of the phototrophic “Purple Nonsulfur Bacteria”. Int. J. Syst. Bacteriol. 1984, 34, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F. Genus Rhodobacter. Imhoff, Trüper and Pfennig 1984, 342VP. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2005; Volume 2, Part C; pp. 161–167. [Google Scholar]
- Kwon, S.Y.; Jiang, S.N.; Zheng, J.H.; Choy, H.E.; Min, J.J. Rhodobacter sphaeroides, a novel tumor-targeting bacteria that emits natural near-infrared fluorescence. Microbiol. Immunol. 2014, 58, 172–179. [Google Scholar] [CrossRef]
- Christ, W.J.; Asano, O.; Robidoux, A.L.C.; Perez, M.; Wang, Y.; Dubuc, G.R.; Gavin, W.E.; Hawkins, L.D.; McGuinness, P.D.; Mullarkey, M.A.; et al. E5531, a pure endotoxin antagonist of high potency. Science 1995, 268, 80–83. [Google Scholar] [CrossRef] [PubMed]
- A Mullarkey, M.; Rose, J.R.; Bristol, J.R.; Kawata, T.; Kimura, A.; Kobayashi, S.; Przetak, M.; Chow, J.; Gusovsky, F.; Christ, W.J.; et al. Inhibition of endotoxin response by e5564, a novel Toll-Like Receptor 4-directed endotoxin entagonist. J. Pharmacol. Exp. Ther. 2003, 304, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Kanie, Y.; Yamaguchi, Y.; Hayashi, A.; Uzawa, J.; Hatakeyama, M.; Hidaka, Y.; Toda, N.; Nakamura, S.; Kanie, O. Structural analysis of a novel lipooligosaccharide (LOS) from Rhodobacter azotoformans. Carbohydr. Res. 2019, 473, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Kamimura, D.; Hasebe, R.; Uchida, M.; Abe, N.; Yamamoto, R.; Jiang, J.J.; Hidaka, Y.; Nakanishi, Y.; Fujita, S.; et al. Rhodobacter azotoformans LPS (RAP99-LPS) is a TLR4 agonist that inhibits lung metastasis and enhances TLR3-mediated chemokine expression. Front. Immunol. 2021, 12, 675909. [Google Scholar] [CrossRef]
- Eckersley, K.; Dow, C.S. Rhodopseudomonas blastica sp. nov: A member of the Rhodospirillaceae. J. Gen. Microbiol. 1980, 119, 465–473. [Google Scholar]
- Khan, I.U.; Habib, N.; Xiao, M.; Li, M.M.; Xian, W.D.; Hejazi, M.S.; Tarhriz, V.; Zhi, X.Y.; Li, W.J. Rhodobacter thermarum sp. nov., a novel phototrophic bacterium isolated from sediment of a hot spring. Antonie Van Leeuwenhoek. 2019, 112, 867–875. [Google Scholar] [CrossRef]
- Venkata Ramana, V.; Anil Kumar, P.; Srinivas, T.N.; Sasikala, C.; Ramana, C.V. Rhodobacter aestuarii sp. nov., a phototrophic alphaproteo bacterium isolated from an estuarine environment. Int. J. Syst. Evol. Microbiol. 2009, 59 Pt 5, 1133–1136. [Google Scholar] [CrossRef] [PubMed]
- Gandham, S.; Lodha, T.; Chintalapati, S.; Chintalapati, V.R. Rhodobacter alkalitolerans sp. nov., isolated from an alkaline brown pond. Arch. Microbiol. 2018, 200, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Chang, T.H.; Yang, C.C.; Sheu, D.S.; Jheng, L.C.; Sheu, S.Y. Rhodobacter amnigenus sp. nov. and Rhodobacter ruber sp. nov., isolated from freshwater habitats. Int. J. Syst. Evol. Microbiol. 2021, 71, 005150. [Google Scholar] [CrossRef]
- Suresh, G.; Sailaja, B.; Ashif, A.; Dave, B.P.; Sasikala, C.; Ramana, C.V. Description of Rhodobacter azollae sp. nov. and Rhodobacter lacus sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Muramatsu, K.; Ueda, Y. Molecular genetic analyses of Rhodobacter azotoformans sp. nov. and related species of phototrophic bacteria. Syst. Appl. Microbiol. 1996, 19, 168–177. [Google Scholar] [CrossRef]
