
Taxonomic Re-Evaluation and Genomic Comparison of Novel Extracellular Electron Uptake-Capable Rhodovulum visakhapatnamense and Rhodovulum sulfidophilum Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Isolation and Cultivtion Conditions
2.2. Genome Sequencing
2.3. Phylogenetic Analysis, Taxonomic Re-Evaluation, and Comparative Genomics
2.4. RNA Isolation, RNA Sequencing, and Differential Expression Analysis
2.5. Supplemental Materials & Methods
3. Results and Discussion
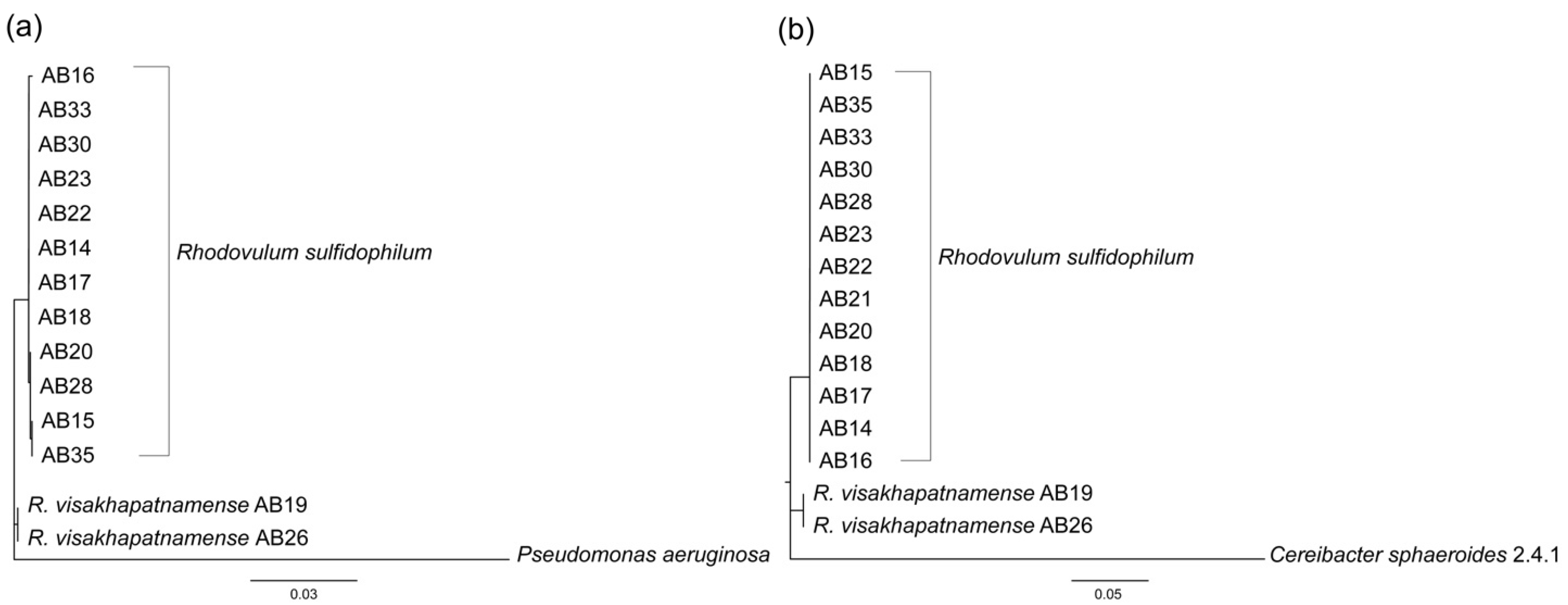
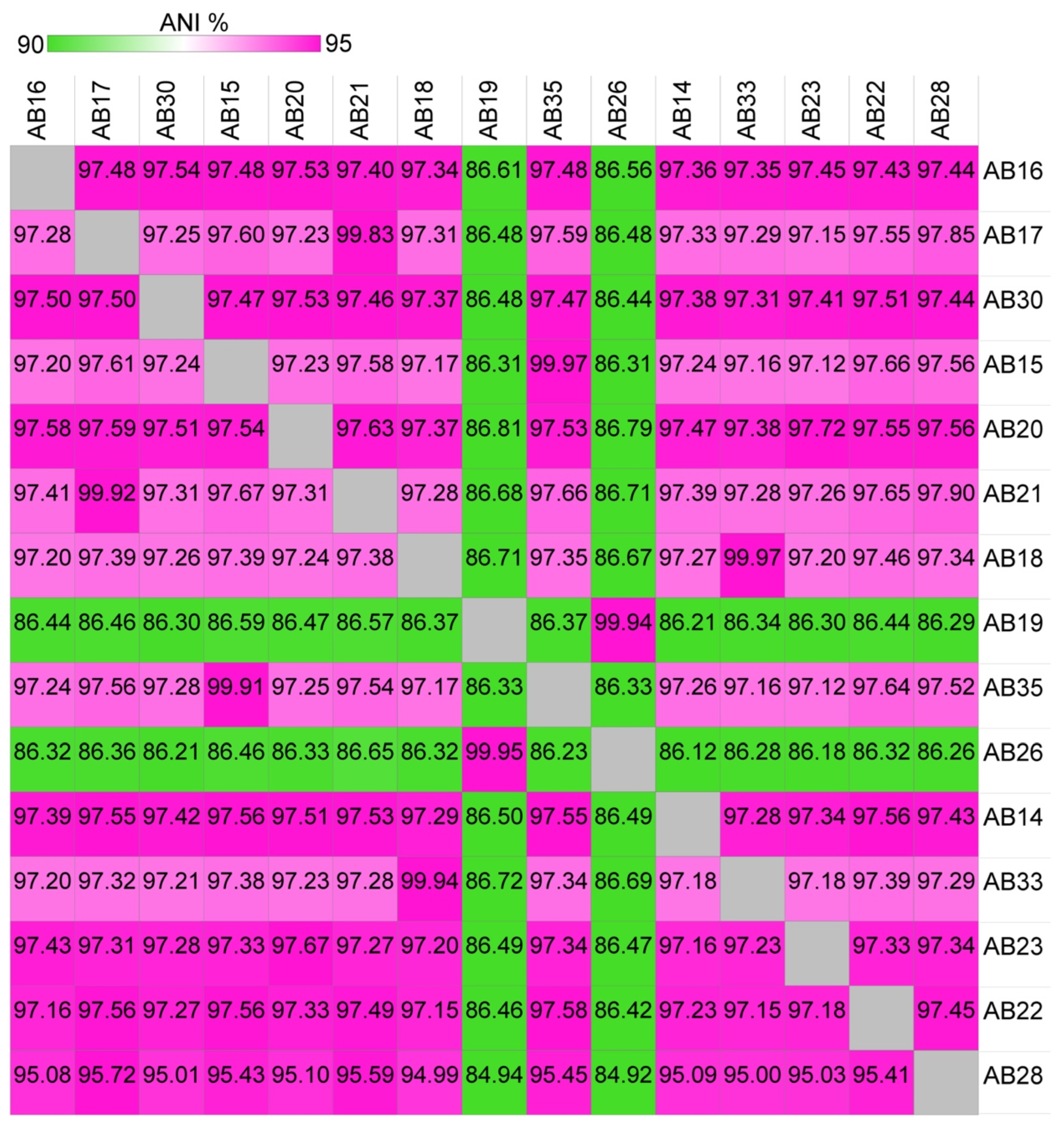
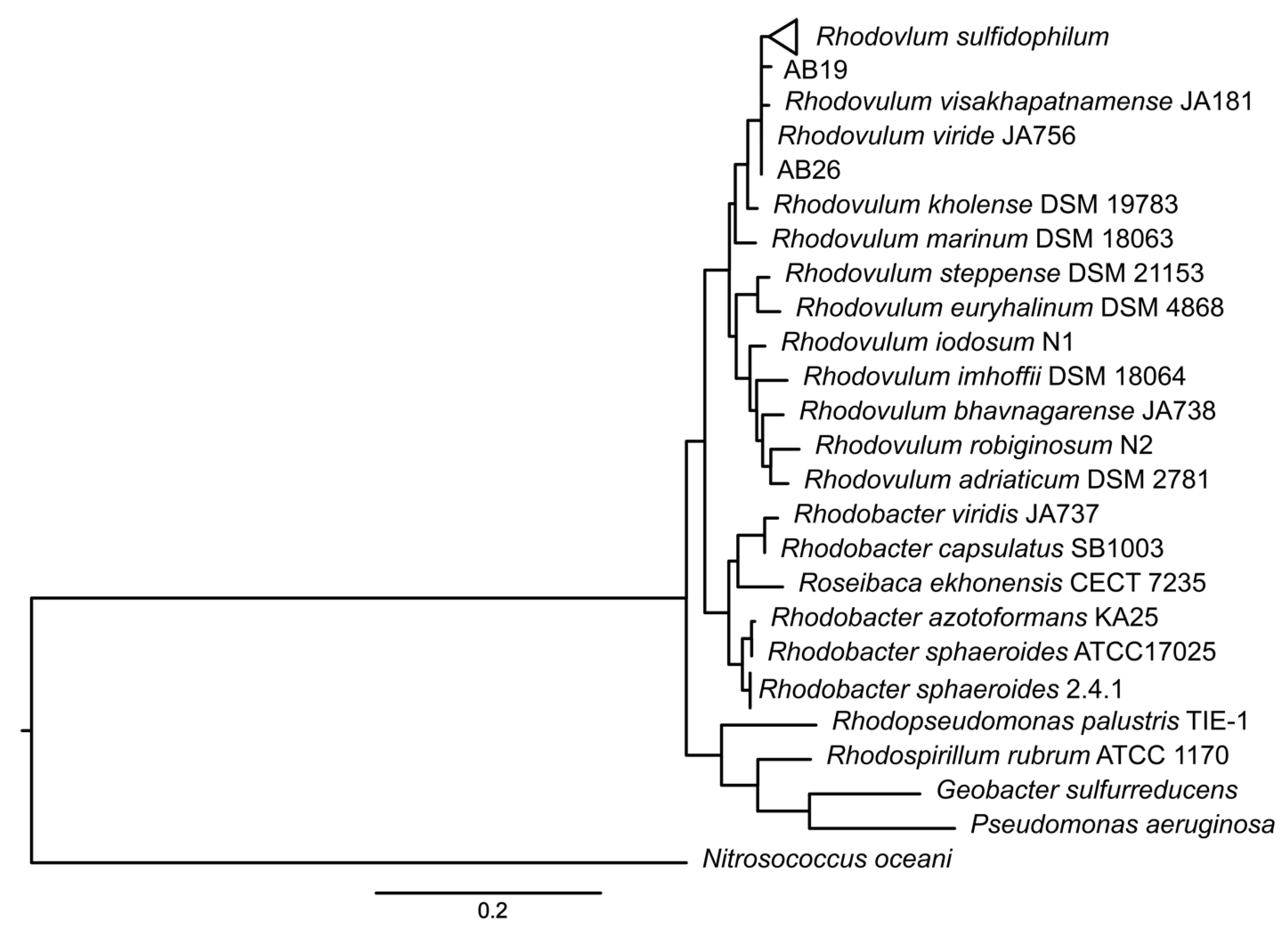
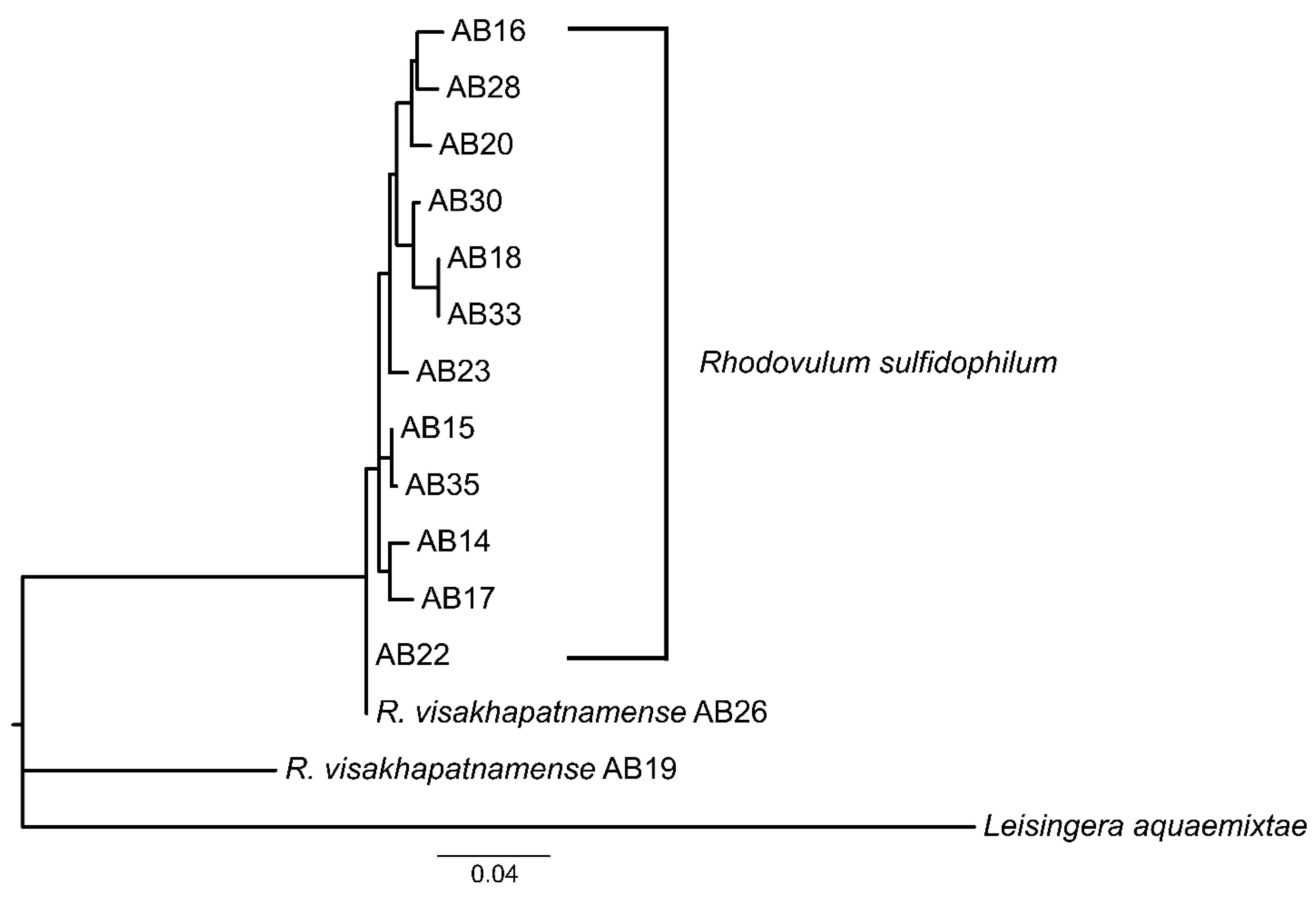
3.1. Taxonomic Re-Evaluation of Strain AB26 and Strain AB19
3.2. Strain AB26 Genome Features and Expression Analysis
3.2.1. Phototrophy
3.2.2. Carbon Dioxide Fixation
3.2.3. Reducing Power
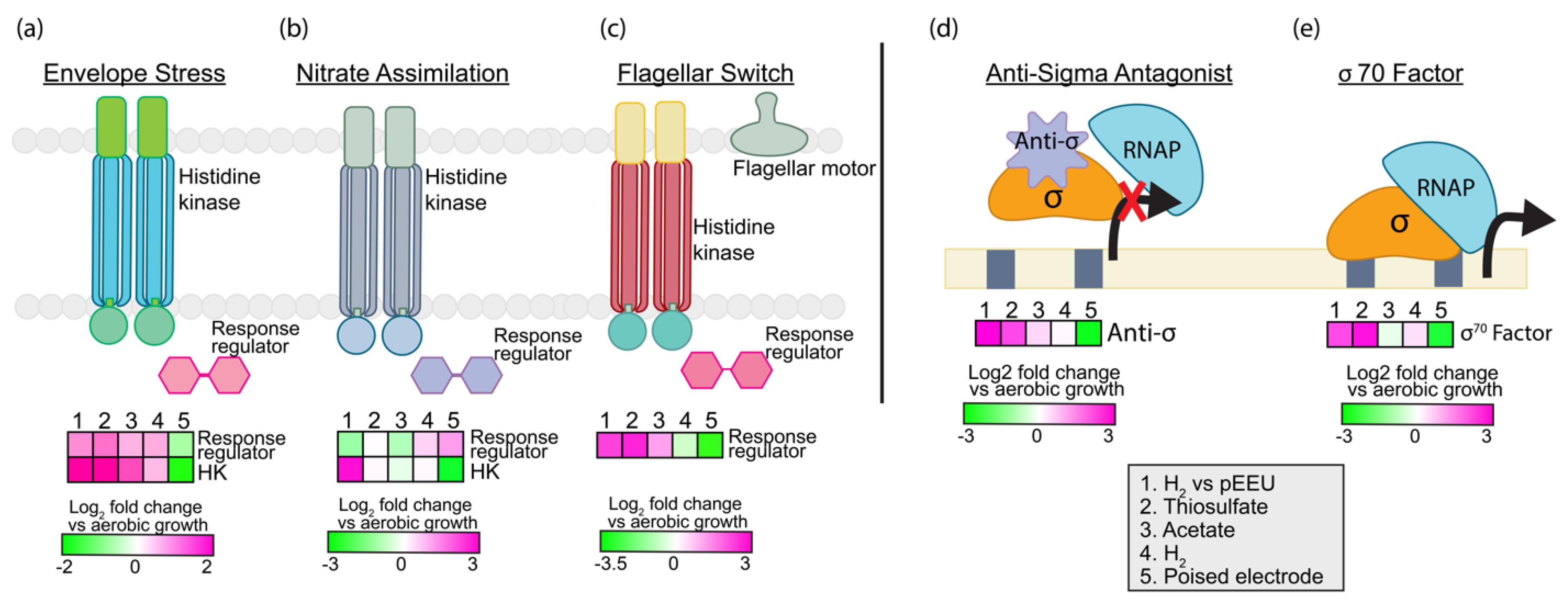
3.2.4. Regulation and Signaling
3.2.5. Biodegradation and Carbon Storage
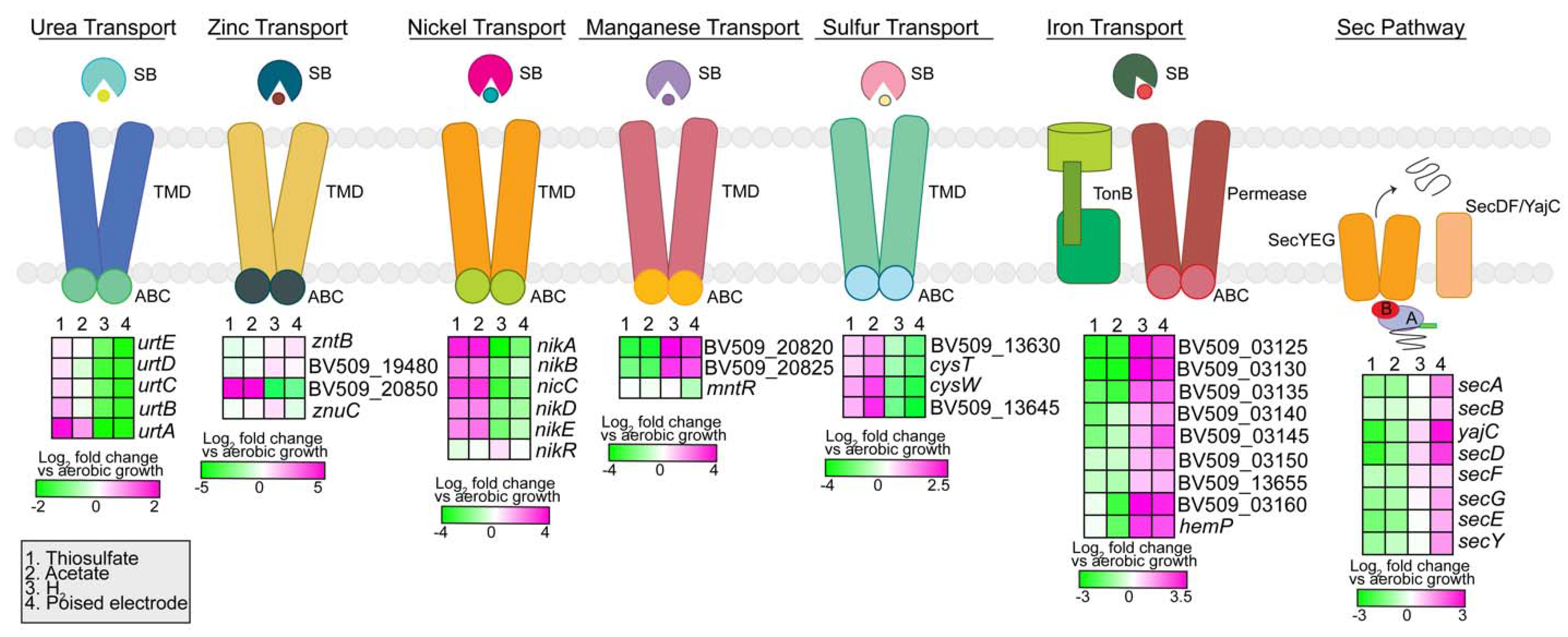
3.2.6. Transporters
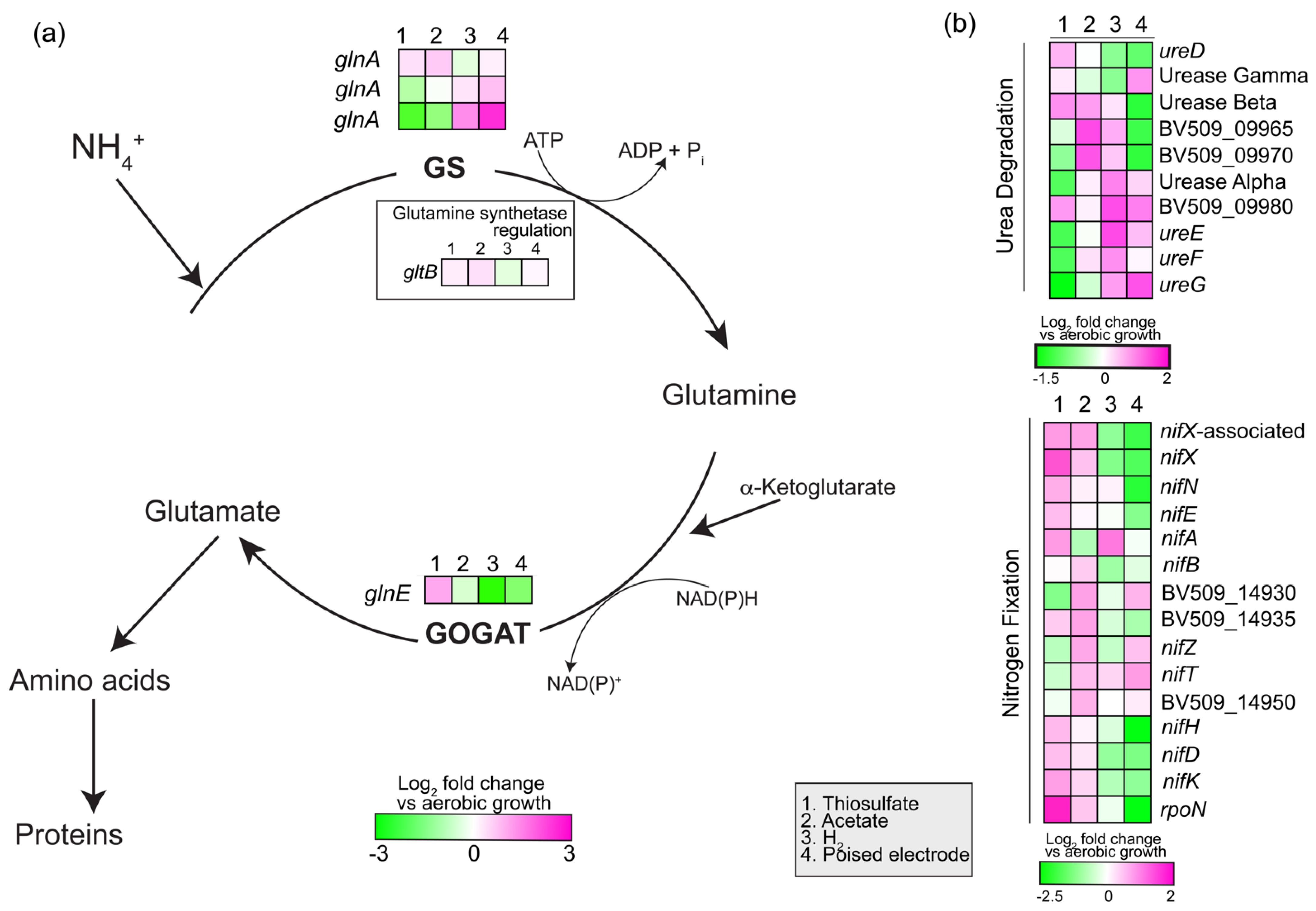
3.2.7. Nitrogen Assimilation and Fixation
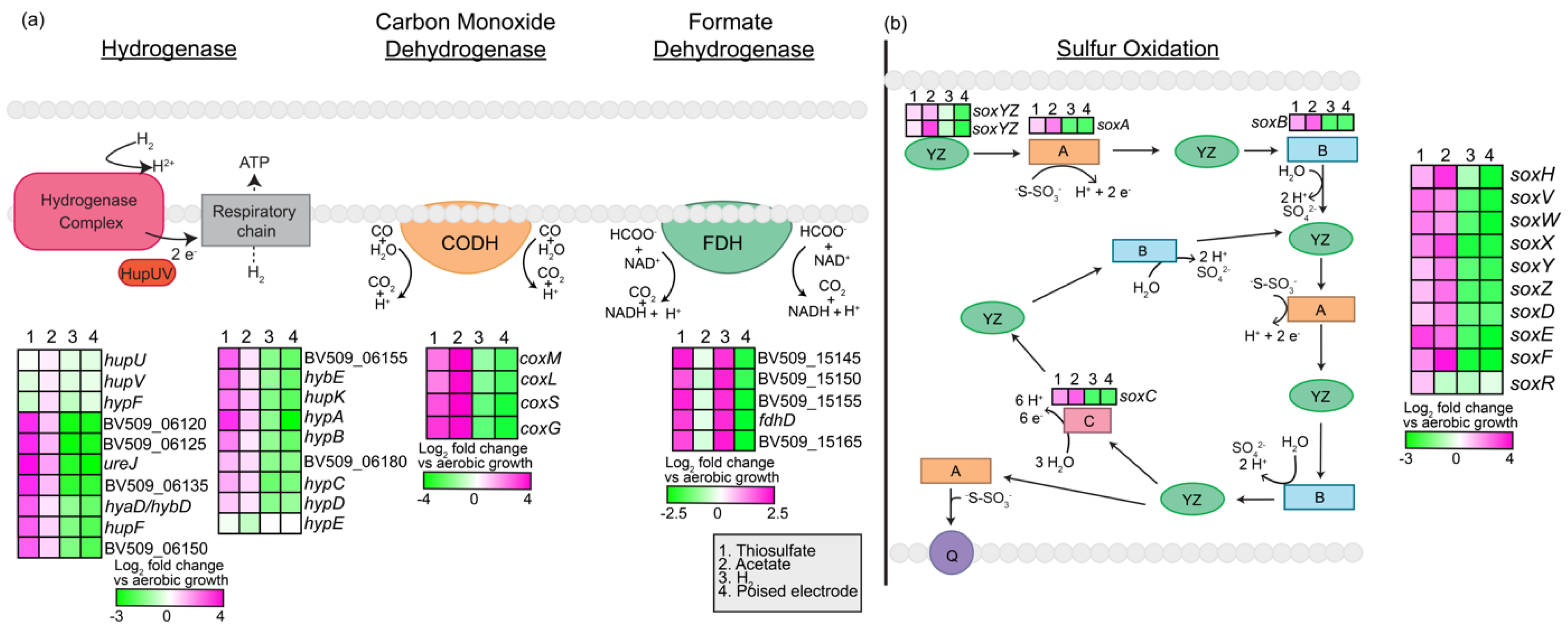
3.2.8. Extracellular Electron Uptake (EEU) Genes
3.3. RNA-Seq Led Genomic Comparison between Rhodovulum spp.
3.3.1. C-Type Cytochromes (Cytc)
Diheme Cytc EeuP
Hypothetical Protein BV509_18570
Diheme Cytc BV509_14915
Cytc BV509_15885
3.3.2. Transcriptional Regulators
Mar Family Transcription Regulator BV509_01095
Transcriptional Regulator BV509_15825
