Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. HIV Origin
3. HIV-1 Genetic Diversity
4. HIV-1 Subtypes and Recombinants
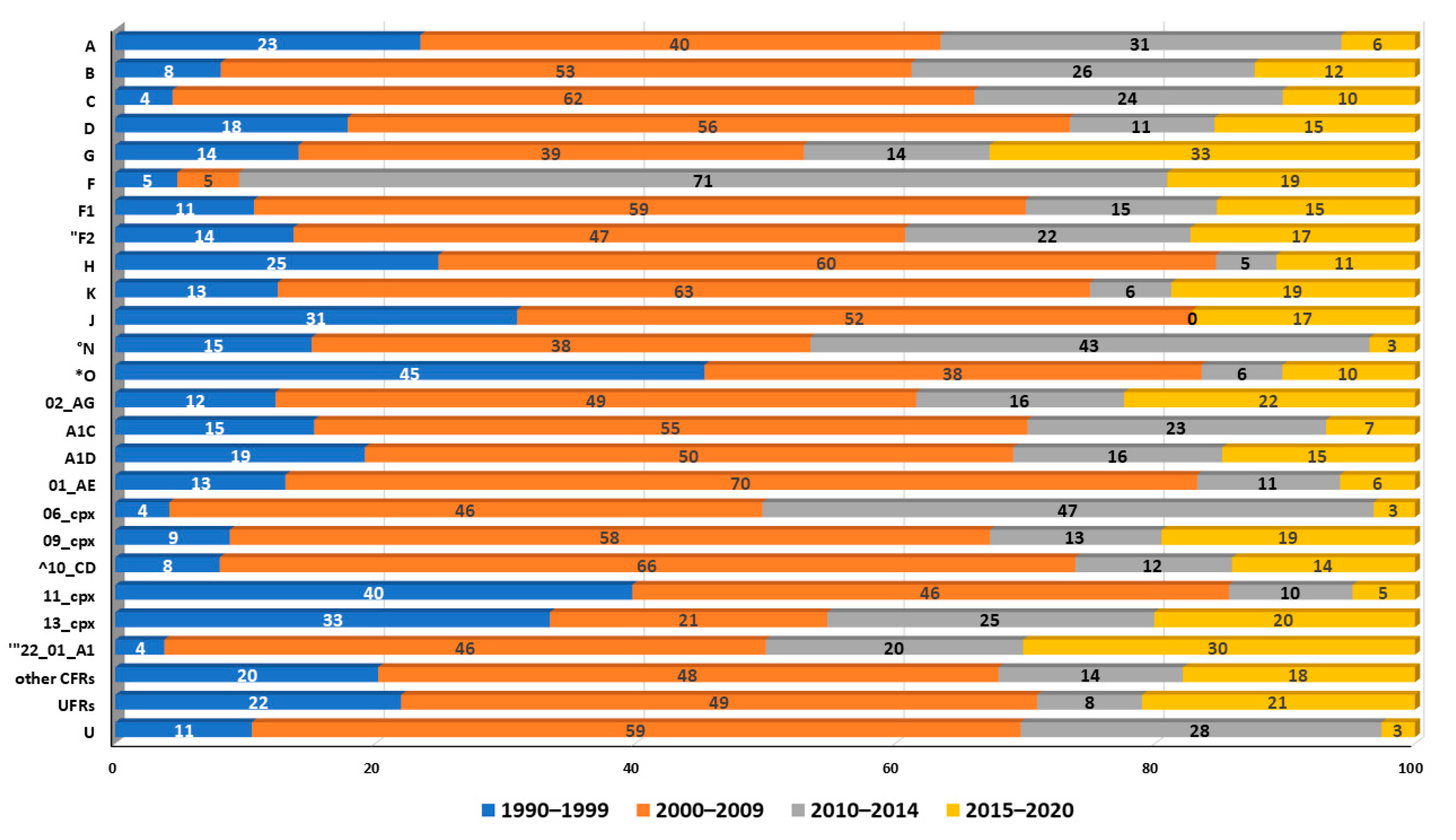
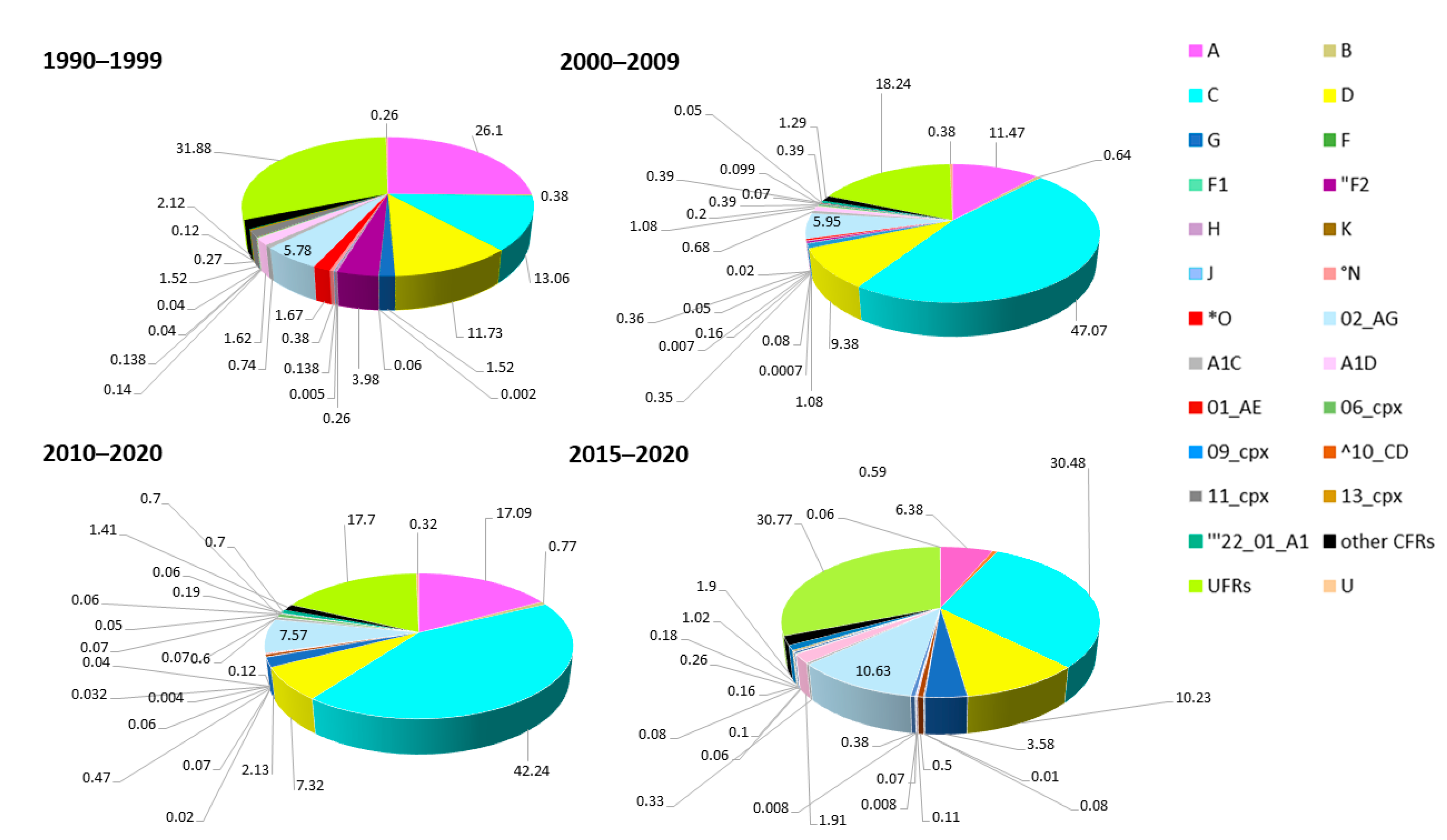
5. Distribution of Subtypes and Recombinants in African Regions
6. Summary
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Report 2020. UNAIDS Report on the Global AIDS Epidemic 2020. UNAIDS Web Site. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 10 November 2020).
- World Health Organization (WHO). Available online: https://www.afro.who.int/health-topics/hivaids (accessed on 10 November 2020).