- Molisch, H. Die Purpurbakterien Nach Neuen Untersuchungen; G. Fischer: Jena, Germany, 1907. [Google Scholar]
- Xian, W.D.; Liu, Z.T.; Li, M.M.; Liu, L.; Ming, Y.Z.; Xiao, M.; Salam, N.; Li, W.J. Rhodobacter flagellatus sp. nov., a thermophilic bacterium isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2020, 70, 1541–1546. [Google Scholar] [CrossRef]
- Girija, K.R.; Sasikala, C.; Ramana, C.V.; Spröer, C.; Takaichi, S.; Thiel, V.; Imhoff, J.F. Rhodobacter johrii sp. nov., an endospore-producing cryptic species isolated from semi-arid tropical soils. Int. J. Syst. Evol. Microbiol. 2010, 60, 2099–2107. [Google Scholar] [CrossRef] [Green Version]
- Venkata Ramana, V.; Sasikala, C.; Ramana, C.V. Rhodobacter maris sp. nov., a phototrophic alphaproteobacterium isolated from a marine habitat of India. Int. J. Syst. Evol. Microbiol. 2008, 58, 1719–1722. [Google Scholar] [CrossRef]
- Srinivas, T.N.; Anil Kumar, P.; Sasikala, C.; Spröer, C.; Ramana, C.V. Rhodobacter ovatus sp. nov., a phototrophic alphaproteobacterium isolated from a polluted pond. Int. J. Syst. Evol. Microbiol. 2008, 58, 1379–1383. [Google Scholar] [CrossRef] [PubMed]
- Subhash, Y.; Lee, S.S. Rhodobacter sediminis sp. nov., isolated from lagoon sediments. Int. J. Syst. Evol. Microbiol. 2016, 66, 2965–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, G.; Dhanesh, K.; Krishnaiah, A.; Sasikala, C.; Ramana, C.V. Rhodobacter sediminicola sp. nov., isolated from a fresh water pond. Int. J. Syst. Evol. Microbiol. 2020, 70, 1294–1299. [Google Scholar] [CrossRef]
- Van Niel, C.B. The culture, general physiology, morphology and classification of the nonsulfur purple and brown bacteria. Bacteriol. Rev. 1944, 8, 1–118. [Google Scholar] [CrossRef]
- Sheu, C.; Li, Z.H.; Sheu, S.Y.; Yang, C.C.; Chen, W.M. Tabrizicola oligotrophica sp. nov. and Rhodobacter tardus sp. nov., two new species of bacteria belonging to the family Rhodobacteraceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 6266–6283. [Google Scholar] [CrossRef]
- Hansen, T.A.; Imhoff, J.F. Rhodobacter veldkampii, a new species of Phototrophic Purple Nonsulfur Bacteria. Int. J. Syst. Evol. Microbiol. 1985, 35, 115–116. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, T.N.R.; Anil Kumar, P.; Sasikala, C.; Ramana, C.V.; Imhoff, J.F. Rhodobacter vinaykumarii sp. nov., a marine phototrophic alphaproteobacterium from tidal waters, and emended description of the genus Rhodobacter. Int. J. Syst. Evol. Microbiol. 2007, 57, 1984–1987. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.S.; Ramaprasad, E.V.V.; Vaseef, S.; Sasikala, C.; Ramana, C.V. Rhodobacter viridis sp. nov., a phototrophic bacterium isolated from mud of a stream. Int. J. Syst. Evol. Microbiol. 2013, 63, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Wang, T.; Chen, Z.; Li, Y.; Yao, L. Rhodobacter xinxiangensis sp. nov., isolated from pakchoi-cultivated soil contaminated with heavy metal and its potential to reduce Cd and Pb accumulation in pakchoi (Brassica campestris L.). Arch. Microbiol. 2020, 202, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Lodha, T.D.; Indu, B.; Sasikala, C.; Ramana, C.V. Taxogenomics resolves conflict in the genus Rhodobacter: A two and half decades pending thought to reclassify the genus Rhodobacter. Front. Microbiol. 2019, 10, 2480. [Google Scholar] [CrossRef] [Green Version]