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shu, W.-S.; Huang, L.-N. Microbial diversity in extreme environments. Nat. Rev. Microbiol. 2022, 20, 219–235. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Aulenta, F.; Villano, M.; Angenent, L.T. Cathodes as electron donors for microbial metabolism: Which extracellular electron transfer mechanisms are involved? Bioresour. Technol. 2011, 102, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Guzman, M.S.; Bose, A. Extracellular electron uptake by autotrophic microbes: Physiological, ecological, and evolutionary implications. J. Ind. Microbiol. Biotechnol. 2020, 47, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The Microbial Engines That Drive Earth’s Biogeochemical Cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D.R.; Nevin, K.P. Electrobiocommodities: Powering microbial production of fuels and commodity chemicals from carbon dioxide with electricity. Curr. Opin. Biotechnol. 2013, 24, 385–390. [Google Scholar] [CrossRef]
- Rabaey, K.; Rodriguez, J.; Blackall, L.L.; Keller, J.; Gross, P.; Batstone, D.; Verstraete, W.; Nealson, K.H. Microbial ecology meets electrochemistry: Electricity-driven and driving communities. ISME J. 2007, 1, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef]
- Bose, A.; Gardel, E.J.; Vidoudez, C.; Parra, E.; Girguis, P.R. Electron uptake by iron-oxidizing phototrophic bacteria. Nat. Commun. 2014, 5, 3391. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.R.; Rajeev, P.; Jain, A.; Pirbadian, S.; Okamoto, A.; Gralnick, J.A.; El-Naggar, M.Y.; Nealson, K.H. Tracking electron uptake from a cathode into Shewanella cells: Implications for energy acquisition from solid-substrate electron donors. MBio 2018, 9, e02203-17. [Google Scholar] [CrossRef] [Green Version]
