
Proteomic Time-Course Analysis of the Filamentous Anoxygenic Phototrophic Bacterium, Chloroflexus aurantiacus, during the Transition from Respiration to Phototrophy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Growth Conditions
2.2. Protein Sample Preparations for LC-MS Proteomic Analysis
2.3. Data Analysis
2.4. Pigment Extraction and HPLC Analysis
3. Results
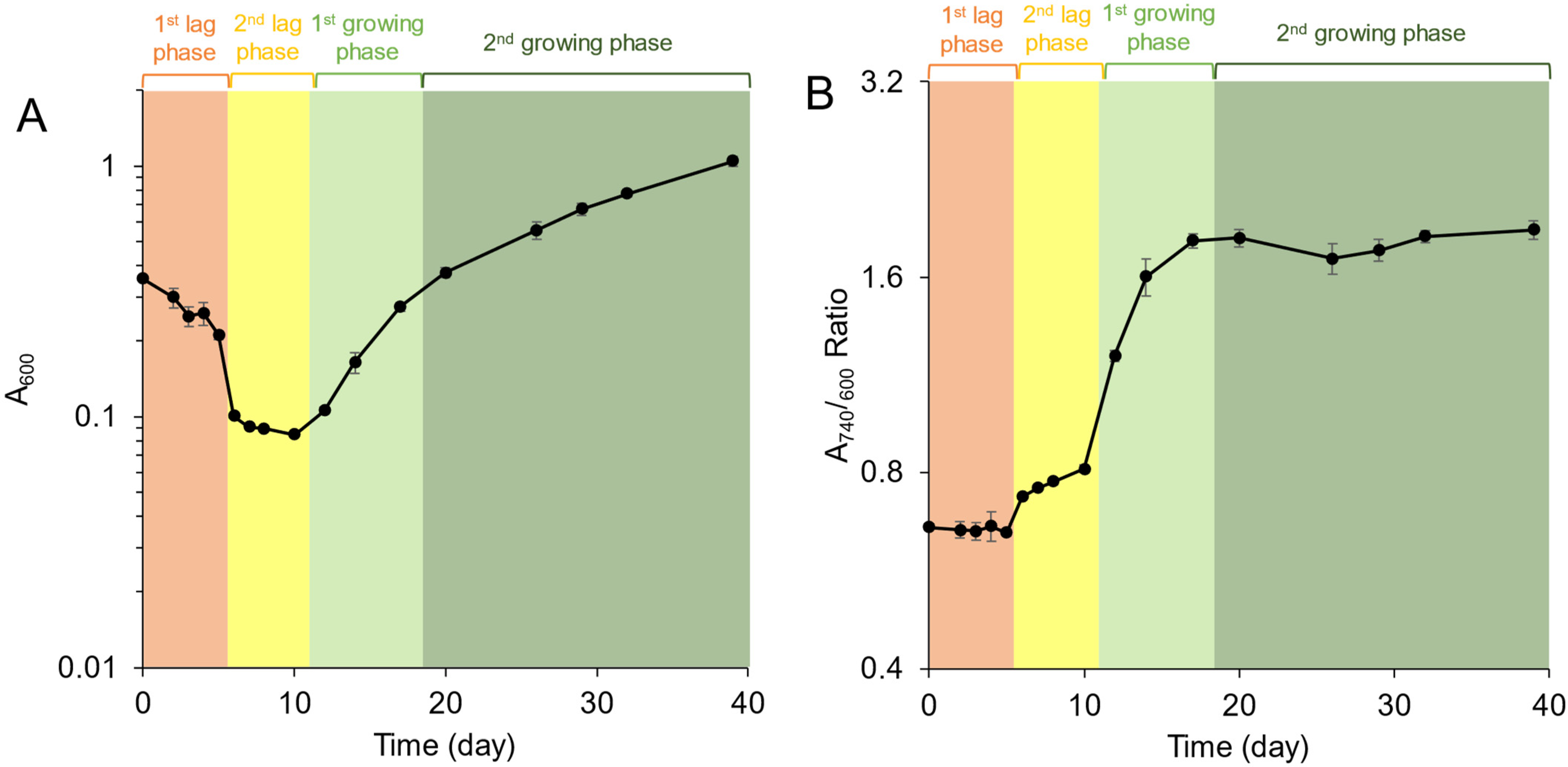
3.1. Growth Profiles
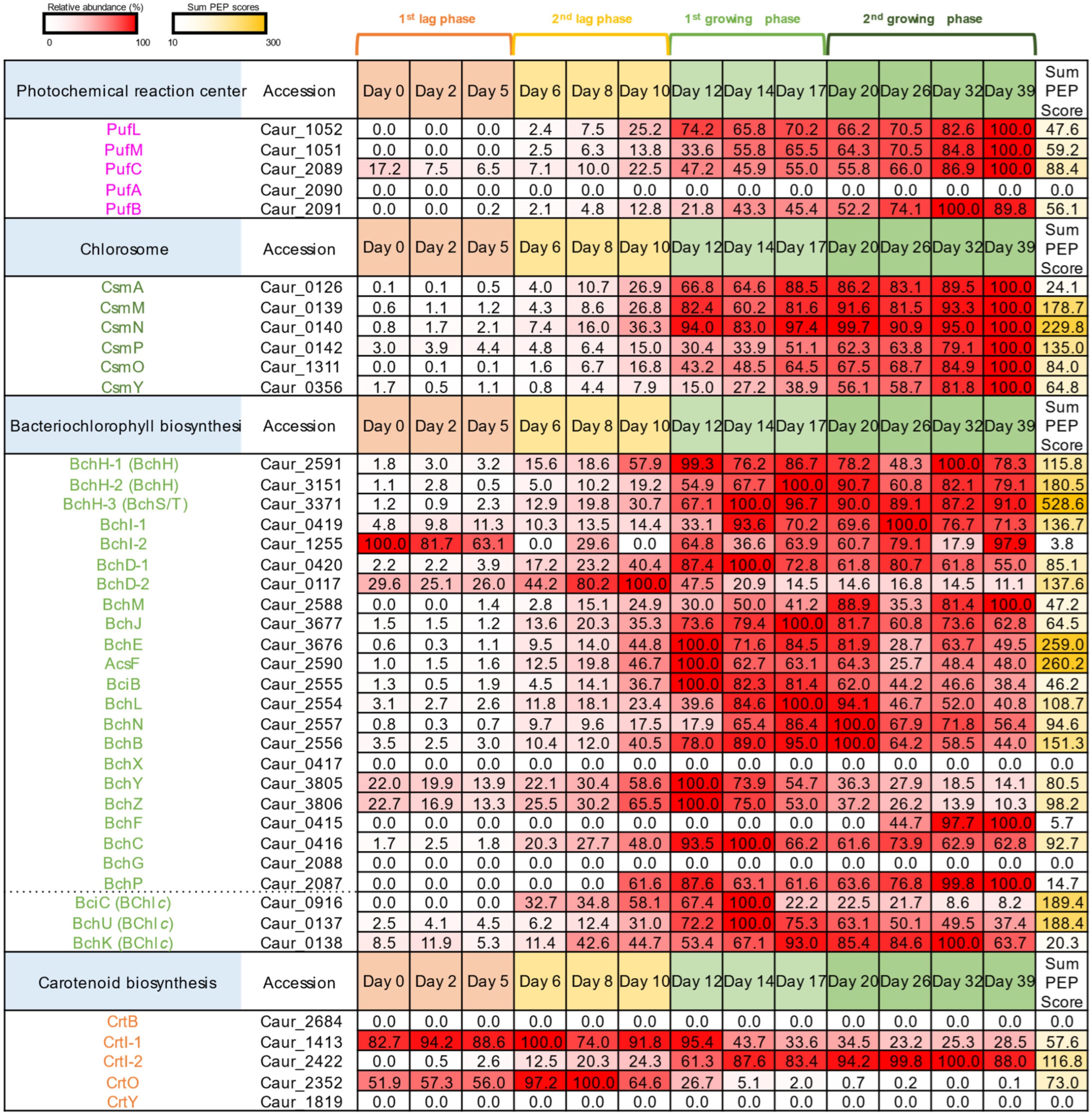
3.2. Protein Expression of Photosynthetic Proteins in C. aurantiacus
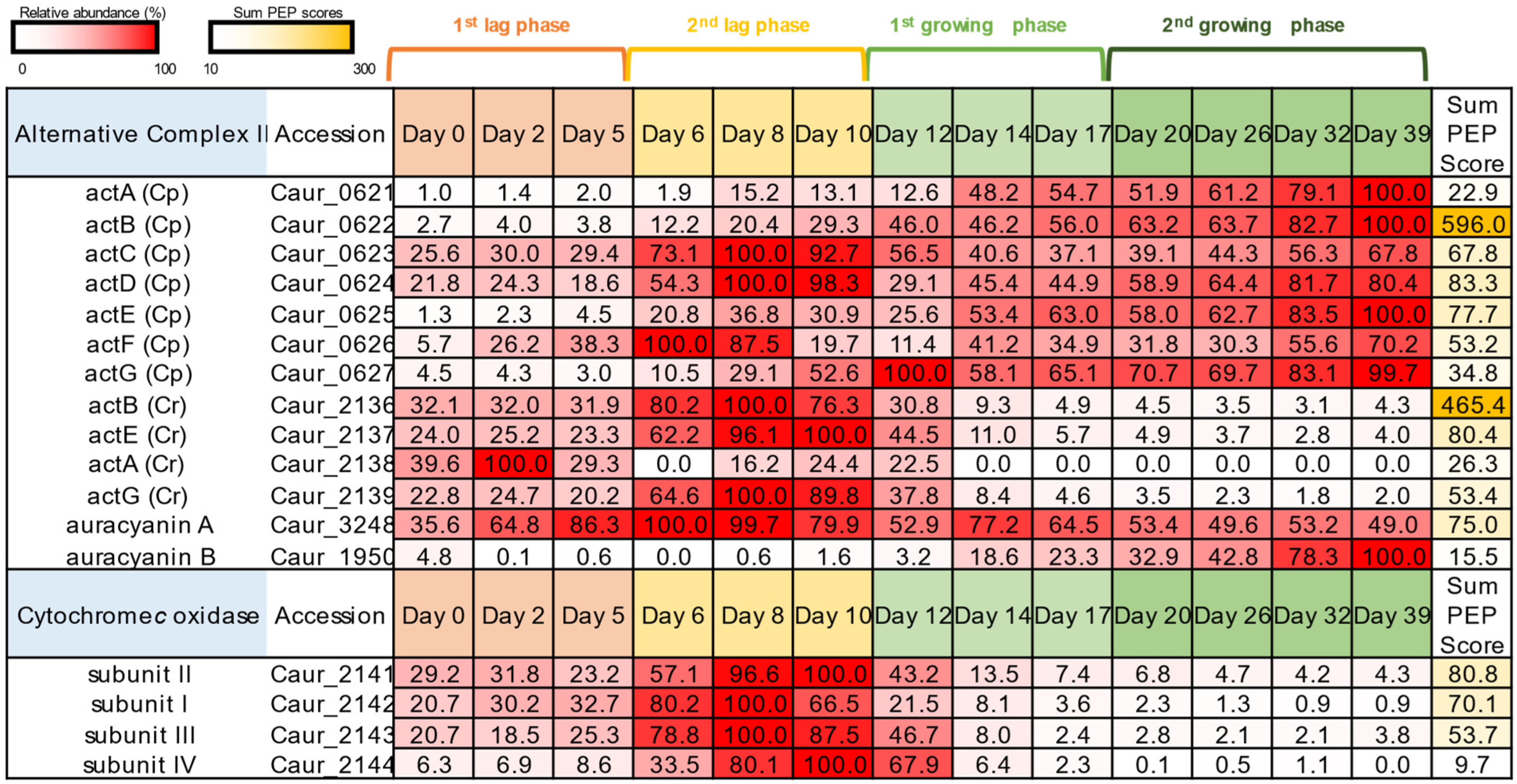
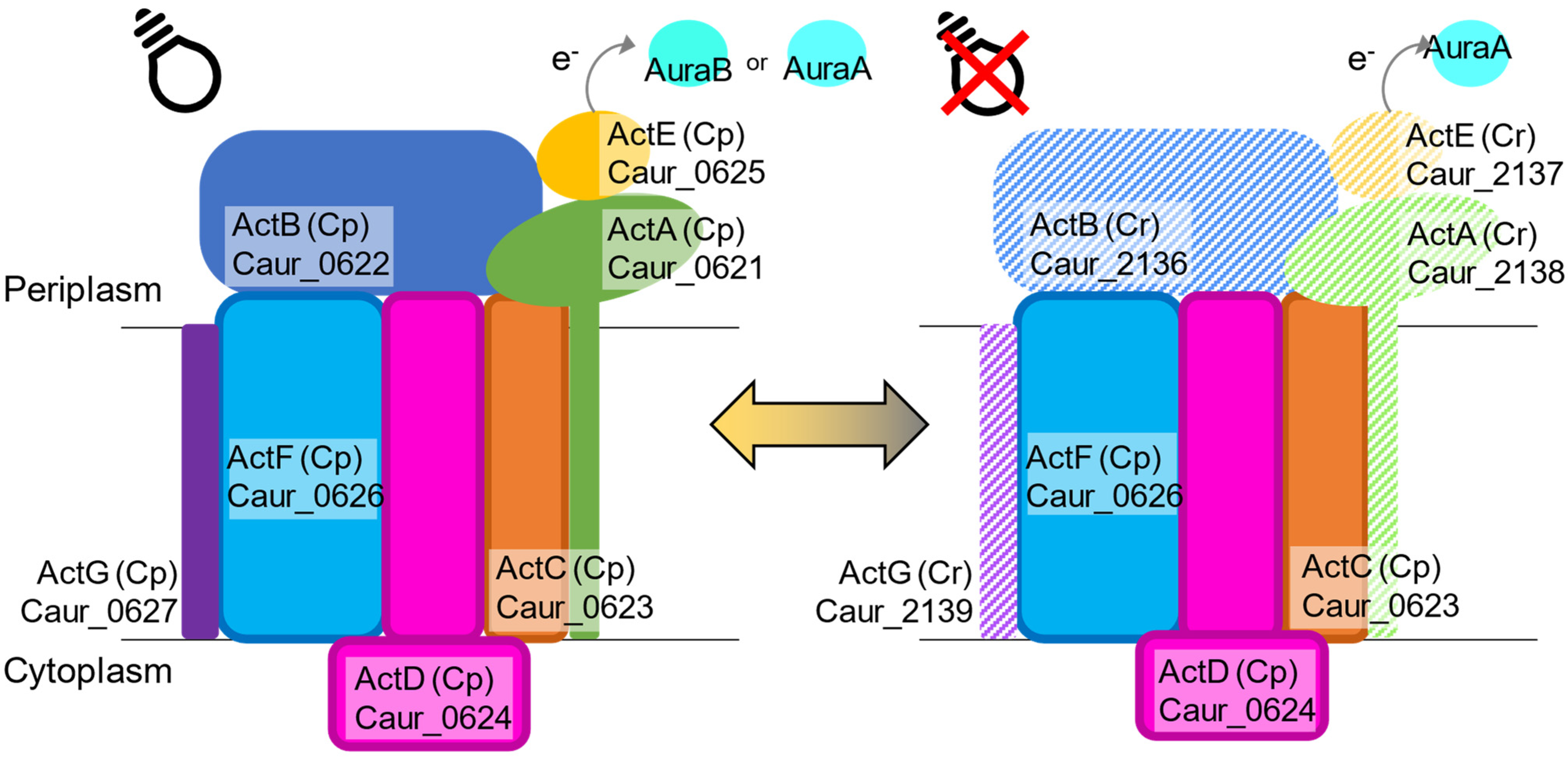
3.3. Photosynthetic and Respiratory Electron Transfer Chain
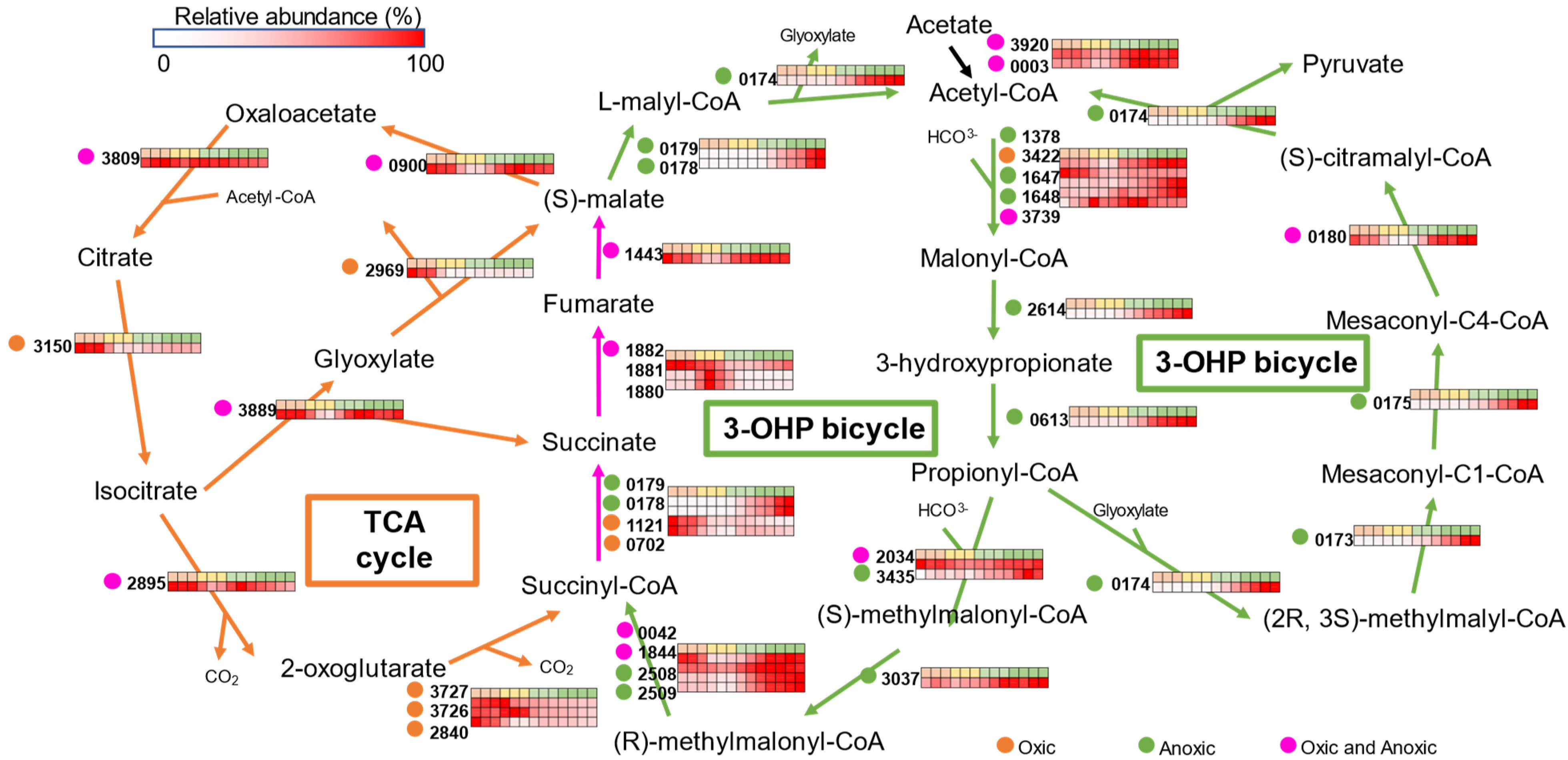
3.4. Central Carbon Metabolisms
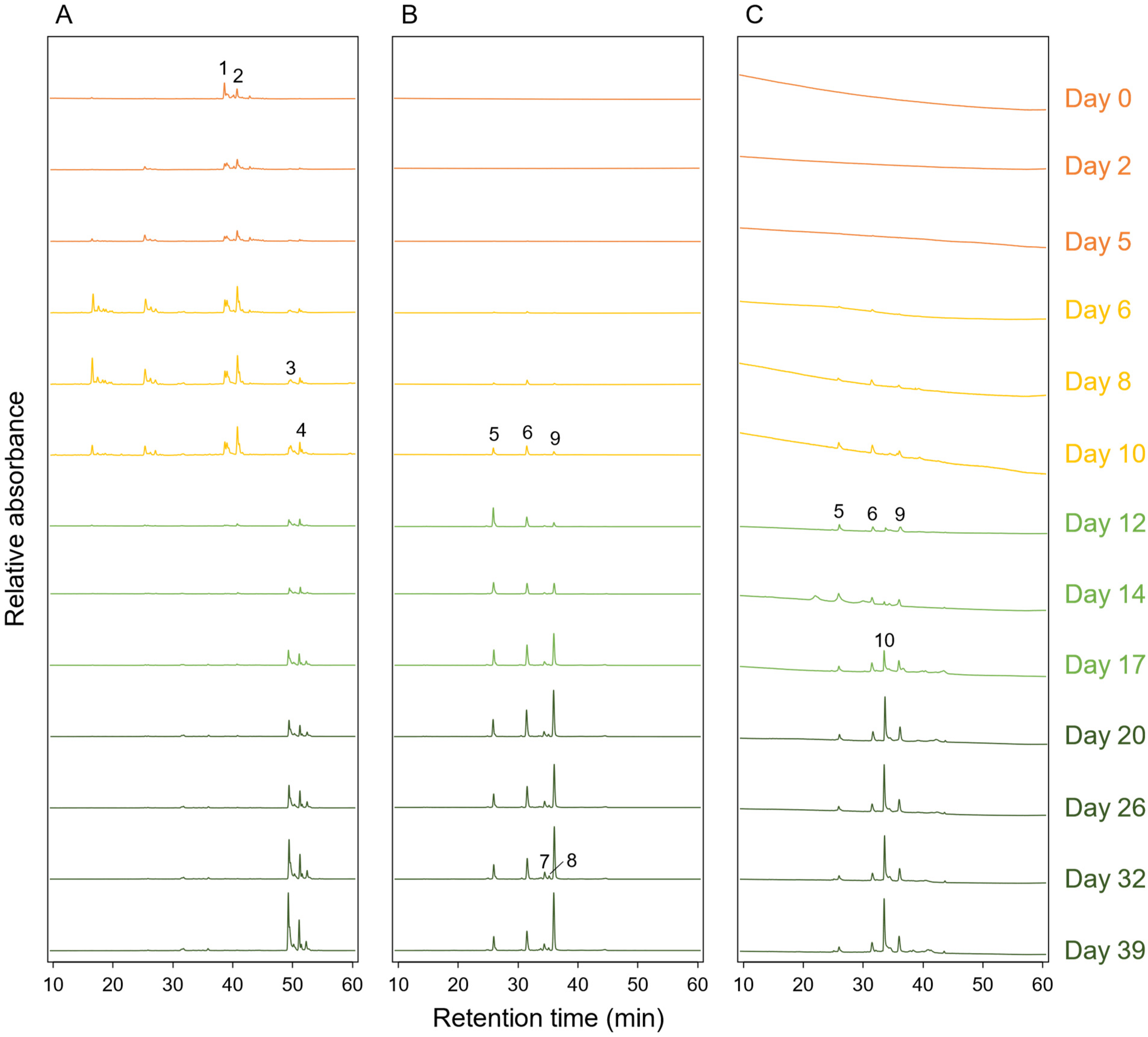
3.5. Pigment Compositions during the Cultivation
4. Discussion
5. Closing Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, B.K.; Castenholz, R.W. A phototrophic gliding filamentous bacterium of hot springs, Chloroflexus aurantiacus, gen. and sp. nov. Arch. Microbiol. 1974, 100, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T.; Petersen, S.R.; Brock, T.D. Nutritional studies on Chloroflexus, a filamentous photosynthetic, gliding bacterium. Arch. Microbiol. 1974, 100, 97–103. [Google Scholar] [CrossRef]
- Thiel, V.; Wood, J.M.; Olsen, M.T.; Tank, M.; Klatt, C.G.; Ward, D.M.; Bryant, D.A. The dark side of the Mushroom spring microbial mat: Life in the shadow of chlorophototrophs. I. Microbial diversity based on 16S rRNA gene amplicons and metagenomic sequencing. Front. Microbiol. 2016, 7, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.N.; Nishihara, A.; Lichtenberg, M.; Trampe, E.; Kawai, S.; Tank, M.; Kühl, M.; Hanada, S.; Thiel, V. Vertical distribution and diversity of phototrophic bacteria within a hot spring microbial mat (Nakabusa hot springs, Japan). Microbes Environ. 2019, 34, 374–387. [Google Scholar] [CrossRef] [Green Version]