- Ramaley, R.F.; Hixson, J. Isolation of a nonpigmented, thermophilic bacterium similar to Thermus aquaticus. J. Bacteriol. 1970, 103, 527–528. [Google Scholar] [CrossRef] [Green Version]
- Weaver, P.; Wall, J.D.; Gest, H. Characterization of Rhodopseudomonas capsulata. Arch. Microbiol. 1975, 105, 207–216. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. Bypassing cultivation to identify bacterial species. Microbe 2014, 9, 111–118. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J.J.S. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.J.B. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Steinegger, M.; Meier, M.; Mirdita, M.; Vöhringer, H.; Haunsberger, S.J.; Söding, J. HH-suite3 for fast remote homology detection and deep protein annotation. BMC Bioinform. 2019, 20, 473. [Google Scholar] [CrossRef] [Green Version]
- Mirdita, M.; von den Driesch, L.; Galiez, C.; Martin, M.J.; Söding, J.; Steinegger, M. Uniclust databases of clustered and deeply annotated protein sequences and alignments. Nucleic Acids Res. 2016, 45, D170–D176. [Google Scholar] [CrossRef] [PubMed]
- Studer, G.; Tauriello, G.; Bienert, S.; Biasini, M.; Johner, N.; Schwede, T. ProMod3—A versatile homology modelling toolbox. PLoS Comp. Biol. 2021, 17, e1008667. [Google Scholar] [CrossRef]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, G.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef]
- Satoh, T.; Hoshino, Y.; Kitamura, H. Rhodopseudomonas sphaeroides forma sp. denitrificans, a denitrifying strain as a subspecies of Rhodopseudomonas sphaeroides. Arch. Microbiol. 1976, 108, 265–269. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Yurkov, V.V.; Beatty, J.T. Aerobic Anoxygenic Phototropic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 695–724. [Google Scholar] [CrossRef] [Green Version]
- Yurkov, V.; Hughes, E. Genes associated with the peculiar phenotypes of the Aerobic Anoxygenic Phototrophs. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2013; pp. 327–358. [Google Scholar]
- Kusian, B.; Bowien, B. Organization and regulation of CBB CO2 assimilation genes in autotrophic bacteria. FEMS Microbiol. Rev. 1997, 21, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Ashida, H.; Danchin, A.; Yokota, A. Was photosynthetic RuBisCO recruited by acquisitive evolution from RuBisCO-like proteins involved in sulfur metabolism? Res. Microbiol. 2005, 156, 611–618. [Google Scholar] [CrossRef]
- Badger, M.R.; Bek, E.J. Multiple Rubisco forms in proteobacteria: Their functional significance in relation to CO2 acquisition by the CBB cycle. J. Exp. Bot. 2008, 59, 1525–1541. [Google Scholar] [CrossRef] [Green Version]