- Tefft, N.M.; TerAvest, M.A. Reversing an extracellular electron transfer pathway for electrode-driven acetoin reduction. ACS Synth. Biol. 2019, 8, 1590–1600. [Google Scholar] [CrossRef]
- Jiao, Y.; Newman, D.K. The pio operon is essential for phototrophic Fe (II) oxidation in Rhodopseudomonas palustris TIE-1. J. Bacteriol. 2007, 189, 1765–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Guzman, M.S.; Rengasamy, K.; Stoica, A.; Singh, R.; Ranaivoarisoa, T.O.; Davenport, E.J.; Bai, W.; McGinley, B.; Meacham, J.M.; et al. Photoferrotrophy and phototrophic extracellular electron uptake is common in the marine anoxygenic phototroph Rhodovulum sulfidophilum. ISME J. 2021, 15, 3384–3398. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.S.; Rengasamy, K.; Binkley, M.M.; Jones, C.; Ranaivoarisoa, T.O.; Singh, R.; Fike, D.A.; Meacham, J.M.; Bose, A. Phototrophic extracellular electron uptake is linked to carbon dioxide fixation in the bacterium Rhodopseudomonas palustris. Nat. Commun. 2019, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Guzman, M.S.; McGinley, B.; Santiago-Merced, N.; Gupta, D.; Bose, A.J.G.A. Draft genome sequences of three closely related isolates of the purple nonsulfur bacterium Rhodovulum sulfidophilum. Genome Announc. 2017, 5, e00029-17. [Google Scholar] [CrossRef] [Green Version]