- Klatt, C.G.; Liu, Z.; Ludwig, M.; Kühl, M.; Jensen, S.I.; Bryant, D.A.; Ward, D.M. Temporal metatranscriptomic patterning in phototrophic Chloroflexi inhabiting a microbial mat in a geothermal spring. ISME J. 2013, 7, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Kawai, S.; Martinez, J.N.; Lichtenberg, M.; Trampe, E.; Kühl, M.; Tank, M.; Haruta, S.; Nishihara, A.; Hanada, S.; Thiel, V. In-situ metatranscriptomic analyses reveal the metabolic flexibility of the thermophilic anoxygenic photosynthetic bacterium Chloroflexus aggregans in a hot spring cyanobacteria-dominated microbial mat. Microorganisms 2021, 9, 652. [Google Scholar] [CrossRef]
- Ruff-Roberts, A.L.; Kuenen, J.G.; Ward, D.M. Distribution of cultivated and uncultivated cyanobacteria and Chloroflexus-like bacteria in hot spring microbial mats. Appl. Environ. Microbiol. 1994, 60, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.M.; Ferris, M.J.; Nold, S.C.; Bateson, M.M. A natural view of microbial biodiversity within hot spring cyanobacterial mat communities. Microbiol. Mol. Biol. Rev. 1998, 62, 1353–1370. [Google Scholar] [CrossRef] [Green Version]
- Blankenship, R.E.; Madigan, M.T.; Bauer, C.E. Anoxygenic Photosynthetic Bacteria; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995. [Google Scholar]
- Xin, Y.; Shi, Y.; Niu, T.; Wang, Q.; Niu, W.; Huang, X.; Ding, W.; Yang, L.; Blankenship, R.E.; Xu, X.; et al. Cryo-EM structure of the RC-LH core complex from an early branching photosynthetic prokaryote. Nat. Commun. 2018, 9, 1568. [Google Scholar] [CrossRef] [Green Version]
- Vassilieva, E.V.; Stirewalt, V.L.; Jakobs, C.U.; Frigaard, N.-U.; Inoue-Sakamoto, K.; Baker, M.A.; Sotak, A.; Bryant, D.A. Subcellular localization of chlorosome proteins in Chlorobium tepidum and characterization of three new chlorosome proteins: CsmF, CsmH, and CsmX. Biochemistry 2002, 41, 4358–4370. [Google Scholar] [CrossRef]