- Rueda, S.; Mitra, S.; Chen, S.; Gogolishvili, D.; Globerman, J.; Chambers, L.; Wilson, M.; Logie, C.H.; Shi, Q.; Morassaei, S.; et al. Examining the associations between HIV-related stigma and health outcomes in people living with HIV/AIDS: A series of meta-analyses. BMJ Open 2016, 6, e011453. [Google Scholar] [CrossRef] [PubMed]
- Mabaso, M.; Makola, L.; Naidoo, I.; Mlangeni, L.L.; Jooste, S.; Simbayi, L. HIV prevalence in South Africa through gender and racial lenses: Results from the 2012 population-based national household survey. Int. J. Equity Health 2019, 18, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haal, K.; Smith, A.; van Doorslaer, E. The rise and fall of mortality inequality in South Africa in the HIV era. SSM-Popul. Health 2018, 5, 239–248. [Google Scholar]
- Maughan-Brown, B.; George, G.; Beckett, S.; Evans, M.; Lewis, L.; Cawood, C.; Khanyile, D.; Kharsany, A.B.M. HIV Risk Among Adolescent Girls and Young Women in Age-Disparate Partnerships: Evidence from KwaZulu-Natal, South Africa. J. Acquir. Immun. Defic. Syndr. 2018, 78, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Alkaiyat, A.; Weiss, M.G. HIV in the Middle East and North Africa: Priority, culture, and control. Int. J. Public Health 2013, 5, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannazadeh Baghi, H.; Soroush, M.H. HIV/AIDS in the Middle East and North Africa: A positive future. Sex. Transm. Infect. 2018, 94, 339. [Google Scholar] [CrossRef] [PubMed]
- Gebregziabher, M.; Dai, L.; Vrana-Diaz, C.; Teklehaimanot, A.; Sweat, M. Gender Disparities in Receipt of HIV Testing Results in Six Sub-Saharan African Countries. Health Equity 2018, 2, 384–394. [Google Scholar] [CrossRef] [Green Version]
- Gallo, R.C.; Sarin, P.S.; Gelmann, E.P.; Robert-Guroff, M.; Richardson, E.; Kalyanaraman, V.S.; Mann, D.; Sidhu, G.D.; Stahl, R.E.; Zolla-Pazner, S.; et al. Isolation of human T-cell leukemia virus in acquired immune deficiency syndrome (AIDS). Science 1983, 220, 865–867. [Google Scholar] [CrossRef]
- Barré-Sinoussi, F.; Chermann, J.C.; Rey, F.; Nugeyre, M.T.; Chamaret, S.; Gruest, J.; Dauguet, C.; Axler-Blin, C.; Vézinet-Brun, F.; Rouzioux, C.; et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983, 220, 868–871. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef]
- Hahn, B.H.; Shaw, G.M.; De Cock, K.M.; Sharp, P.M. AIDS as a zoonosis: Scientific and public health implications. Science 2000, 287, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Faria, N.R.; Rambaut, A.; Suchard, M.A.; Baele, G.; Bedford, T.; Ward, M.J.; Tatem, A.J.; Sousa, J.D.; Arinaminpathy, N.; Pepin, J.; et al. HIV epidemiology. The early spread and epidemic ignition of HIV-1 in human populations. Science 2014, 346, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visseaux, B.; Damond, F.; Matheron, S.; Descamps, D.; Charpentier, C. HIV-2 molecular epidemiology. Infect. Genet. Evol. 2016, 46, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.S.; Jespersen, S.; da Silva, Z.J.; Rodrigues, A.; Erikstrup, C.; Aaby, P.; Wejse, C.; Hønge, B.L. HIV-2 continues to decrease, whereas HIV-1 is stabilizing in Guinea-Bissau. AIDS 2018, 32, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Buonaguro, L.; Tornesello, M.L.; Buonaguro, F.M. Human Immunodeficiency Virus Type 1 Subtype Distribution in the Worldwide Epidemic: Pathogenetic and Therapeutic Implications. J. Virol. 2007, 81, 10209–10219. [Google Scholar] [CrossRef] [Green Version]