- Guzman, M.S.; Bose, A. Draft genome sequences of four Rhodobacter sphaeroides strains isolated from a marine ecosystem. Microbiol. Resour. Announc. 2019, 8, e01648-18. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, T.; Kumar, P.A.; Sasikala, C.; Ramana, C.V.; Imhoff, J.F. Rhodovulum visakhapatnamense sp. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 1762–1764. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bosi, E.; Donati, B.; Galardini, M.; Brunetti, S.; Sagot, M.-F.; Lió, P.; Crescenzi, P.; Fani, R.; Fondi, M. MeDuSa: A multi-draft based scaffolder. Bioinformatics. 2015, 31, 2443–2451. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wu, J.; Yang, J.; Sun, S.; Xiao, J.; Yu, J. PGAP: Pan-genomes analysis pipeline. Bioinformatics 2011, 28, 416–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2015, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Béjà, O.; Suzuki, M.T.; Heidelberg, J.F.; Nelson, W.C.; Preston, C.M.; Hamada, T.; Eisen, J.A.; Fraser, C.M.; DeLong, E.F. Unsuspected diversity among marine aerobic anoxygenic phototrophs. Nature 2002, 415, 630–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yutin, N.; Suzuki, M.T.; Béjà, O. Novel primers reveal wider diversity among marine aerobic anoxygenic phototrophs. Appl. Environ. Microbiol. 2005, 71, 8958–8962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Ohta, T. On the stochastic model for estimation of mutational distance between homologous proteins. J. Mol. Evol. 1972, 2, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Nickens, D.G.; Bauer, C.E. Analysis of the puc operon promoter from Rhodobacter capsulatus. J. Bacteriol. 1998, 180, 4270–4277. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, T.; Leimkühler, S. The oxygen-tolerant and NAD+-dependent formate dehydrogenase from Rhodobacter capsulatus is able to catalyze the reduction of CO2 to formate. FEBS J. 2013, 280, 6083–6096. [Google Scholar] [CrossRef]