- Frigaard, N.-U.; Bryant, D.A. Chlorosomes: Antenna organelles in photosynthetic green bacteria. In Complex Intracellular Structures in Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 79–114. [Google Scholar]
- Garcia Costas, A.M.; Tsukatani, Y.; Romberger, S.P.; Oostergetel, G.T.; Boekema, E.J.; Golbeck, J.H.; Bryant, D.A. Ultrastructural analysis and identification of envelope proteins of “Candidatus Chloracidobacterium thermophilum” chlorosomes. J. Bacteriol. 2011, 193, 6701–6711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukatani, Y.; Wen, J.; Blankenship, R.E.; Bryant, D.A. Characterization of the FMO protein from the aerobic chlorophototroph, Candidatus Chloracidobacterium thermophilum. Photosynth. Res. 2010, 104, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Tsukatani, Y.; Cui, W.; Zhang, H.; Gross, M.L.; Bryant, D.A.; Blankenship, R.E. Structural model and spectroscopic characteristics of the FMO antenna protein from the aerobic chlorophototroph, Candidatus Chloracidobacterium thermophilum. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauska, G.; Schoedl, T.; Remigy, H.; Tsiotis, G. The reaction center of green sulfur bacteria. Biochim. Biophys. Acta-Bioenerg. 2001, 1507, 260–277. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.M.; Cole, P.; Cox, R.; Fenna, R.; Filmer, D.; Ke, B.; Melkozernov, A.; Miller, M.; Pearlstein, R.; Raymond, J.; et al. The FMO protein. Photosynth. Res. 2004, 80, 181–187. [Google Scholar] [CrossRef]
- Chew, A.G.M.; Bryant, D.A. Chlorophyll biosynthesis in bacteria: The origins of structural and functional diversity. Annu. Rev. Microbiol. 2007, 61, 113–129. [Google Scholar] [CrossRef]
