
Molecular Evolution of Far-Red Light-Acclimated Photosystem II


Abstract
:1. Introduction
2. Materials and Methods
2.1. Phylogenetic Tree Construction
2.2. Homology Modeling
3. Results
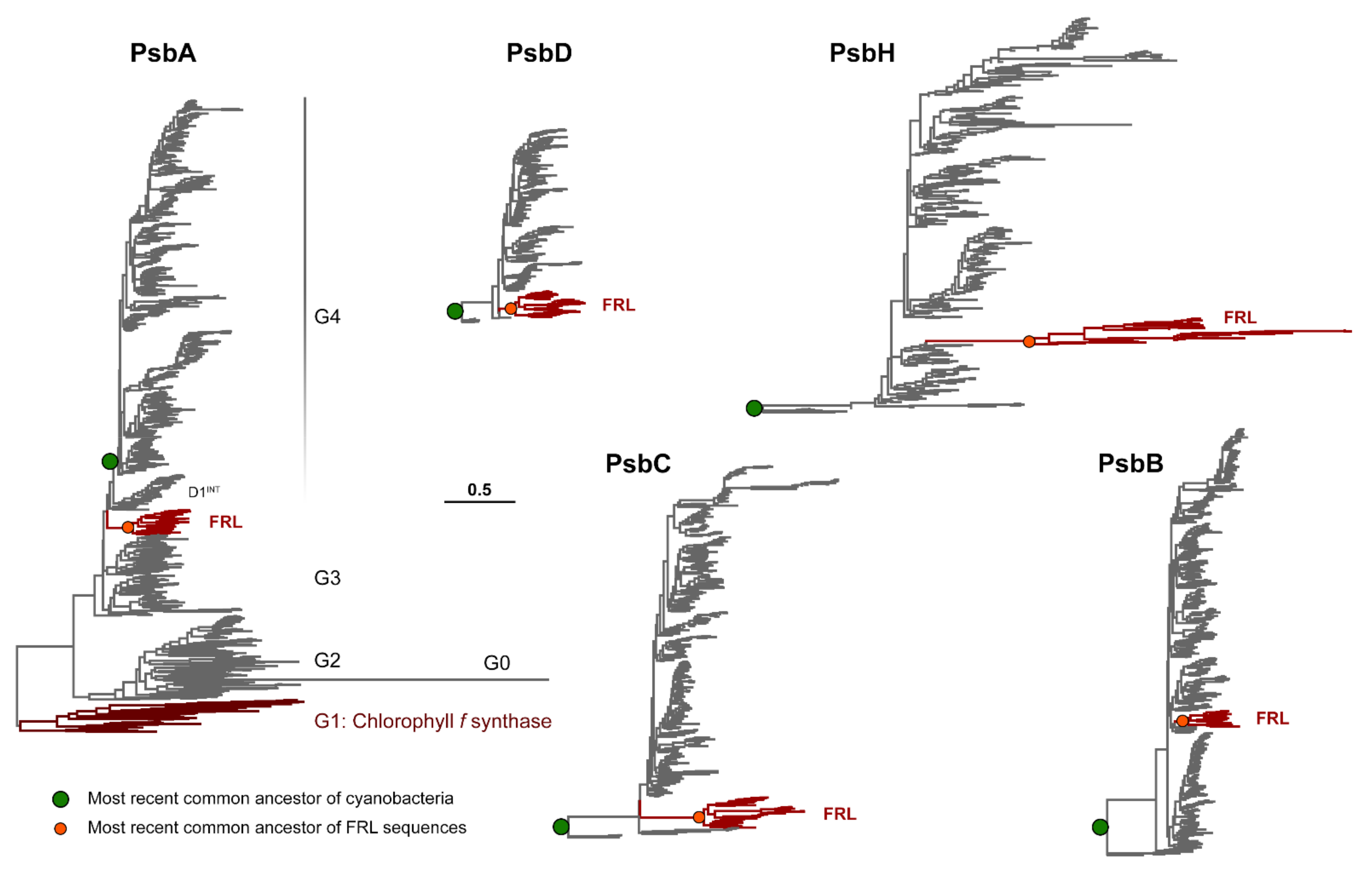
3.1. Phylogenetic Analysis
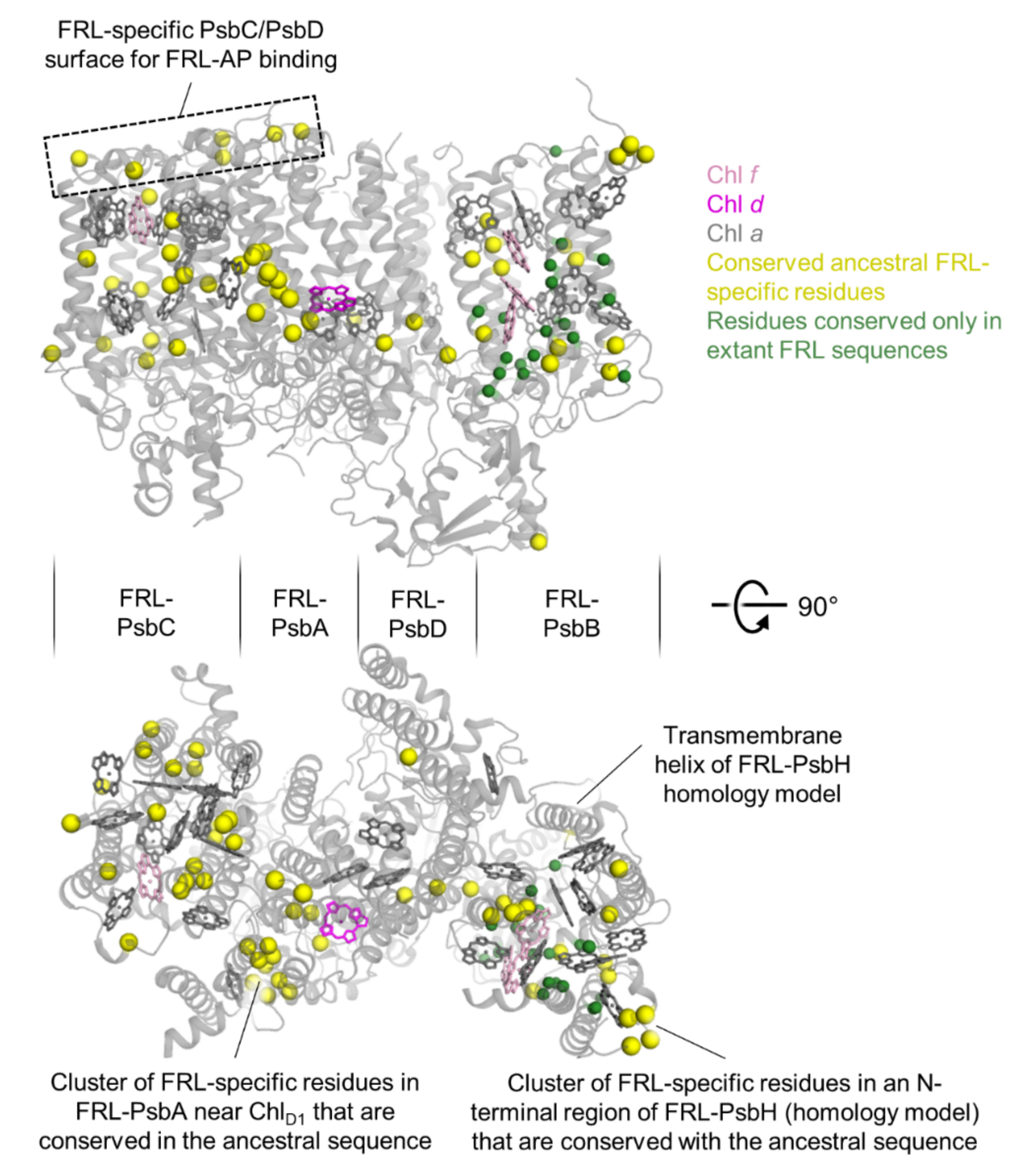
3.2. Sequence Annotations and Structural View
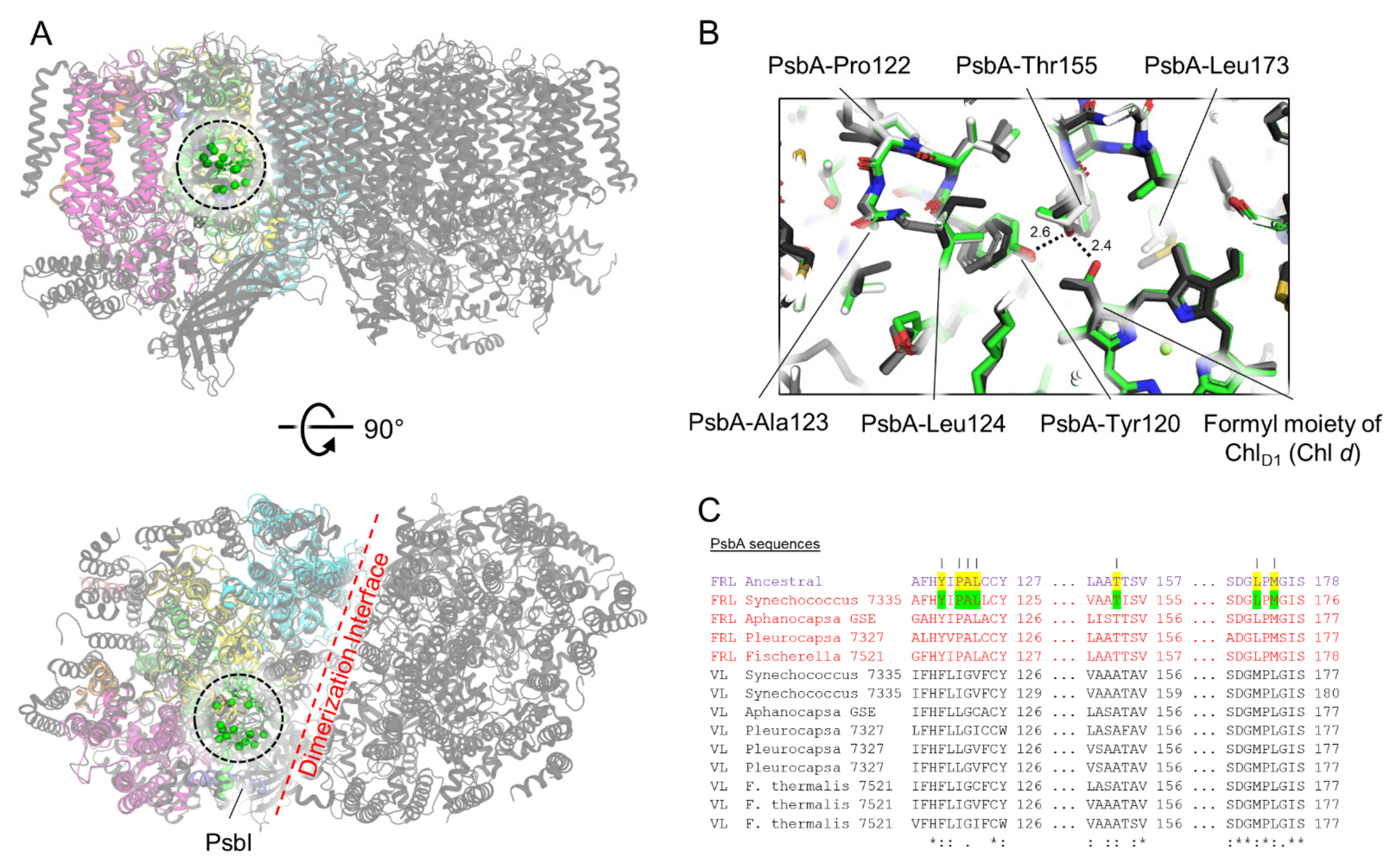
3.3. Conserved Features of FRL-PsbA
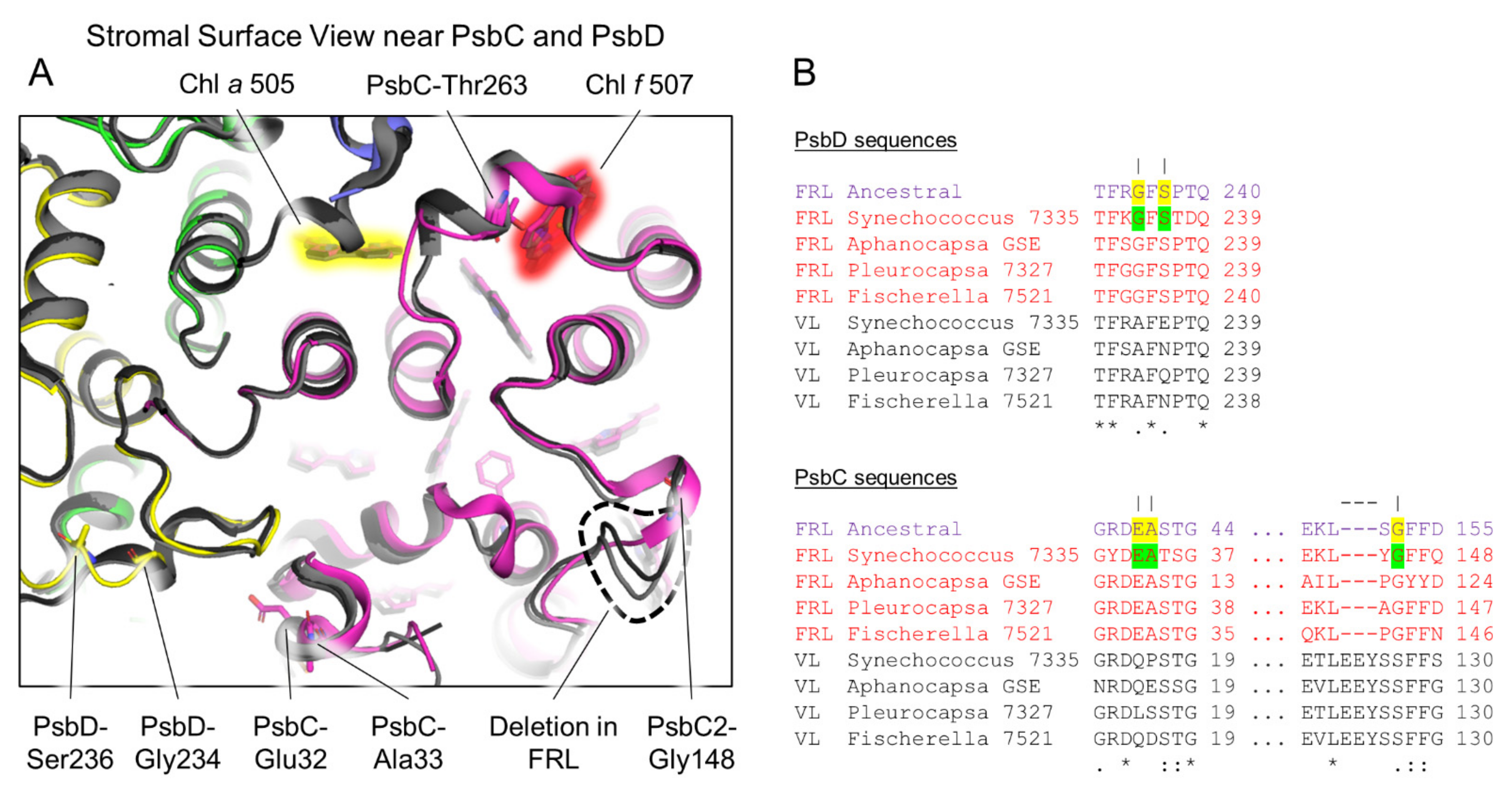
3.4. Conserved Features of FRL-PsbD
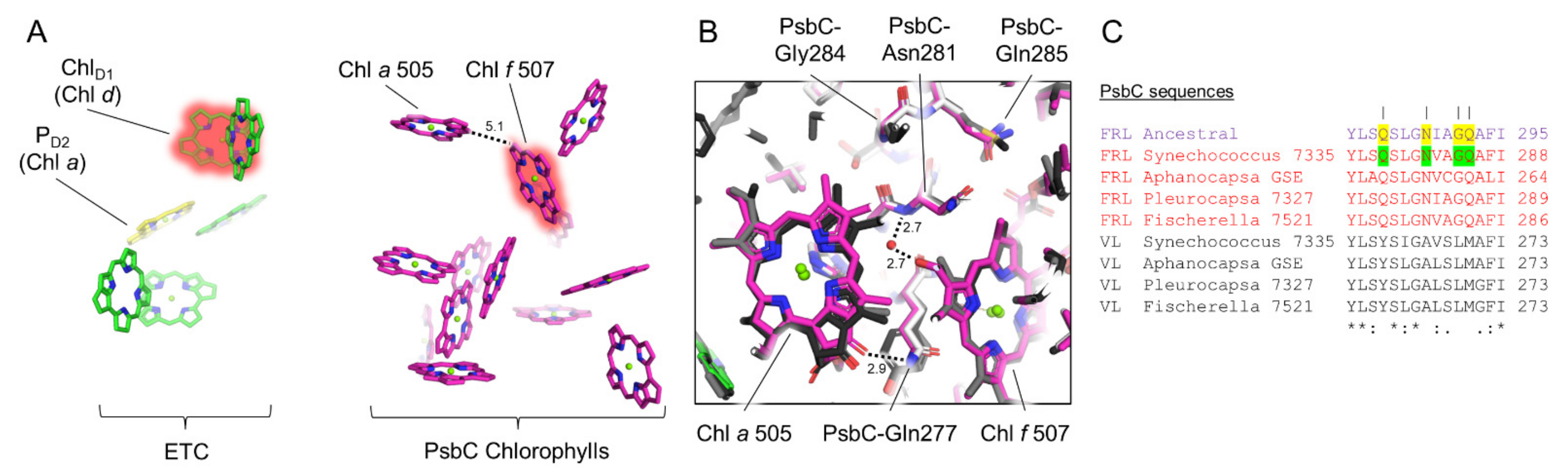
3.5. Conserved Features of FRL-PsbC
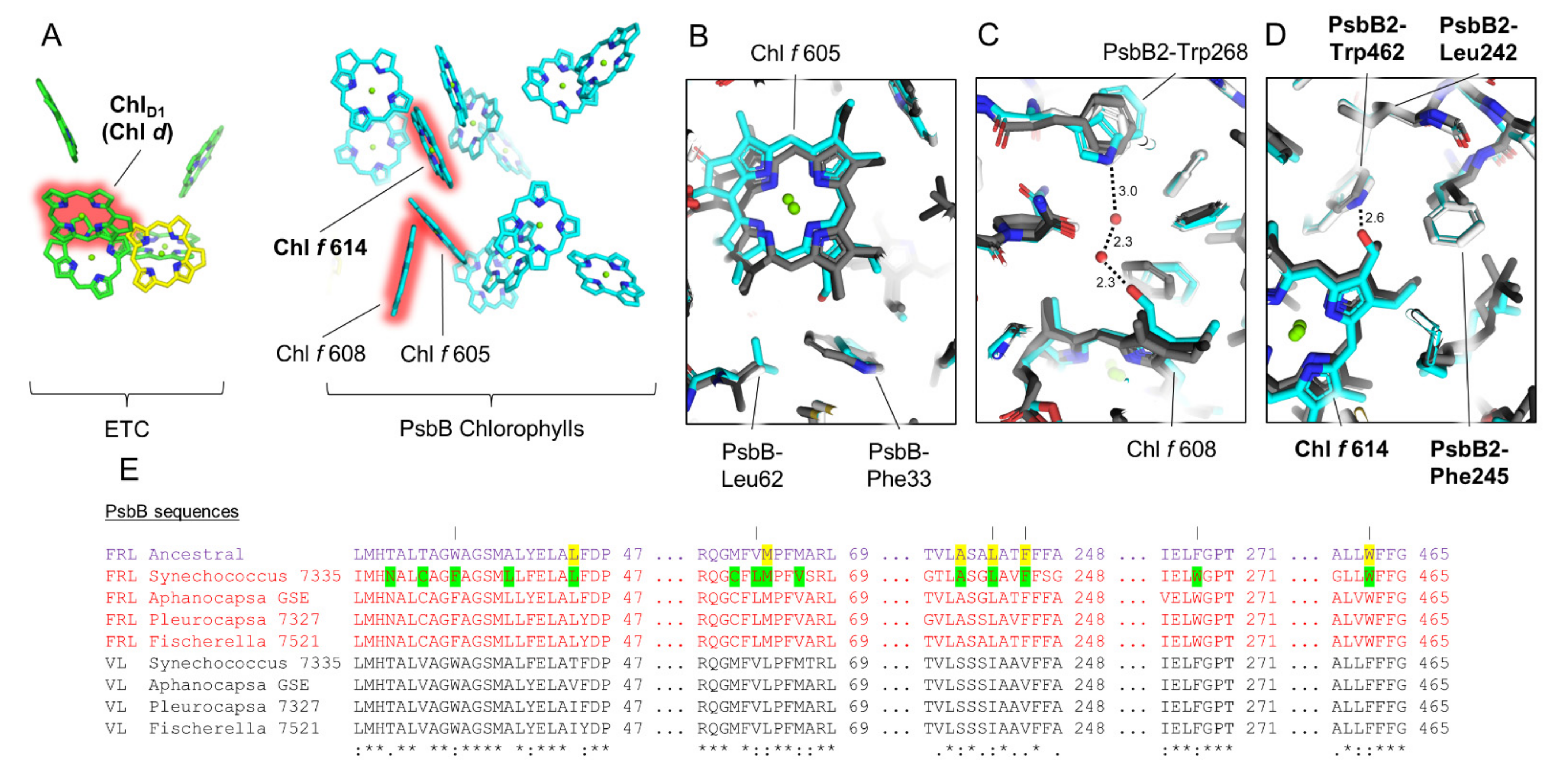
3.6. Conserved Features of FRL-PsbB
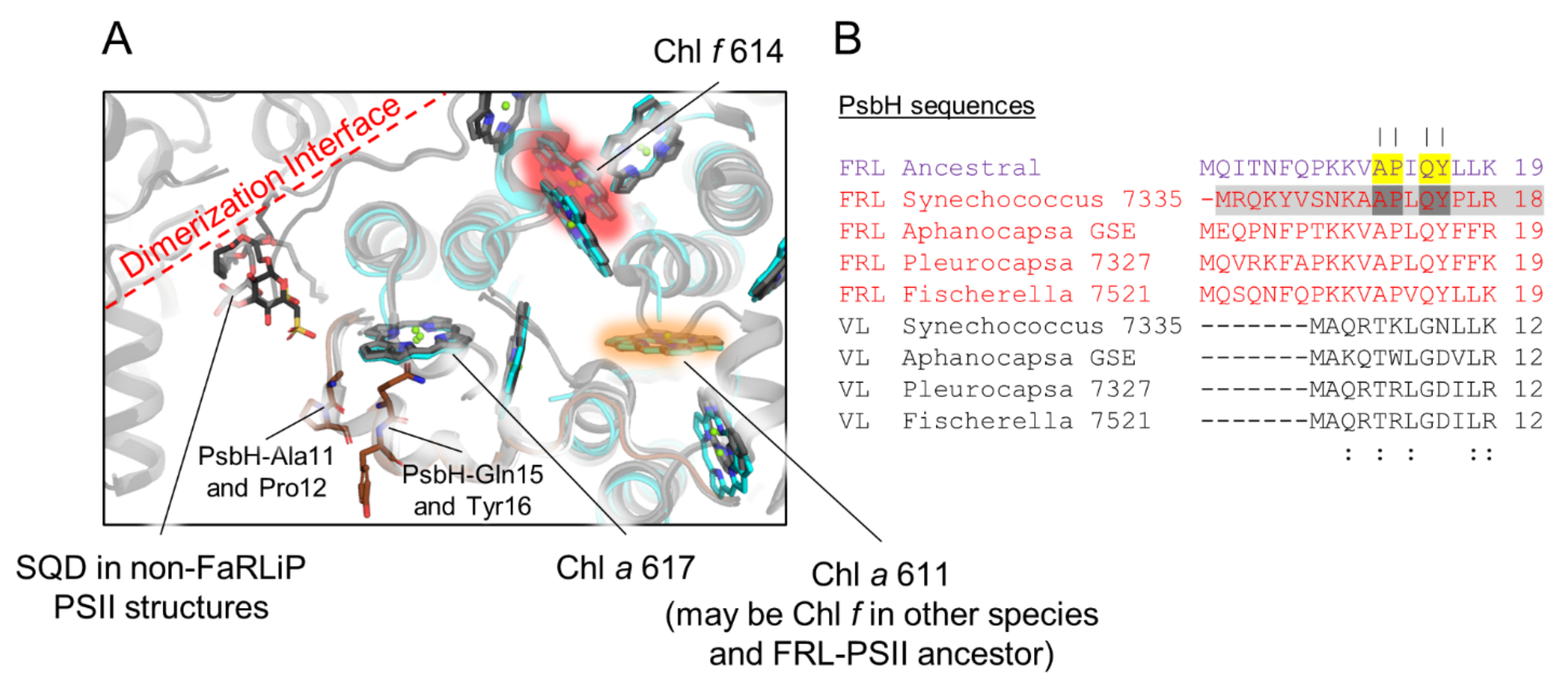
3.7. Conserved Features of FRL-PsbH
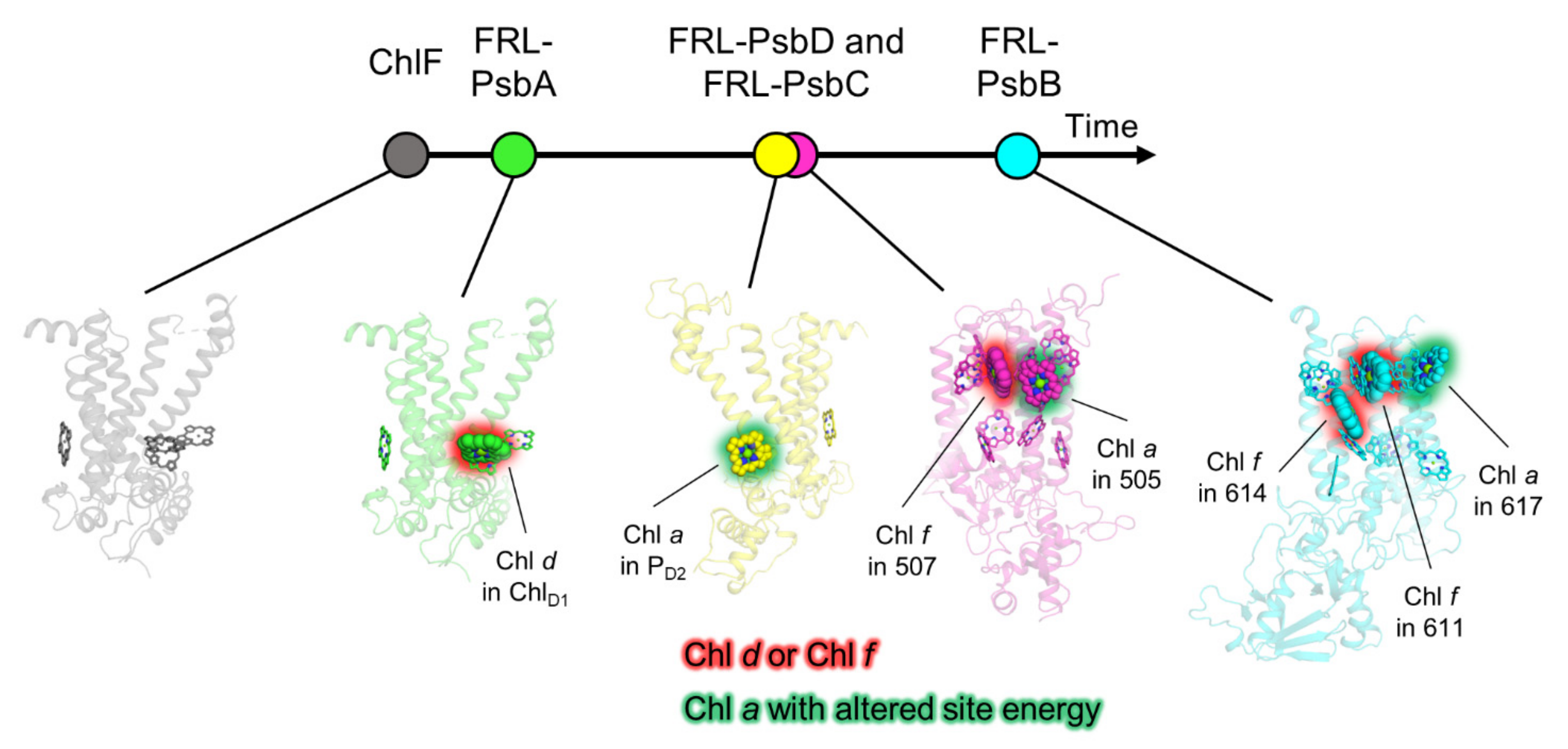
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis, 3rd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; ISBN 978-1-119-80001-9. [Google Scholar]