- Lahiri, C.; Mandal, S.; Ghosh, W.; Dam, B.; Roy, P. A novel gene cluster soxSRT is essential for the chemolithotrophic oxidation of thiosulfate and tetrathionate by Pseudaminobacter salicylatoxidans KCT001. Curr. Microbiol. 2006, 52, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Grabarczyk, D.B.; Berks, B.C. Intermediates in the Sox sulfur oxidation pathway are bound to a sulfane conjugate of the carrier protein SoxYZ. PLoS ONE 2017, 12, e0173395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larimer, F.W.; Chain, P.; Hauser, L.; Lamerdin, J.; Malfatti, S.; Do, L.; Land, M.L.; Pelletier, D.A.; Beatty, J.T.; Lang, A.S.; et al. Complete genome sequence of the metabolically versatile photosynthetic bacterium Rhodopseudomonas palustris. Nat. Biotechnol. 2004, 22, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravind, L.; Ponting, C.P. The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signalling proteins. FEMS Microbiol. Lett. 1999, 176, 111–116. [Google Scholar] [CrossRef]
- Kyndt, J.A.; Meyer, T.E.; Cusanovich, M.A. Photoactive yellow protein, bacteriophytochrome, and sensory rhodopsin in purple phototrophic bacteria. Photochem. Photobiol. Sci. 2004, 3, 519–530. [Google Scholar] [CrossRef]
- Martinez, S.E.; Beavo, J.A.; Hol, W.G. GAF domains: Two–billion–year–old molecular switches that bind cyclic nucleotides. Mol. Interv. 2002, 2, 317–323. [Google Scholar] [CrossRef]