- Bbosa, N.; Kaleebu, P.; Ssemwanga, D. HIV subtype diversity worldwide. Curr. Opin. HIV AIDS 2019, 14, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Sharp, P.M.; McCutchan, F.E.; Hahn, B.H. Recombination in HIV-1. Nature 1995, 374, 124–126. [Google Scholar] [CrossRef]
- Delwart, E.L.; Shaper, E.G.; Louwagie, J.; McCutchan, F.E.; Grez, M.; Rübsamen-Waigmann, H.; Mullins, J.I. Genetic relationships determined by a DNA heteroduplex mobility analysis of HIV-1 env genes. Science 1993, 262, 1257–1261. [Google Scholar] [CrossRef]
- Lukashov, V.V.; Goudsmit, J. HIV heterogeneity and disease progression in AIDS: A model of continuous virus adaptation. AIDS 1998, 12 (Suppl. A), S43–S52. [Google Scholar]
- Mansky, L.M. Retrovirus mutation rates and their role in genetic variation. J. Gen. Virol. 1998, 79, 1337–1345. [Google Scholar] [CrossRef]
- Song, H.; Giorgi, E.E.; Ganusov, V.V.; Cai, F.; Athreya, G.; Yoon, H.; Carja, O.; Hora, B.; Hraber, P.; Romero-Severson, E.; et al. Tracking HIV-1 recombination to resolve its contribution to HIV-1 evolution in natural infection. Nat. Commun. 2018, 9, 1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korber, B.; Gaschen, B.; Yusim, K.; Thakallapally, R.; Kesmir, C.; Detours, V. Evolutionary and immunological implications of contemporary HIV-1 variation. Br. Med. Bull. 2001, 58, 19–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peteers, M. Recombinant HIV Sequences: Their Role in the Global Epidemic. HIV Sequence Compendium 2000; pp. 54–72. Available online: https://www.hiv.lanl.gov/content/immunology/pdf/2000/1/Peeters.pdf (accessed on 10 November 2020).
- Grobler, J.; Gray, C.M.; Rademeyer, C.; Seoighe, C.; Ramjee, G.; Karim, S.A.; Morris, L.; Williamson, C. Incidence of HIV-1 dual infection and its association with increased viral load set point in a cohort of HIV-1 subtype C-infected female sex workers. J. Infect. Dis. 2004, 190, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Ssemwanga, D.; Lyagoba, F.; Ndembi, N.; Mayanja, B.N.; Larke, N.; Wang, S.; Baalwa, J.; Williamson, C.; Grosskurth, H.; Kaleebu, P. Multiple HIV-1 infections with evidence of recombination in heterosexual partnerships in a low risk rural clinical cohort in Uganda. Virology 2011, 411, 113–131. [Google Scholar] [CrossRef] [Green Version]
- Robertson, D.L.; Anderson, J.P.; Bradac, J.A.; Carr, J.K.; Foley, B.; Funkhouser, R.K.; Gao, F.; Hahn, B.H.; Kalish, M.L.; Kuiken, C.; et al. HIV-1 nomenclature proposal. Science 2000, 288, 55–56. [Google Scholar] [CrossRef]
- Simon, F.; Mauclère, P.; Roques, P.; Loussert-Ajaka, I.; Müller-Trutwin, M.C.; Saragosti, S.; Georges-Courbot, M.C.; Barré-Sinoussi, F.; Brun-Vézinet, F. Identification of a new human immunodeficiency virus type 1 distinct from group M and group O. Nat. Med. 1998, 4, 1032–1037. [Google Scholar] [CrossRef]
- Vallari, A.; Holzmayer, V.; Harris, B.; Yamaguchi, J.; Ngan- sop, C.; Makamche, F.; Mbanya, D.; Kaptué, L.; Ndembi, N.; Gürtler, L.; et al. Confirmation of putative HIV-1 group P in Cameroon. J. Virol. 2011, 85, 1403–1407. [Google Scholar] [CrossRef] [Green Version]