- Tamiaki, H.; Teramura, M.; Tsukatani, Y. Reduction processes in biosynthesis of chlorophyll molecules: Chemical implication of enzymatically regio- and stereoselective hydrogenations in the late stages of their biosynthetic pathway. Bull. Chem. Soc. Jpn. 2016, 89, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Yen, H.C.; Marrs, B. Map of genes for carotenoid and bacteriochlorophyll biosynthesis in Rhodopseudomonas capsulata. J. Bacteriol. 1976, 126, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, J.M.; Horne, I.M.; McEwan, A.G. Regulation of photosynthetic gene expression in purple bacteria. Microbiology 1998, 144, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.H.; Barry, K.; Chertkov, O.; Dalin, E.; Han, C.S.; Hauser, L.J.; Honchak, B.M.; Karbach, L.E.; Land, M.L.; Lapidus, A.; et al. Complete genome sequence of the filamentous anoxygenic phototrophic bacterium Chloroflexus aurantiacus. BMC Genom. 2011, 12, 334. [Google Scholar] [CrossRef] [Green Version]
- Yanyushin, M.F. Fractionation of cytochromes of phototrophically grown Chloroflexus aurantiacus. Is there a cytochrome bc complex among them? FEBS Lett. 2002, 512, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.M.; Refojo, P.N.; Hreggvidsson, G.O.; Hjorleifsdottir, S.; Teixeira, M. The alternative complex III from Rhodothermus marinus—A prototype of a new family of quinol:electron acceptor oxidoreductases. FEBS Lett. 2007, 581, 4831–4835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.M.; Carita, J.N.; Teixeira, M. Membrane-bound electron transfer chain of the thermohalophilic bacterium Rhodothermus marinus: A novel multihemic cytochrome bc, a new complex III. Biochemistry 1999, 38, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Refojo, P.N.; Sousa, F.L.; Teixeira, M.; Pereira, M.M. The alternative complex III: A different architecture using known building modules. Biochim. Biophys. Acta-Bioenerg. 2010, 1797, 1869–1876. [Google Scholar] [CrossRef] [Green Version]
