Multiple Antigenic Peptide-Based Vaccines Targeting Ixodes ricinus Neuropeptides Induce a Specific Antibody Response but Do Not Impact Tick Infestation

 , ,
, ,
Abstract
:1. Introduction
2. Results
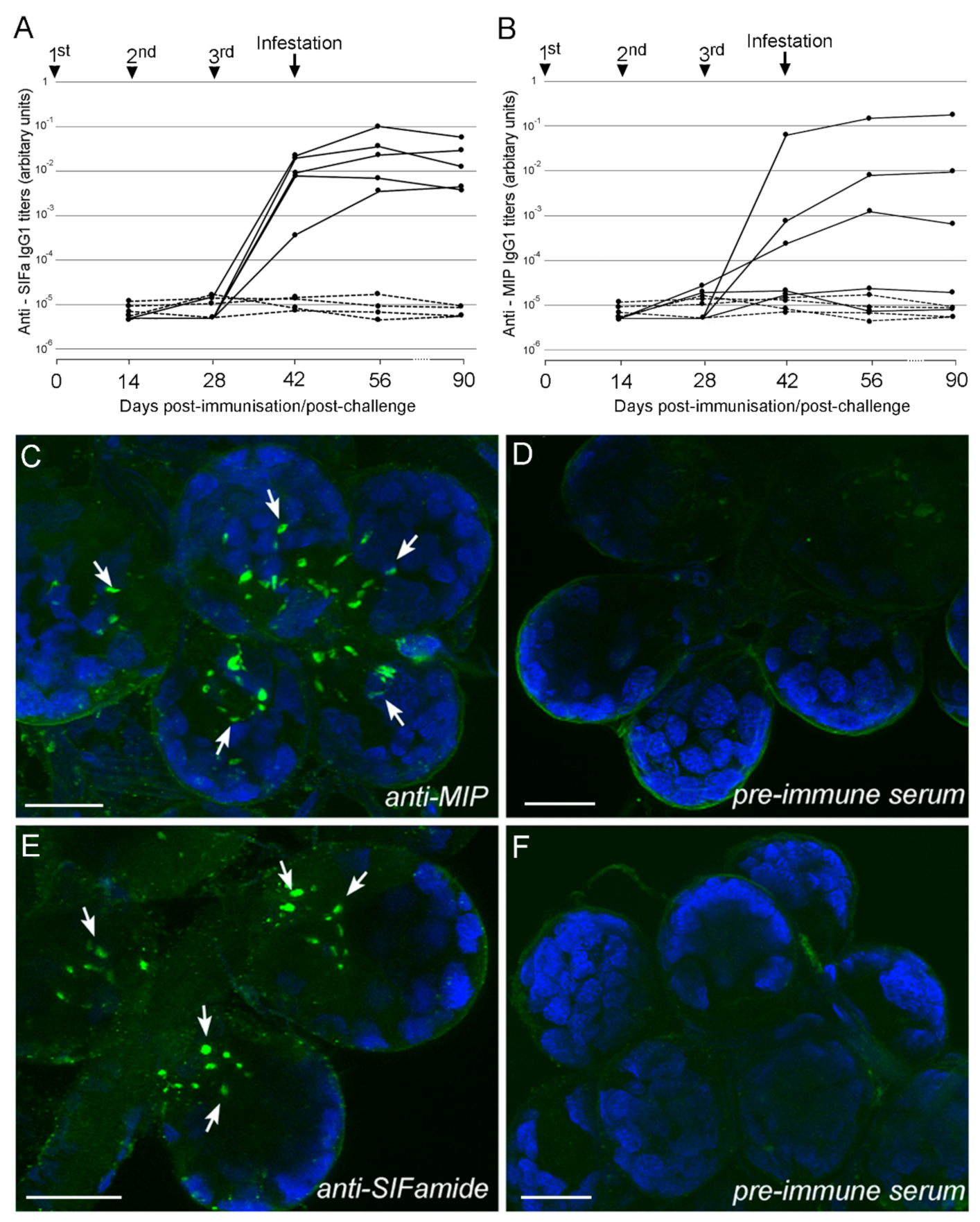
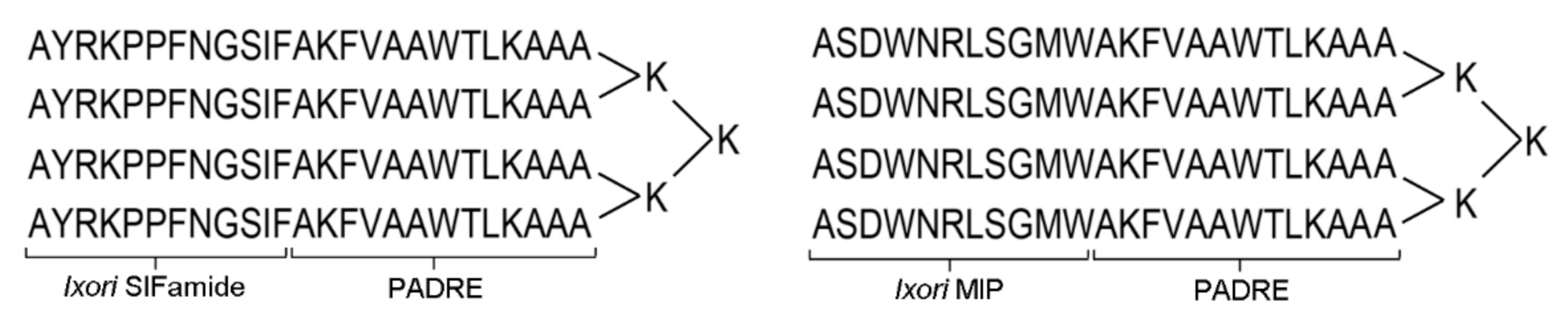