- Kyndt, J.A.; Aviles, F.A.; Imhoff, J.F.; Künzel, S.; Neulinger, S.C.; Meyer, T.E. Comparative genome analysis of the photosynthetic betaproteobacteria of the genus Rhodocyclus: Heterogeneity within strains assigned to Rhodocyclus tenuis and description of Rhodocyclus gracilis sp. nov. as a new species. Microorganisms 2022, 10, 649. [Google Scholar] [CrossRef]
- Imker, H.J.; Fedorov, A.A.; Fedorov, E.V.; Almo, S.C.; Gerlt, J.A. Mechanistic diversity in the RuBisCO superfamily: The “enolase” in the methionine salvage pathway in Geobacillus kaustophilus. Biochemistry 2007, 46, 4077–4089. [Google Scholar] [CrossRef]
- Pemberton, J.M.; Horne, I.M.; McEwan, A.G. Regulation of photosynthetic gene expression in purple bacteria. Microbiology 1998, 144, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Gregor, J.; Klug, G. Regulation of bacterial photosynthesis genes by oxygen and light. FEMS Microbiol. Lett. 1999, 179, 1–9. [Google Scholar] [CrossRef]
- Yurkov, V.; van Gemerden, H. Impact of light/dark regime on growth rate, biomass formation and bacteriochlorophyll synthesis in Erythromicrobium hydrolyticum. Arch. Microbiol. 1993, 159, 84–89. [Google Scholar] [CrossRef]
- Takamiya, K.I.; Shioi, Y.; Shimada, H.; Arata, H. Inhibition of accumulation of bacteriochlorophyll and carotenoids by blue light in an aerobic photosynthetic bacterium Roseobacter denitrificans, during anaerobic respiration. Plant Cell Physiol. 1992, 33, 1171–1174. [Google Scholar]
- Yurkov, V.; Krasilnikova, E.N.; Gorlenko, V.M. Effect of light and oxygen on metabolism of the aerobic bacterium Erythromicrobium sibiricum. Microbiology 1993, 62, 35–38. [Google Scholar]
- Cartron, M.L.; Olsen, J.D.; Sener, M.; Jackson, P.J.; Brindley, A.A.; Qian, P.; Dickman, M.J.; Leggett, G.J.; Schulten, K.; Neil Hunter, C. Integration of energy and electron transfer processes in the photosynthetic membrane of Rhodobacter sphaeroides. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.D.; Tucker, J.D.; Timney, J.A.; Qian, P.; Vassilev, C.; Hunter, C.N. The organization of LH2 complexes in membranes from Rhodobacter sphaeroides. J. Biol. Chem. 2008, 283, 30772–30779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrielsen, M.; Gardiner, A.T.; Cogdell, R. Peripheral complexes of purple bacteria. In The Purple Phototrophic Bacteria, Advances in Photosynthesis and Respiration; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 28, pp. 135–153. [Google Scholar]
- Jaschke, P.R.; Leblanc, H.N.; Lang, A.S.; Beatty, J.T. The PucC protein of Rhodobacter capsulatus mitigates an inhibitory effect of light-harvesting 2 alpha and beta proteins on light-harvesting complex 1. Photosynth Res. 2008, 95, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Mothersole, D.J.; Jackson, P.J.; Vasilev, C.; Tucker, J.D.; Brindley, A.A.; Dickman, M.J.; Hunter, C.N. PucC and LhaA direct efficient assembly of the light-harvesting complexes in Rhodobacter sphaeroides. Mol. Microbiol. 2016, 99, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Selyanin, V.; Hauruseu, D.; Koblížek, M. The variability of light-harvesting complexes in aerobic anoxygenic phototrophs. Photosynth Res. 2016, 128, 35–43. [Google Scholar] [CrossRef]