- Majumder, E.L.W.; King, J.D.; Blankenship, R.E. Alternative complex III from phototrophic bacteria and its electron acceptor auracyanin. Biochim. Biophys. Acta-Bioenerg. 2013, 1827, 1383–1391. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xin, Y.; Blankenship, R.E. Enzymatic activity of the alternative complex III as a menaquinol:auracyanin oxidoreductase in the electron transfer chain of Chloroflexus aurantiacus. FEBS Lett. 2009, 583, 3275–3279. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xin, Y.; Bell, P.D.; Wen, J.; Blankenship, R.E. Structural analysis of alternative complex III in the photosynthetic electron transfer chain of Chloroflexus aurantiacus. Biochemistry 2010, 49, 6670–6679. [Google Scholar] [CrossRef] [Green Version]
- Izaki, K.; Haruta, S. Aerobic production of bacteriochlorophylls in the filamentous anoxygenic photosynthetic bacterium, Chloroflexus aurantiacus in the light. Microbes Environ. 2020, 35, ME20015. [Google Scholar] [CrossRef]
- Frigaard, N.-U.; Takaichi, S.; Hirota, M.; Shimada, K.; Matsuura, K. Quinones in chlorosomes of green sulfur bacteria and their role in the redox-dependent fluorescence studied in chlorosome-like bacteriochlorophyll c aggregates. Arch. Microbiol. 1997, 167, 343–349. [Google Scholar] [CrossRef]
- Maresca, J.A.; Graham, J.E.; Bryant, D.A. The biochemical basis for structural diversity in the carotenoids of chlorophototrophic bacteria. Photosynth. Res. 2008, 97, 121–140. [Google Scholar] [CrossRef]
- Sandmann, G. Antioxidant protection from UV-and light-stress related to carotenoid structures. Antioxidants 2019, 8, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refojo, P.N.; Teixeira, M.; Pereira, M.M. The alternative complex III: Properties and possible mechanisms for electron transfer and energy conservation. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 1852–1859. [Google Scholar] [CrossRef] [Green Version]