2.1. Immunogenicity of Multiple Antigenic Peptides (MAPs) in Mice and Sheep
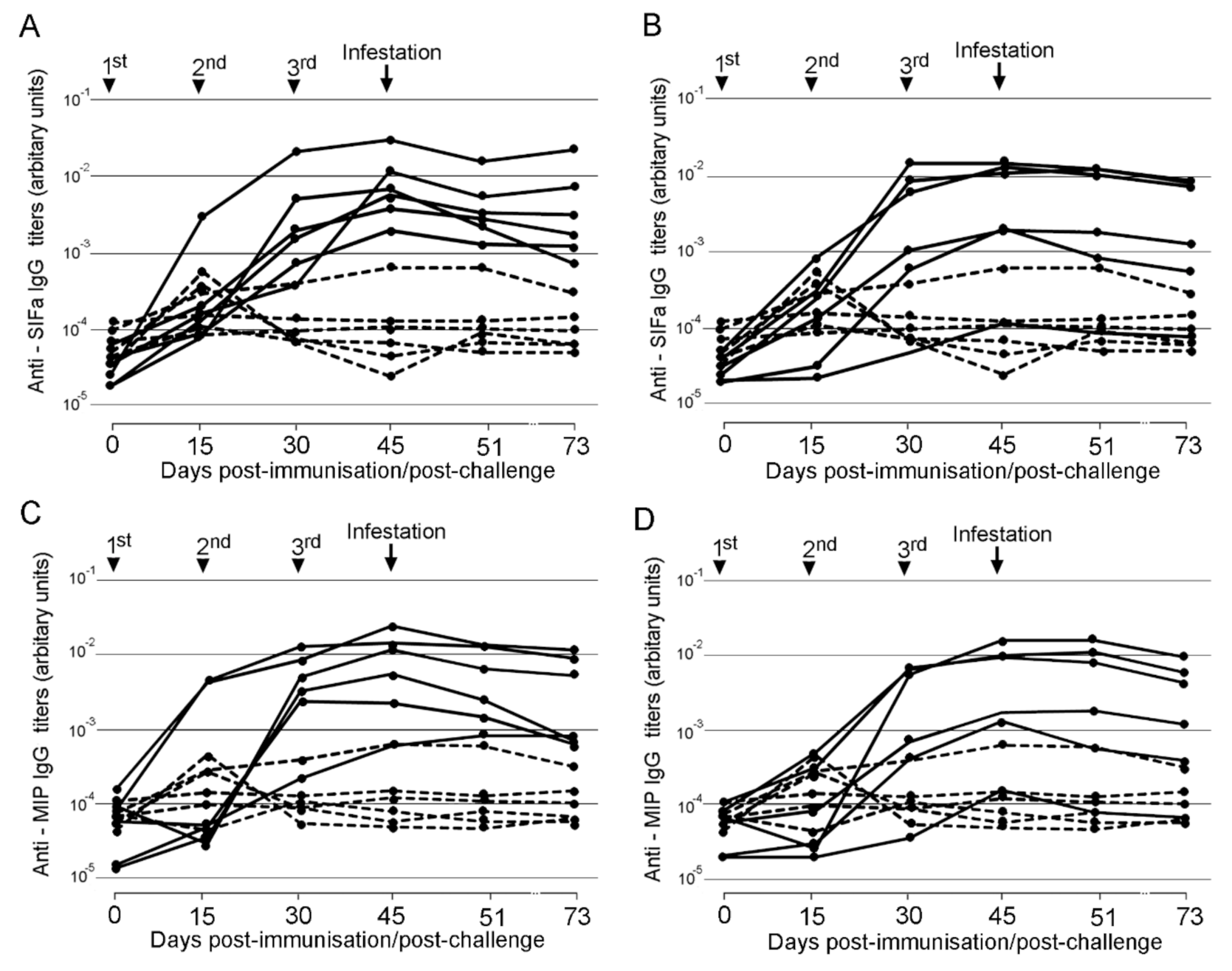
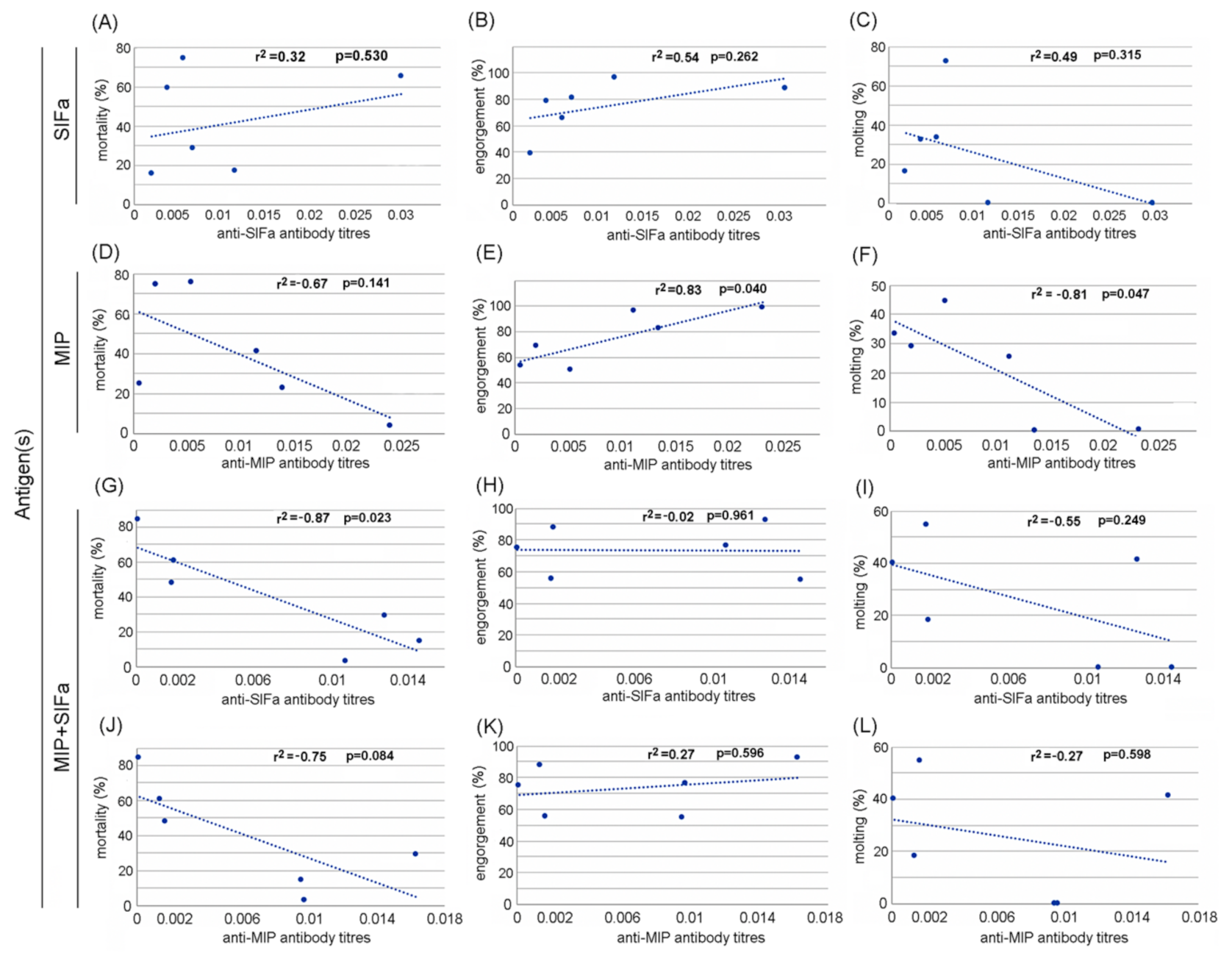
2.2. Efficacy of MAPs in Prevention of Tick Infestation and Development
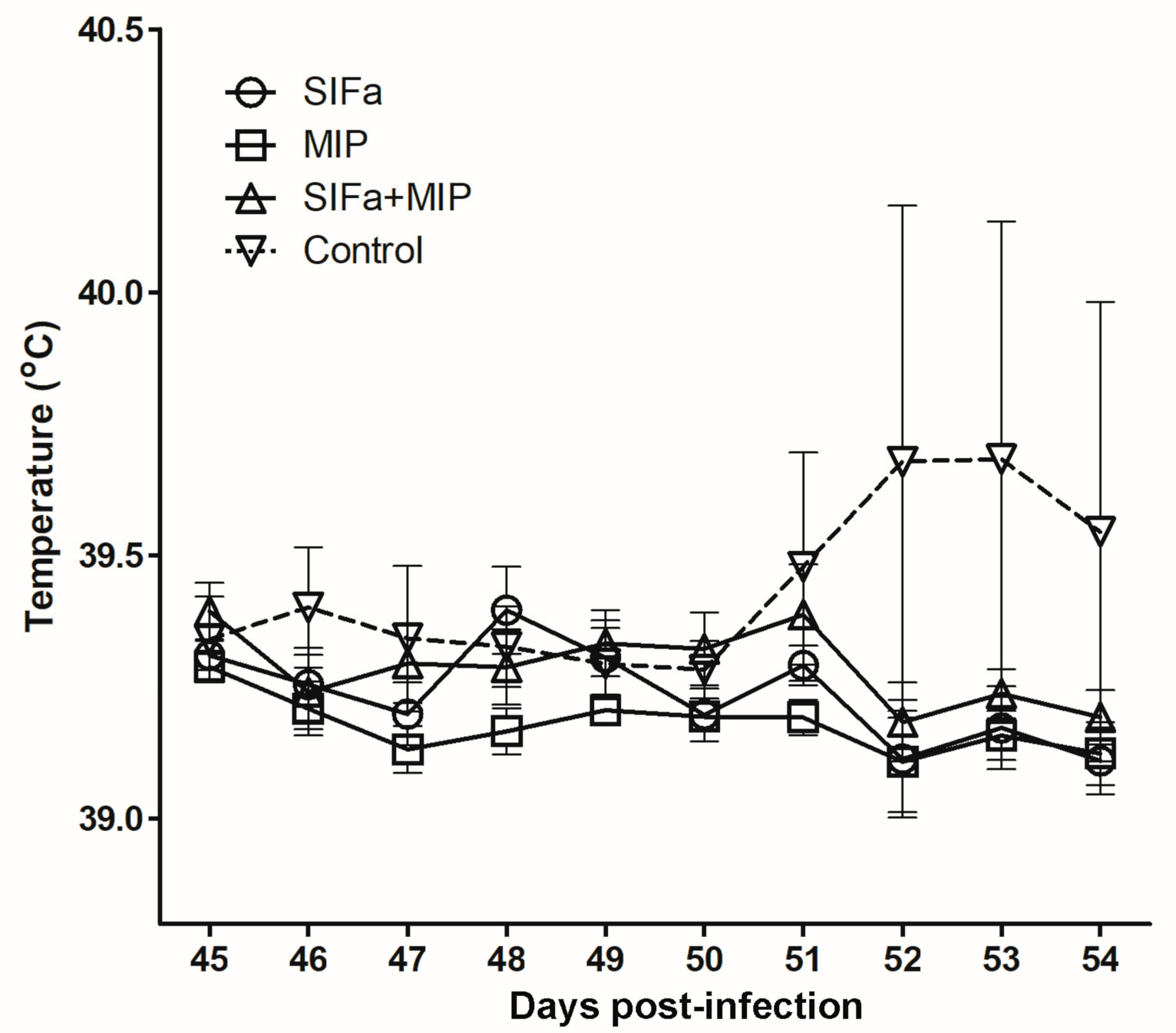
2.3. Impact of Vaccination on A. Phagocytophilum Transmission
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Ticks
4.3. Synthesis of Multiple Antigenic Peptides
4.4. Vaccine Formulation
4.5. Immunization of Mice and Tick Infestation
4.6. Whole-Mount Immunohistochemistry
4.7. Immunization of Sheep and Tick Infestation
4.8. Evaluation of Vaccine Innocuity
4.9. Follow-Up of A. phagocytophilum Infection in Sheep
4.10. Enzyme Linked Immunosorbent Assay (ELISA)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Fact Sheet: Vector-Borne Diseases. 2020. Available online: https://www.who.int/en/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 26 October 2020).