- Wakao, N.; Shiba, T.; Hiraishi, A.; Ito, M.; Sakurai, Y. Distribution of bacteriochlorophyll a in species of the genus Acidiphilium. Curr. Microbiol. 1993, 27, 277–279. [Google Scholar] [CrossRef]
- Wakao, N.; Yokoi, N.; Isoyama, N.; Hiraishi, A.; Shimada, K.; Kobayashi, M.; Kise, H.; Iwaki, M.; Itoh, S.; Takaichi, S. Discovery of natural photosynthesis using Zn-containing bacteriochlorophyll in an aerobic bacterium Acidiphilium rubrum. Plant Cell Physiol. 1996, 37, 889–893. [Google Scholar] [CrossRef] [Green Version]
- Koblížek, M. Ecology of aerobic anoxygenic phototrophs in aquatic environments. FEMS Microbiol. Rev. 2015, 39, 854–870. [Google Scholar] [CrossRef] [Green Version]
- Bekker, A.; Holland, H.D.; Wang, P.L.; Rumble, D., 3rd; Stein, H.J.; Hannah, J.L.; Coetzee, L.L.; Beukes, N.J. Dating the rise of atmospheric oxygen. Nature 2004, 427, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Koblížek, M.; Zeng, Y.; Horák, A.; Oborník, M. Regressive evolution of photosynthesis in the Roseobacter clade. In Advances in Botanical Research; Thomas Beatty, J., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2013; Volume 66, pp. 385–405. [Google Scholar] [CrossRef]
- Balashov, S.P.; Imasheva, E.S.; Boichenko, V.A.; Antón, J.; Wang, J.M.; Lanyi, J.K. Xanthorhodopsin: A proton pump with a light-harvesting carotenoid antenna. Science 2005, 309, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanyi, J.K.; Balashov, S.P. Xanthorhodopsin: A bacteriorhodopsin-like proton pump with a carotenoid antenna. Biochim. Biophys. Acta 2008, 1777, 684–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morizumi, T.; Ou, W.L.; Van Eps, N.; Inoue, K.; Kandori, H.; Brown, L.S.; Ernst, O.P. X-ray crystallographic structure and oligomerization of Gloeobacter rhodopsin. Sci. Rep. 2019, 9, 11283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.-P.; Liu, Y.; Wang, F.; Zhou, Y.-G.; Liu, H.-C.; Liu, Z.-P. Aquisalinus flavus gen. nov., sp. nov., a member of the family Parvularculaceae isolated from a saline lake. Int. J. Syst. Evol. Microbiol. 2016, 66, 1813–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.X.; Dorji, P.; Liu, H.C.; Li, A.H.; Zhou, Y.G. Tabrizicola sediminis sp. nov., one aerobic anoxygenic photoheterotrophic bacteria from sediment of saline lake. Int. J. Syst. Evol. Microbiol. 2019, 69, 2565–2570. [Google Scholar] [CrossRef] [PubMed]
- Woggon, W.-D. Oxidative cleavage of carotenoids catalyzed by enzyme models and beta-carotene 15,15′-monooxygenase. Pure Appl. Chem. 2002, 74, 1397–1408. [Google Scholar] [CrossRef]
- Kim, J.; Smith, J.J.; Tian, L.; Dellapenna, D. The evolution and function of carotenoid hydroxylases in Arabidopsis. Plant Cell Physiol. 2009, 50, 463–479. [Google Scholar] [CrossRef] [Green Version]
- Canniffe, D.P.; Hitchcock, A. Photosynthesis: Carotenoids in Photosynthesis—Structure and Biosynthesis. In Encyclopedia of Biological Chemistry III, 3rd ed.; Jez, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 163–185. ISBN 9780128220405. [Google Scholar]
- Maresca, J.A.; Graham, J.E.; Wu, M.; Eisen, J.A.; Bryant, D.A. Identification of a fourth family of lycopene cyclases in photosynthetic bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 11784–11789. [Google Scholar] [CrossRef] [Green Version]