- Hiraishi, A.; Kitamura, H. Distribution of phototrophic purple nonsulfur bacteria in activated sludge systems and other aquatic environments. Nippon Suisan Gakkaishi 1984, 50, 1929–1937. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, Y.; Chen, Z.; Wang, Y.; Zhu, F.; Cao, B.; Wu, Y.; Li, N. The organophosphorus pesticides in soil was degradated by Rhodobacter sphaeroides after wastewater treatment. Biochem. Eng. J. 2019, 141, 247–251. [Google Scholar] [CrossRef]
- Hassan, M.A.; Shirai, Y.; Kusubayashi, N.; Karim, M.I.A.; Nakanishi, K.; Hasimoto, K. The production of polyhydroxyalkanoate from anaerobically treated palm oil mill effluent by Rhodobacter sphaeroides. J. Biosci. Bioeng. 1997, 83, 485–488. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Lloyd, R.; Fuller, R.C. The accumulation of poly (3-hydroxyalkanoates) in Rhodobacter sphaeroides. Arch. Microbiol. 1991, 155, 337–340. [Google Scholar] [CrossRef]
- Ranaivoarisoa, T.O.; Singh, R.; Rengasamy, K.; Guzman, M.S.; Bose, A. Towards sustainable bioplastic production using the photoautotrophic bacterium Rhodopseudomonas palustris TIE-1. J. Ind. Microbiol. Biotechnol. 2019, 46, 1401–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.; Kelly, D.J.; Thomas, G.H. Tripartite ATP-independent periplasmic transporters: Application of a relational database for genome-wide analysis of transporter gene frequency and organization. J. Mol. Microbiol. Biotechnol. 2007, 12, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Rosso, K.M.; Zachara, J.M.; Fredrickson, J.K. Mtr extracellular electron-transfer pathways in Fe (III)-reducing or Fe (II)-oxidizing bacteria: A genomic perspective. Biochem. Soc. Trans. 