- Björn, L.O.; Papageorgiou, G.C.; Blankenship, R.E. Govindjee A Viewpoint: Why Chlorophyll a? Photosynth. Res. 2009, 99, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Zhang, S.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Bryant, D.A. Extensive Remodeling of a Cyanobacterial Photosynthetic Apparatus in Far-Red Light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.-Y.; Niedzwiedzki, D.M.; MacGregor-Chatwin, C.; Gerstenecker, G.; Hunter, C.N.; Blankenship, R.E.; Bryant, D.A. Extensive Remodeling of the Photosynthetic Apparatus Alters Energy Transfer among Photosynthetic Complexes When Cyanobacteria Acclimate to Far-Red Light. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148064. [Google Scholar] [CrossRef] [PubMed]
- Nürnberg, D.J.; Morton, J.; Santabarbara, S.; Telfer, A.; Joliot, P.; Antonaru, L.A.; Ruban, A.V.; Cardona, T.; Krausz, E.; Boussac, A.; et al. Photochemistry beyond the Red Limit in Chlorophyll f–Containing Photosystems. Science 2018, 360, 1210–1213. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.A.; Hunter, C.N.; Warren, M.J. Biosynthesis of the Modified Tetrapyrroles-the Pigments of Life. J. Biol. Chem. 2020, 295, 6888–6925. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Blankenship, R.E. Expanding the Solar Spectrum Used by Photosynthesis. Trends Plant Sci. 2011, 16, 427–431. [Google Scholar] [CrossRef]
- Chen, M. Chlorophyll Modifications and Their Spectral Extension in Oxygenic Photosynthesis. Annu. Rev. Biochem. 2014, 83, 317–340. [Google Scholar] [CrossRef]
- Gan, F.; Bryant, D.A. Adaptive and Acclimative Responses of Cyanobacteria to Far-Red Light. Environ. Microbiol. 2015, 17, 3450–3465. [Google Scholar] [CrossRef]
- Viola, S.; Roseby, W.; Santabarabara, S.; Nürnberg, D.; Assunção, R.; Dau, H.; Sellés, J.; Boussac, A.; Fantuzzi, A.; Rutherford, A.W. Impact of Energy Limitations on Function and Resilience in Long-Wavelength Photosystem II. bioRxiv 2022. [Google Scholar] [CrossRef]