- Frigaard, N.U.; Martinez, A.; Mincer, T.J.; DeLong, E.F. Proteorhodopsin lateral gene transfer between marine planktonic Bacteria and Archaea. Nature 2006, 439, 847–850. [Google Scholar] [CrossRef]
- Finkel, O.; Béjà, O.; Belkin, S. Global abundance of microbial rhodopsins. ISME J. 2013, 7, 448–451. [Google Scholar] [CrossRef] [Green Version]
- Gushchin, I.; Shevchenko, V.; Polovinkin, V.; Kovalev, K.; Alekseev, A.; Round, E.; Borshchevskiy, V.; Balandin, T.; Popov, A.; Gensch, T.; et al. Crystal structure of a light-driven sodium pump. Nat. Struct. Mol. Biol. 2015, 22, 390–395. [Google Scholar] [CrossRef]
- Kato, H.E.; Inoue, K.; Abe-Yoshizumi, R.; Kato, Y.; Ono, H.; Konno, M.; Hososhima, S.; Ishizuka, T.; Hoque, M.R.; Kunitomo, H.; et al. Structural basis for Na(+) transport mechanism by a light-driven Na(+) pump. Nature 2015, 521, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Boichenko, V.A.; Wang, J.M.; Antón, J.; Lanyi, J.K.; Balashov, S.P. Functions of carotenoids in xanthorhodopsin and archaerhodopsin, from action spectra of photoinhibition of cell respiration. Biochim. Biophys. Acta–Bioenerg. 2006, 1757, 1649–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, V.; Hügler, M.; Ward, D.M.; Bryant, D.A. The dark side of the Mushroom Spring microbial mat: Life in the shadow of Chlorophototrophs. II. Metabolic functions of abundant community members predicted from metagenomic snalyses. Front. Microbiol. 2017, 8, 943. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Consarnau, L.; González, J.M.; Coll-Lladó, M.; Gourdon, P.; Pascher, T.; Neutze, R.; Pedrós-Alió, C.; Pinhassi, J. Light stimulates growth of proteorhodopsin-containing marine Flavobacteria. Nature 2007, 445, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Consarnau, L.; Akram, N.; Lindell, K.; Pedersen, A.; Neutze, R.; Milton, D.L.; González, J.M.; Pinhassi, J. Proteorhodopsin phototrophy promotes survival of marine bacteria during starvation. PLoS Biol. 2010, 8, e1000358. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; O’Shaughnessy, T.J.; Soto, C.M.; Rahbar, A.M.; Robertson, K.L.; Lebedev, N.; Vora, G.J. Function and regulation of Vibrio campbellii Proteorhodopsin: Acquired phototrophy in a classical organoheterotroph. PLoS ONE 2012, 7, e38749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, Y.; Yoshizawa, S.; Nakajima, Y.; Watanabe, M.; Fukunaga, T.; Ogura, Y.; Hayashi, T.; Oshima, K.; Hattori, M.; Ikeuchi, M.; et al. Solar-panel and parasol strategies shape the proteorhodopsin distribution pattern in marine Flavobacteria. ISME J. 2018, 12, 1329–1343. [Google Scholar] [CrossRef] [Green Version]
- Thiel, V.; Wood, J.M.; Olsen, M.T.; Tank, M.; Klatt, C.G.; Ward, D.M.; Bryant, D.A. The dark side of the Mushroom Spring microbial mat: Life in the shadow of Chlorophototrophs. I. Microbial diversity based on 16S rRNA gene amplicons and metagenomic sequencing. Front. Microbiol. 2016, 7, 919. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genome | Growth | Rhodopsin | Rubisco | LH II | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| size (Mb) | GC % | CDS | Genbank Acc. | malate | succinate | glutamate | glucose | temp. optimum | anaerobic growth | |||||