- Klatt, C.G.; Bryant, D.A.; Ward, D.M. Comparative genomics provides evidence for the 3-hydroxypropionate autotrophic pathway in filamentous anoxygenic phototrophic bacteria and in hot spring microbial mats. Environ. Microbiol. 2007, 9, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Hügler, M.; Sievert, S.M. Beyond the calvin cycle: Autotrophic carbon fixation in the ocean. Ann. Rev. Mar. Sci. 2011, 3, 261–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hägerhäll, C. Succinate: Quinone oxidoreductases. Biochim. Biophys. Acta-Bioenerg. 1997, 1320, 107–141. [Google Scholar] [CrossRef] [Green Version]
- Lemos, R.S.; Fernandes, A.S.; Pereira, M.M.; Gomes, C.M.; Teixeira, M. Quinol:fumarate oxidoreductases and succinate:quinone oxidoreductases: Phylogenetic relationships, metal centres and membrane attachment. Biochim. Biophys. Acta-Bioenerg. 2002, 1553, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Halfen, L.N.; Pierson, B.K.; Francis, G.W. Carotenoids of a gliding organism containing bacteriochlorophylls. Arch. Mikrobiol. 1972, 82, 240–246. [Google Scholar] [CrossRef]
- Larsen, K.L.; Miller, M.; Cox, R.P. Incorporation of exogenous long-chain alcohols into bacteriochlorophyll c homologs by Chloroflexus aurantiacus. Arch. Microbiol. 1995, 163, 119–123. [Google Scholar] [CrossRef]
- Tsukatani, Y.; Mizoguchi, T.; Thweatt, J.; Tank, M.; Bryant, D.A.; Tamiaki, H. Glycolipid analyses of light-harvesting chlorosomes from envelope protein mutants of Chlorobaculum tepidum. Photosynth. Res. 2016, 128, 235–241. [Google Scholar] [CrossRef]
- Liu, Z.; Bryant, D.A. Identification of a gene essential for the first committed step in the biosynthesis of bacteriochlorophyll c. J. Biol. Chem. 2011, 286, 22393–22402. [Google Scholar] [CrossRef] [Green Version]