- Wolf, B.M.; Blankenship, R.E. Far-Red Light Acclimation in Diverse Oxygenic Photosynthetic Organisms. Photosynth. Res. 2019, 142, 349–359. [Google Scholar] [CrossRef]
- Hu, C.; Nawrocki, W.J.; Croce, R. Long-Term Adaptation of Arabidopsis thaliana to Far-Red Light. Plant. Cell Environ. 2021. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Shen, G.; Bryant, D.A. Occurrence of Far-Red Light Photoacclimation (FaRLiP) in Diverse Cyanobacteria. Life 2014, 5, 4–24. [Google Scholar] [CrossRef] [PubMed]
- Antonaru, L.A.; Cardona, T.; Larkum, A.W.D.; Nürnberg, D.J. Global Distribution of a Chlorophyll f Cyanobacterial Marker. ISME J. 2020, 14, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Averina, S.; Velichko, N.; Senatskaya, E.; Pinevich, A. Far-Red Light Photoadaptations in Aquatic Cyanobacteria. Hydrobiologia 2018, 813, 1–17. [Google Scholar] [CrossRef]
- Ohkubo, S.; Miyashita, H. A Niche for Cyanobacteria Producing Chlorophyll f within a Microbial Mat. ISME J. 2017, 11, 2368–2378. [Google Scholar] [CrossRef]
- Behrendt, L.; Brejnrod, A.; Schliep, M.; Sørensen, S.J.; Larkum, A.W.D.; Kühl, M. Chlorophyll f-Driven Photosynthesis in a Cavernous Cyanobacterium. ISME J. 2015, 9, 2108–2111. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, L.; Trampe, E.L.; Nord, N.B.; Nguyen, J.; Kühl, M.; Lonco, D.; Nyarko, A.; Dhinojwala, A.; Hershey, O.S.; Barton, H. Life in the Dark: Far-Red Absorbing Cyanobacteria Extend Photic Zones Deep into Terrestrial Caves. Environ. Microbiol. 2019. [Google Scholar] [CrossRef]
- Kühl, M.; Trampe, E.; Mosshammer, M.; Johnson, M.; Larkum, A.W.D.; Frigaard, N.-U.; Koren, K. Substantial Near-Infrared Radiation-Driven Photosynthesis of Chlorophyll f-Containing Cyanobacteria in a Natural Habitat. Elife 2020, 9, e50871. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Gisriel, C.J.; Shen, G.; Ho, M.-Y.; Kurashov, V.; Flesher, D.A.; Wang, J.; Armstrong, W.H.; Golbeck, J.H.; Gunner, M.R.; Vinyard, D.J.; et al. Structure of a Monomeric Photosystem II Core Complex from a Cyanobacterium Acclimated to Far-Red Light Reveals the Functions of Chlorophylls d and f. J. Biol. Chem. 2022, 298, 101424. [Google Scholar] [CrossRef]
- Soulier, N.; Laremore, T.N.; Bryant, D.A. Characterization of Cyanobacterial Allophycocyanins Absorbing Far-Red Light. Photosynth. Res. 2020, 145, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as Designed by Its Users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 1–6. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An Interactive Tool for Rooted Phylogenetic Trees and Networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- The PyMOL Molecular Graphics System, Version 2.0 Schrödinger, LLC. Available online: https://pymol.org/2/ (accessed on 26 May 2022).