| Rba. sp. M37P | 3.63 | 67.5 | 3732 | JAANHS000000000 | Y | Y | Y | Y | 38–39 | N | Xantho-rhodopsin | none | N | this paper |
| Rba. thermarum YIM73036 | 3.65 | 66.8 | 3695 | QMJY00000000 | N | Y | N | Y | 37–45 | N | none | none | N | [9] |
| Rba. flagellatus SYSU G03088 | 3.62 | 67.7 | 3677 | VMDU00000000 | N | n.d. | N | N | 45 | n.d. | none | rbcS; rbcL | N | [16] |
| Rba. blasticus DSM 2131 | 3.59 | 66.5 | 3625 | PZKE00000000 | Y | n.d. | Y | Y | 30–35 | Y | none | rbcII; rbcS; rbcL | Y | [8] |
| Rba. capsulatus DSM 1710 | 3.67 | 66.5 | 3601 | QKZO00000000 | Y | Y | Y | Y | 30 | Y | none | rbcII; rbcS; rbcL | Y | [1,15] |
| Rba. sediminis SB1003 | 3.87 | 66.6 | 3728 | CP001312 | Y | n.d. | Y | Y | 30–35 | Y | none | rbcII; rbcS; rbcL | Y | [20] |
| Rba. viridis JA737 | 3.87 | 65.3 | 3780 | QJTK00000000 | N | N | N | Y | 30 | Y | none | rbcII | Y | [26] |
| Rba. ovatus JA234 | 3.81 | 66.5 | 3775 | OAOQ00000000 | N | Y | N | N | 25–30 | Y | none | rbcII; rbcS; rbcL | Y | [19] |
| Rba. vinaykumarii JA123 | 3.49 | 68.2 | 3366 | OBMN00000000 | Y | Y | N | Y | 20–30 | Y | none | rbcII; rbcS; rbcL | Y | [25] |
| Rba. maris JA276 | 3.83 | 65.1 | 3806 | OBMT00000000 | Y | N | N | N | 25–35 | Y | none | rbcII; rbcS; rbcL | Y | [18] |
| Rba. aestuarii JA296 | 3.84 | 61.1 | 3742 | QAXT00000000 | N | N | N | N | 25–30 | Y | none | rbcII; rbcS; rbcL | Y | [10] |
| Rba. sphaeroides ATH 2.4.1 | 4.63 | 68.7 | 4657 | CP030271 | Y | Y | Y | Y | 30–34 | Y | none | rbcII; rbcS; rbcL | Y | [22] |
| Rba. sphaeroides IL106 | 4.79 | 69.1 | 4848 | QRBG00000000 | Y | Y | Y | Y | 30 | Y | none | rbcII; rbcS; rbcL | Y | [51] |
| Rba. azotoformans KA25 | 4.41 | 68.4 | 4505 | QAOT00000000 | Y | Y | Y | Y | 30–35 | Y | none | rbcII; rbcS; rbcL | Y | [14] |
| Rba. johri JA192 | 4.59 | 69.3 | 4438 | PZZW00000000 | Y | Y | Y | Y | 25–35 | Y | none | rbcII; rbcS; rbcL | Y | [17] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyndt, J.A.; Robertson, S.; Shoffstall, I.B.; Ramaley, R.F.; Meyer, T.E. Genome Sequence and Characterization of a Xanthorhodopsin-Containing, Aerobic Anoxygenic Phototrophic Rhodobacter Species, Isolated from Mesophilic Conditions at Yellowstone National Park. Microorganisms 2022, 10, 1169. https://doi.org/10.3390/microorganisms10061169
Kyndt JA, Robertson S, Shoffstall IB, Ramaley RF, Meyer TE. Genome Sequence and Characterization of a Xanthorhodopsin-Containing, Aerobic Anoxygenic Phototrophic Rhodobacter Species, Isolated from Mesophilic Conditions at Yellowstone National Park. Microorganisms. 2022; 10(6):1169. https://doi.org/10.3390/microorganisms10061169
Chicago/Turabian StyleKyndt, John A., Sydney Robertson, Isabella B. Shoffstall, Robert F. Ramaley, and Terrance E. Meyer. 2022. "Genome Sequence and Characterization of a Xanthorhodopsin-Containing, Aerobic Anoxygenic Phototrophic Rhodobacter Species, Isolated from Mesophilic Conditions at Yellowstone National Park" Microorganisms 10, no. 6: 1169. https://doi.org/10.3390/microorganisms10061169