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Foldvari, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalska, E.; et al. Ixodes ricinus and Its Transmitted Pathogens in Urban and Peri-Urban Areas in Europe: New Hazards and Relevance for Public Health. Front. Pub. Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Trimnell, A.R.; Kazimírová, M.; Labuda, M. Exposed and concealed antigens as vaccine targets for controlling ticks and tick-borne diseases. Parasit. Immunol. 2006, 28, 155–163. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Merino, O. Vaccinomics, the new road to tick vaccines. Vaccine 2013, 31, 5923–5929. [Google Scholar] [CrossRef] [PubMed]
- Rego, R.O.M.; Trentelman, J.J.A.; Anguita, J.; Nijhof, A.M.; Sprong, H.; Klempa, B.; Hajdusek, O.; Tomás-Cortázar, J.; Azagi, T.; Strnad, M.; et al. Counterattacking the tick bite: Towards a rational design of anti-tick vaccines targeting pathogen transmission. Parasites Vectors 2019, 12, 229. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mallon, A. Developing Anti-tick Vaccines. Meth. Mol. Biol. 2016, 1404, 243–259. [Google Scholar] [CrossRef]
- Bhowmick, B.; Han, Q. Understanding Tick Biology and Its Implications in Anti-tick and Transmission Blocking Vaccines Against Tick-Borne Pathogens. Front. Vet. Sci. 2020, 7, 319. [Google Scholar] [CrossRef]
- Willadsen, P.; Bird, P.; Cobon, G.S.; Hungerford, J. Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 1995, 110 (Suppl. S1), S43–S50. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Rodríguez, M.; Montero, C.; Redondo, M.; García-García, J.C.; Méendez, L.; Serrano, E.; Valdés, M.; Enríquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine Gavac. Genet. Anal. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- Bonnet, S.; Kazimírová, M.; Richardson, J.; Šimo, L. Tick saliva and its role in pathogen transmission. In Skin and Arthropod Vectors; Boulanger, N., Ed.; Elsevier: Eastbourne, UK, 2018. [Google Scholar]
- Näassel, D.R.; Zandawala, M. Recent advances in neuropeptide signaling in Drosophila, from genes to physiology and behavior. Prog. Neurobiol. 2019, 179, 101607. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Žitňan, D.; Park, Y. Two novel neuropeptides in innervation of the salivary glands of the black-legged tick, Ixodes scapularis: Myoinhibitory peptide and SIFamide. J. Comp. Neurol. 2009, 517, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Park, Y. Neuropeptidergic control of the hindgut in the black-legged tick Ixodes scapularis. Int. J. Parasitol. 2014, 44, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimo, L.; Žitňan, D.; Park, Y. Neural control of salivary glands in ixodid ticks. J. Ins. Physiol. 2012, 58, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Šimo, L.; Vancová, M.; Urban, J.; Park, Y. Neural and endocrine regulation of osmoregulatory organs in tick: Recent discoveries and implications. Gen. Comp. Endocrinol. 2019, 278, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Trager, W. Acquired immunity to ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Davis, N.T.; Blackburn, M.B.; Golubeva, E.G.; Hildebrand, J.G. Localization of myoinhibitory peptide immunoreactivity in Manduca sexta and Bombyx mori, with indications that the peptide has a role in molting and ecdysis. J. Exp. Biol. 2003, 206, 1449–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, C.; Pech, U.; Kobbenbring, S.; Pauls, D.; Bahl, B.; Sommer, M.V.; Pooryasin, A.; Barth, J.; Arias, C.W.P.; Vassiliou, C.; et al. SIFamide Translates Hunger Signals into Appetitive and Feeding Behavior in Drosoph. Cell. Rep. 2017, 20, 464–478. [Google Scholar] [CrossRef] [Green Version]