2012, 40, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, A.; Kumar, B.V.; Sree, B.D.; Tushar, L.; Sasikala, C.; Ramana, C.V.J. Rhodovulum salis sp. nov. and Rhodovulum viride sp. nov., phototrophic Alphaproteobacteria isolated from marine habitats. Int. J. Syst. Evol. Microbiol. 2014, 64, 957–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao, N.; Hirose, Y.; Misawa, N.; Ohtsubo, Y.; Umekage, S.; Kikuchi, Y. Complete genome sequence of Rhodovulum sulfidophilum DSM 2351, an extracellular nucleic acid-producing bacterium. Genome Announc. 2015, 3, e00388-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, S.; Hori, K.; Maruyama, F.; Ren, S.; Sugimoto, S.; Yamamoto, N.; Mori, H.; Yamada, T.; Sato, S.; Tabata, S.; et al. Whole-genome sequence of the purple photosynthetic bacterium Rhodovulum sulfidophilum strain W4. Genome Announc. 2013, 1, e00577-13. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.W.; Daltrop, O.; Stevens, J.M.; Ferguson, S.J. C-type cytochromes: Diverse structures and biogenesis systems pose evolutionary problems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Estevez-Canales, M.; Kuzume, A.; Borjas, Z.; Füeg, M.; Lovley, D.; Wandlowski, T.; Esteve-Núñez, A. A severe reduction in the cytochrome C content of Geobacter sulfurreducens eliminates its capacity for extracellular electron transfer. Environ. Microbiol. Rep. 2015, 7, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Bagos, P.G.; Nikolaou, E.P.; Liakopoulos, T.D.; Tsirigos, K.D. Combined prediction of Tat and Sec signal peptides with hidden Markov models. Bioinformatics 2010, 26, 2811–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Size (bp) | 4,380,746 | |