- Désiré, N.; Cerutti, L.; Le Hingrat, Q.; Perrier, M.; Emler, S.; Calvez, V.; Descamps, D.; Marcelin, A.G.; Hué, S.; Visseaux, B. Characterization update of HIV-1 M subtypes diversity and proposal for subtypes A and D sub-subtypes reclassification. Retrovirology 2018, 15, 80. [Google Scholar] [CrossRef] [Green Version]
- Triques, K.; Bourgeois, A.; Vidal, N.; Mpoudi-Ngole, E.; Mulanga-Kabeya, C.; Nzilambi, N.; Torimiro, N.; Saman, E.; Delaporte, E.; Peeters, M. Near-full-length genome sequencing of divergent African HIV type 1 subtype F viruses leads to the identification of a new HIV type 1 subtype designated K. AIDS Res. Hum. Retroviruses 2000, 16, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, J.; McArthur, C.; Vallari, A.; Sthreshley, L.; Cloherty, G.A.; Berg, M.G.; Rodgers, M.A. Complete genome sequence of CG-0018a-01 establishes HIV-1 subtype L. J. Acquir. Immun. Defic. Syndr. 2020, 83, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Trask, S.A.; Hui, H.; Mamaeva, O.; Chen, Y.; Theodore, T.S.; Foley, B.T.; Korber, B.T.; Shaw, G.M.; Hahn, B.H. Molecular characterization of a highly divergent HIV type 1 isolate obtained early in the AIDS epidemic from the Democratic Republic of Congo. AIDS Res. Hum. Retroviruses 2001, 17, 1217–1222. [Google Scholar] [CrossRef] [PubMed]
- Los Alamos National Laboratory. HIV Circulating Recombinant Forms (CRFs). Available online: http://www.hiv.lanl.gov/content/sequence/HIV/CRFs/CRFs.html (accessed on 25 November 2020).
- Zhou, J.; Li, M.; Chen, M.; Ma, Y.; Shao, Y.; Xing, H. Near Full-Length Genomic Characterization of a Novel HIV-1 Circulating Recombinant Form (CRF106_cpx) Identified among Heterosexuals in China. AIDS Res. Hum. Retroviruses 2020, 36, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Janssens, W.; Salminen, M.O.; Laukkanen, T.; Heyndrickx, L.; van der Auwera, C.R.; McCutchan, F.E.; van der Groen, G. Near full-length genome analysis of HIV type 1 CRF02. AG subtype C and CRF02. AG subtype G recombinants. AIDS Res. Hum. Retroviruses 2000, 16, 1183–1189. [Google Scholar] [CrossRef]
- Oelrichs, R.B.; Workman, C.; Laukkanen, T.; McCutchan, F.E.; Deacon, N.J. A novel subtype A/G/J recombinant full-length HIV type 1 genome from Burkina Faso. AIDS Res. Hum. Retroviruses 1998, 14, 1495–1500. [Google Scholar] [CrossRef]
- Vidal, N.; Peeters, M.; Mulanga-Kabeya, C.; Nzilamb, N.; Robertson, D.; Ilunga, W.; Sema, H.; Tshimanga, K.; Bongo, B.; Delaporte, E. Unprecedented degree of human immunodeficiency virus type 1 (HIV-1) group M genetic diversity in the Democratic Republic of Congo suggests that the HIV-1 pandemic originated in Central Africa. J. Virol. 2000, 74, 10498–10507. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Li, L.; Li, H.; Liu, S.; Wang, X.; Bao, Z.; Li, T.; Zhuang, D.; Liu, Y.; Li, J. Recombination pattern reanalysis of some HIV-1 circulating recombination forms suggest the necessity and difficulty of revision. PLoS ONE 2014, 9, e107349. [Google Scholar] [CrossRef]
- Taylor, B.S.; Hammer, S.M. The challenge of HIV-1 subtype diversity. N. Engl. J. Med. 2008, 12, 1590–1602. [Google Scholar] [CrossRef] [Green Version]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D. WHO–UNAIDS Networ k for HIV Isolation Global and regional molecular epidemiology of HIV-1, 1990–2015: A systematic review, global survey, and trend analysis. Lancet Infect. Dis. 2019, 19, 143–155. [Google Scholar] [CrossRef]