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated Comparative Protein Structure Modeling with SWISS-MODEL and Swiss-PdbViewer: A Historical Perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef]
- Gisriel, C.J.; Wang, J.; Liu, J.; Flesher, D.A.; Reiss, K.M.; Huang, H.-L.; Yang, K.R.; Armstrong, W.H.; Gunner, M.R.; Batista, V.S.; et al. High-Resolution Cryo-EM Structure of Photosystem II from the Mesophilic Cyanobacterium, Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2022, 119, e2116765118. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, K.J.; Duncan, E.J.; Eaton-Rye, J.J.; Summerfield, T.C. The Diversity and Distribution of D1 Proteins in Cyanobacteria. Photosynth. Res. 2020, 145, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T.; Murray, J.W.; Rutherford, A.W. Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria. Mol. Biol. Evol. 2015, 32, 1310–1328. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T.; Sánchez-Baracaldo, P.; Rutherford, A.W.; Larkum, A.W. Early Archean Origin of Photosystem II. Geobiology 2019, 17, 127–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, T.; Sánchez-Baracaldo, P.; Larkum, A.W.; Rutherford, A.W.; Cardona, T. Time-Resolved Comparative Molecular Evolution of Oxygenic Photosynthesis. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148400. [Google Scholar] [CrossRef]
- Ho, M.Y.; Shen, G.; Canniffe, D.P.; Zhao, C.; Bryant, D.A. Light-Dependent Chlorophyll f Synthase Is a Highly Divergent Paralog of PsbA of Photosystem II. Science 2016, 353, aaf9178. [Google Scholar] [CrossRef] [Green Version]
- Trinugroho, J.P.; Bečková, M.; Shao, S.; Yu, J.; Zhao, Z.; Murray, J.W.; Sobotka, R.; Komenda, J.; Nixon, P.J. Chlorophyll f Synthesis by a Super-Rogue Photosystem II Complex. Nat. Plants 2020, 6, 238–244. [Google Scholar] [CrossRef]
- Shen, G.; Canniffe, D.P.; Ho, M.-Y.; Kurashov, V.; van der Est, A.; Golbeck, J.H.; Bryant, D.A. Characterization of Chlorophyll f Synthase Heterologously Produced in Synechococcus Sp. PCC 7002. Photosynth. Res. 2019, 140, 77–92. [Google Scholar] [CrossRef]
- Saw, J.H.; Cardona, T.; Montejano, G. Complete Genome Sequencing of a Novel Gloeobacter Species from a Waterfall Cave in Mexico. Genome Biol. Evol. 2021, 13, evab264. [Google Scholar] [CrossRef]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla ECalteau, A.; Cai, F.; de Marsac, N.T.; Rippka, R. Improving the Coverage of the Cyanobacterial Phylum Using Diversity-Driven Genome Sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.Y.; Gan, F.; Shen, G.; Bryant, D.A. Far-Red Light Photoacclimation (FaRLiP) in Synechococcus sp. PCC 7335. II. Characterization of Phycobiliproteins Produced during Acclimation to Far-Red Light. Photosynth. Res. 2017, 131, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Llansola-Portoles, M.J.; Li, F.; Xu, P.; Streckaite, S.; Ilioaia, C.; Yang, C.; Gall, A.; Pascal, A.A.; Croce, R.; Robert, B. Tuning Antenna Function through Hydrogen Bonds to Chlorophyll a. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148078. [Google Scholar] [CrossRef]
- Umena, Y.; Kawakami, K.; Shen, J.-R.; Kamiya, N. Crystal Structure of Oxygen-Evolving Photosystem II at a Resolution of 1.9 Å. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Gisriel, C.J.; Huang, H.-L.; Reiss, K.M.; Flesher, D.A.; Batista, V.S.; Bryant, D.A.; Brudvig, G.W.; Wang, J. Quantitative Assessment of Chlorophyll Types in Cryo-EM Maps of Photosystem I Acclimated to Far-Red Light. BBA Adv. 2021, 1, 100019. [Google Scholar] [CrossRef]
- Gisriel, C.J.; Wang, J.; Brudvig, G.W.; Bryant, D.A. Opportunities and Challenges for Assigning Cofactors in Cryo-EM Density Maps of Chlorophyll-Containing Proteins. Commun. Biol. 2020, 3, 408. [Google Scholar] [CrossRef] [PubMed]
- Gisriel, C.J.; Shen, G.; Kurashov, V.; Ho, M.-Y.; Zhang, S.; Williams, D.; Golbeck, J.H.; Fromme, P.; Bryant, D.A. The Structure of Photosystem I Acclimated to Far-Red Light Illuminates an Ecologically Important Acclimation Process in Photosynthesis. Sci. Adv. 2020, 6, eaay6415. [Google Scholar] [CrossRef] [Green Version]