- Larsen, K.L.; Cox, R.P.; Miller, M. Effects of illumination intensity on bacteriochlorophyll c homolog distribution in Chloroflexus aurantiacus grown under controlled conditions. Photosynth. Res. 1994, 41, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, R.; Mizoguchi, T.; Tamiaki, H.; Kashimura, S.; Saga, Y. Biosynthesis of unnatural bacteriochlorophyll c derivatives esterified with α,ω-diols in the green sulfur photosynthetic bacterium Chlorobaculum tepidum. Biochemistry 2011, 50, 7756–7764. [Google Scholar] [CrossRef] [PubMed]
- Orf, G.S.; Blankenship, R.E. Chlorosome antenna complexes from green photosynthetic bacteria. Photosynth. Res. 2013, 116, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Hypothesis on chlorosome biogenesis in green photosynthetic bacteria. FEBS Lett. 2007, 581, 800–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, M.O.; Linnanto, J.; Frigaard, N.-U.; Nielsen, N.C.; Miller, M. A model of the protein-pigment baseplate complex in chlorosomes of photosynthetic green bacteria. Photosynth. Res. 2010, 104, 233–243. [Google Scholar] [CrossRef]
- Tsukatani, Y.; Nakayama, N.; Shimada, K.; Mino, H.; Itoh, S.; Matsuura, K.; Hanada, S.; Nagashima, K.V.P. Characterization of a blue-copper protein, auracyanin, of the filamentous anoxygenic phototrophic bacterium Roseiflexus castenholzii. Arch. Biochem. Biophys. 2009, 490, 57–62. [Google Scholar] [CrossRef]
- Gordon, G.C.; McKinlay, J.B. Calvin cycle mutants of photoheterotrophic purple nonsulfur bacteria fail to grow due to an electron imbalance rather than toxic metabolite accumulation. J. Bacteriol. 2014, 196, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Swingley, W.D.; Blankenship, R.E.; Raymond, J. Evolutionary Relationships Among Purple Photosynthetic Bacteria and the Origin of Proteobacterial Photosynthetic Systems. In The Purple Phototrophic Bacteria. Advances in Photosynthesis and Respiration; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 17–29. [Google Scholar]