- Gartner, M.J.; Roche, M.; Churchill, M.J.; Paul, R.; Gorry, P.R.; Flynn, J.K. Understanding the mechanisms driving the spread of subtype C HIV-1. EBioMedicine 2020, 53, 102682. [Google Scholar] [CrossRef]
- Tatem, A.J.; Hemelaar, J.; Gray, R.R.; Salemi, M. Spatial accessibility and the spread of HIV-1 subtypes and recombinants in sub-Saharan Africa. AIDS 2012, 26, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Paraskevis, D.; Nikolopoulos, G.K.; Magiorkinis, G.; Hodges-Mameletzis, I.; Hatzakis, A. The application of HIV molecular epidemiology to public health. Infect. Genet. Evol. 2016, 46, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Lihana, R.W.; Ssemwanga, D.; Abimiku, A.; Ndembi, N. Update on HIV-1 diversity in Africa: A decade in review. AIDS Rev. 2012, 14, 83–100. [Google Scholar] [PubMed]
- Venner, C.M.; Nankya, I.; Kyeyune, F.; Demers, K.; Kwok, C.; Chen, P.L.; Rwambuya, S.; Munjoma, M.; Chipato, T.; Byamugisha, J.; et al. Infecting HIV-1 subtype predicts disease progression in women of Sub-Saharan Africa. EBioMedicine 2016, 13, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, M.; Zhang, W.; Zhang, X.; Jiang, W.; Huan, X.; Meng, X.; Zhu, B.; Yang, Y.; Tao, Y.; Tian, T.; et al. HIV-1 CRF01_AE strain is associated with faster HIV/AIDS progression in Jiangsu Province, China. Sci. Rep. 2017, 7, 1570. [Google Scholar] [CrossRef] [PubMed]
- Pillay, D.; Herbeck, J.; Cohen, M.S.; de Oliveira, T.; Fraser, C.; Ratmann, O.; Brown, A.L.; Kellam, P.; PANGEA-HIV Consortium. PANGEA-HIV: Phylogenetics for generalised epidemics in Africa. Lancet Infect. Dis. 2015, 15, 259–261. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, A.; Farcomeni, S.; Fontanelli Sulekova, L.; Grieco, S.; Sernicola, L.; Baesso, M.; Maggiorella, M.T.; Angeletti, S.; Foley, B.; Ciccozzi, M.; et al. Characterization of HIV-1 subtypes among South Sudanese patients. AIDS Res. Hum. Retroviruses 2019, 35, 968–971. [Google Scholar] [CrossRef]
- Hemelaar, J. The origin and diversity of the HIV-1 pandemic. Trends Mol. Med. 2012, 18, 182–192. [Google Scholar] [CrossRef]
- McCutchan, F.E. Understanding the genetic diversity of HIV-1. AIDS 2000, 14, S31–S44. [Google Scholar]
- Hemelaar, J.; Loganathan, S.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Kirtley, S. WHO-UNAIDS Network for HIV Country-level diversity of the HIV-1 pandemic between 1990 and 2015. J. Virol. 2020, 21, JVI.01580-20. [Google Scholar] [CrossRef]
- Hentz, J.M. The Impact of HIV on the Rape Crisis in the African Great Lakes Region. Hum. Rights Brief. 2005, 12, 12–15. [Google Scholar]
- TenoRes Study Group. Global epidemiology of drug resistance after failure of WHO recommended first-line regimens for adult HIV-1 infection: A multicenter retrospective cohort study. Lancet Infect. Dis. 2016, 16, 565–575. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovanetti, M.; Ciccozzi, M.; Parolin, C.; Borsetti, A. Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview. Pathogens 2020, 9, 1072. https://doi.org/10.3390/pathogens9121072
Giovanetti M, Ciccozzi M, Parolin C, Borsetti A. Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview. Pathogens. 2020; 9(12):1072. https://doi.org/10.3390/pathogens9121072
Chicago/Turabian StyleGiovanetti, Marta, Massimo Ciccozzi, Cristina Parolin, and Alessandra Borsetti. 2020. "Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview" Pathogens 9, no. 12: 1072. https://doi.org/10.3390/pathogens9121072