- Gisriel, C.J.; Flesher, D.A.; Shen, G.; Wang, J.; Ho, M.-Y.; Brudvig, G.W.; Bryant, D.A. Structure of a Photosystem I-Ferredoxin Complex from a Marine Cyanobacterium Provides Insights into Far-Red Light Photoacclimation. J. Biol. Chem. 2022, 298, 101408. [Google Scholar] [CrossRef]
- Komenda, J.; Tichý, M.; Eichacker, L.A. The PsbH Protein Is Associated with the Inner Antenna CP47 and Facilitates D1 Processing and Incorporation into PSII in the Cyanobacterium Synechocystis PCC 6803. Plant Cell Physiol. 2005, 46, 1477–1483. [Google Scholar] [CrossRef] [Green Version]
- Komenda, J.; Lupínková, L.; Kopecký, J. Absence of the PsbH Gene Product Destabilizes Photosystem II Complex and Bicarbonate Binding on Its Acceptor Side in Synechocystis PCC 6803. Eur. J. Biochem. 2002, 269, 610–619. [Google Scholar] [CrossRef] [Green Version]
- D’Haene, S.E.; Sobotka, R.; Bučinská, L.; Dekker, J.P.; Komenda, J. Interaction of the PsbH Subunit with a Chlorophyll Bound to Histidine 114 of CP47 Is Responsible for the Red 77K Fluorescence of Photosystem II. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabret, J.; Bohn, S.; Schuller, S.K.; Arnolds, O.; Möller, M.; Meier-Credo, J.; Liauw, P.; Chan, A.; Tajkhorshid, E.; Langer, J.D.; et al. Structural Insights into Photosystem II Assembly. Nat. Plants 2021, 7, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Graça, A.T.; Hall, M.; Persson, K.; Schröder, W.P. High-Resolution Model of Arabidopsis Photosystem II Reveals the Structural Consequences of Digitonin-Extraction. Sci. Rep. 2021, 11, 15534. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Hamaguchi, T.; Nakajima, Y.; Kato, K.; Kawakami, K.; Akita, F.; Yonekura, K.; Shen, J.-R. Cryo-EM Structure of Monomeric Photosystem II at 2.78 Å Resolution Reveals Factors Important for the Formation of Dimer. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148471. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Xiao, Y.; Pi, X.; Zhao, L.; Zhu, Q.; Wang, W.; Kuang, T.; Han, G.; Sui, S.-F.; Shen, J.-R. Structural Insights into a Dimeric Psb27-Photosystem II Complex from a Cyanobacterium Thermosynechococcus vulcanus. Proc. Natl. Acad. Sci. USA 2021, 118, e2018053118. [Google Scholar] [CrossRef]
- Bryant, D.A.; Shen, G.; Turner, G.M.; Soulier, N.; Laremore, T.N.; Ho, M.-Y. Far-Red Light Allophycocyanin Subunits Play a Role in Chlorophyll d Accumulation in Far-Red Light. Photosynth. Res. 2020, 143, 81–95. [Google Scholar] [CrossRef]
- Carpenter, S.D.; Charite, J.; Eggers, B.; Vermaas, W.F.J. The PsbC Start Codon in Synechocystis sp. PCC 6803. FEBS Lett. 1990, 260, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Hessa, T.; Meindl-Beinker, N.M.; Bernsel, A.; Kim, H.; Sato, Y.; Lerch-Bader, M.; Nilsson, I.; White, S.H.; von Heijne, G. Molecular Code for Transmembrane-Helix Recognition by the Sec61 Translocon. Nature 2007, 450, 1026–1030. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FRL Sequence from Synechococcus 7335 | FRL-Specific Positions Conserved in the Ancestral Sequence (%) | FRL-Specific Positions Conserved Only in the Extant Sequences (%) |
|---|---|---|
| FRL-PsbA | 3.3 | 3.3 |
| FRL-PsbB | 2.4 | 6.3 |
| FRL-PsbC | 3.9 | 3.9 |
| FRL-PsbD | 1.7 | 1.7 |
| FRL-PsbH | 6.1 | 7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisriel, C.J.; Cardona, T.; Bryant, D.A.; Brudvig, G.W. Molecular Evolution of Far-Red Light-Acclimated Photosystem II. Microorganisms 2022, 10, 1270. https://doi.org/10.3390/microorganisms10071270
Gisriel CJ, Cardona T, Bryant DA, Brudvig GW. Molecular Evolution of Far-Red Light-Acclimated Photosystem II. Microorganisms. 2022; 10(7):1270. https://doi.org/10.3390/microorganisms10071270
Chicago/Turabian StyleGisriel, Christopher J., Tanai Cardona, Donald A. Bryant, and Gary W. Brudvig. 2022. "Molecular Evolution of Far-Red Light-Acclimated Photosystem II" Microorganisms 10, no. 7: 1270. https://doi.org/10.3390/microorganisms10071270