- Shih, P.M.; Ward, L.M.; Fischer, W.W. Evolution of the 3-hydroxypropionate bicycle and recent transfer of anoxygenic photosynthesis into the Chloroflexi. Proc. Natl. Acad. Sci. USA 2017, 114, 10749–10754. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.M.; Hemp, J.; Shih, P.M.; McGlynn, S.E.; Fischer, W.W. Evolution of phototrophy in the Chloroflexi phylum driven by horizontal gene transfer. Front. Microbiol. 2018, 9, 260. [Google Scholar] [CrossRef]
- Bryant, D.A.; Liu, Z.; Li, T.; Zhao, F. Comparative and functional genomics of anoxygenic green bacteria from the taxa Chlorobi, Chloroflexi, and Acidobacteria. In Functional Genomics and Evolution of Photosynthetic Systems; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2012; pp. 47–72. ISBN 978-94-007-1532-5. [Google Scholar]
- Tsuji, J.; Shaw, N.; Nagashima, S.; Venkiteswaran, J.; Schiff, S.; Hanada, S.; Tank, M.; Neufeld, J. Anoxygenic phototrophic Chloroflexota member uses a type I reaction center. bioRxiv 2020, 190934. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawai, S.; Shimamura, S.; Shimane, Y.; Tsukatani, Y. Proteomic Time-Course Analysis of the Filamentous Anoxygenic Phototrophic Bacterium, Chloroflexus aurantiacus, during the Transition from Respiration to Phototrophy. Microorganisms 2022, 10, 1288. https://doi.org/10.3390/microorganisms10071288
Kawai S, Shimamura S, Shimane Y, Tsukatani Y. Proteomic Time-Course Analysis of the Filamentous Anoxygenic Phototrophic Bacterium, Chloroflexus aurantiacus, during the Transition from Respiration to Phototrophy. Microorganisms. 2022; 10(7):1288. https://doi.org/10.3390/microorganisms10071288
Chicago/Turabian StyleKawai, Shigeru, Shigeru Shimamura, Yasuhiro Shimane, and Yusuke Tsukatani. 2022. "Proteomic Time-Course Analysis of the Filamentous Anoxygenic Phototrophic Bacterium, Chloroflexus aurantiacus, during the Transition from Respiration to Phototrophy" Microorganisms 10, no. 7: 1288. https://doi.org/10.3390/microorganisms10071288