|---|---|---|
| GC% | 67.9 | |
| Scaffolds | 3 | |
| Genes total number | 4375 | % Total |
| Protein coding genes with predicted function | 3212 | 73.42 |
| rRNA genes | 3 | |
| tRNA genes | 49 | |
| Other RNA genes | 7 | |
| Protein coding genes with function prediction | 3212 | 73.4 |
| Without function prediction | 1084 | 24.8 |
| Biosynthetic gene clusters | 5 | |
| Genes in biosynthetic clusters | 115 | 2.6 |
| Protein coding genes coding signal peptides | 370 | 8.5 |
| Protein coding genes coding transmembrane proteins | 919 | 21.0 |
| COG clusters | 1869 | 56.4 |
| Pfam clusters | 2235 | 65.0 |
| TIGRfam clusters | 1083 | 83.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davenport, E.J.; Bose, A. Taxonomic Re-Evaluation and Genomic Comparison of Novel Extracellular Electron Uptake-Capable Rhodovulum visakhapatnamense and Rhodovulum sulfidophilum Isolates. Microorganisms 2022, 10, 1235. https://doi.org/10.3390/microorganisms10061235
Davenport EJ, Bose A. Taxonomic Re-Evaluation and Genomic Comparison of Novel Extracellular Electron Uptake-Capable Rhodovulum visakhapatnamense and Rhodovulum sulfidophilum Isolates. Microorganisms. 2022; 10(6):1235. https://doi.org/10.3390/microorganisms10061235
Chicago/Turabian StyleDavenport, Emily J., and Arpita Bose. 2022. "Taxonomic Re-Evaluation and Genomic Comparison of Novel Extracellular Electron Uptake-Capable Rhodovulum visakhapatnamense and Rhodovulum sulfidophilum Isolates" Microorganisms 10, no. 6: 1235. https://doi.org/10.3390/microorganisms10061235