- Terhzaz, S.; Rosay, P.; Goodwin, S.F.; Veenstra, J.A. The neuropeptide SIFamide modulates sexual behavior in Drosophila. Biochem. Biophys. Res. Commun. 2007, 352, 305–310. [Google Scholar] [CrossRef]
- Nichols, R.; McCormick, J.; Lim, I. Multiple antigenic peptides designed to structurally related Drosophila peptides. Peptides 1997, 18, 41–45. [Google Scholar] [CrossRef]
- Del Guercio, M.F.; Alexander, J.; Kubo, R.T.; Arrhenius, T.; Maewal, A.; Appella, E.; Hoffman, S.L.; Jones, T.; Valmori, D.; Sakaguchi, K.; et al. Potent immunogenic short linear peptide constructs composed of B cell epitopes and Pan DR T helper epitopes (PADRE) for antibody responses in vivo. Vaccine 1997, 15, 441–448. [Google Scholar] [CrossRef]
- Coumou, J.; Wagemakers, A.; Trentelman, J.J.; Nijhof, A.M.; Hovius, J.W. Vaccination against Bm86 Homologues in Rabbits Does Not Impair Ixodes ricinus Feeding or Oviposition. PLoS ONE 2014, 10, e0123495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevot, P.P.; Couvreur, B.; Denis, V.; Brossard, M.; Vanhamme, L.; Godfroid, E. Protective immunity against Ixodes ricinus induced by a salivary serpin. Vaccine 2007, 25, 3284–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogden, N.H.; Bigras-Poulin, M.; O’Callaghan, C.J.; Barker, I.K.; Kurtenbach, K.; Lindsay, L.R.; Charron, D.F. Vector seasonality, host infection dynamics and fitness of pathogens transmitted by the tick Ixodes scapularis. Parasitology 2007, 134, 209–227. [Google Scholar] [CrossRef]
- Contreras, M.; de la Fuente, J. Control of infestations by Ixodes ricinus tick larvae in rabbits vaccinated with aquaporin recombinant antigens. Vaccine 2017, 35, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Contreras, M.; de la Fuente, J. Control of Ixodes ricinus and Dermacentor reticulatus tick infestations in rabbits vaccinated with the Q38 Subolesin/Akirin chimera. Vaccine 2016, 34, 3010–3013. [Google Scholar] [CrossRef] [PubMed]
- Almazán, C.; Fourniol, L.; Rouxel, C.; Alberdi, P.; Gandoin, C.; Lagrée, A.-C.; Boulouis, H.-J.; de la Fuente, J.; Bonnet, S.I. Experimental Ixodes ricinus-Sheep Cycle of Anaplasma phagocytophilum NV2Os Propagated in Tick Cell Cultures. Front. Vet. Sci. 2020, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Commission, E. National Competent Authorities for the Implementation of Directive 2010/63/EU on the Protection of Animals Used for Scientific Purposes. 2010. Available online: https://ec.europa.eu/environment/chemicals/lab_animals/pdf/guidance/animal_welfare_bodies/en.pdf (accessed on 26 October 2020).
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’Hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef]
- Almazán, C.; Bonnet, S.; Cote, M.; Slovák, M.; Park, Y.; Šimo, L. A Versatile Model of Hard Tick Infestation on Laboratory Rabbits. J. Vis. Exp. 2018, 140, e57994. [Google Scholar] [CrossRef]
- Alexander, J.; del Guercio, M.F.; Maewal, A.; Qiao, L.; Fikes, J.; Chesnut, R.W.; Paulson, J.; Bundle, D.R.; DeFrees, S.; Sette, A. Linear PADRE T helper epitope and carbohydrate B cell epitope conjugates induce specific high titer IgG antibody responses. J. Immunol. 2000, 164, 1625–1633. [Google Scholar] [CrossRef]
- Reis, C.; Cote, M.; Le Rhun, D.; Lecuelle, B.; Levin, M.L.; Vayssier-Taussat, M.; Bonnet, S.I. Vector competence of the tick Ixodes ricinus for transmission of Bartonella birtlesii. PLoS Negl. Trop. Dis. 2011, 5, e1186. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Koči, J.; Park, Y. Receptors for the neuropeptides, myoinhibitory peptide and SIFamide, in control of the salivary glands of the blacklegged tick Ixodes scapularis. Ins. Biochem. Mol. Biol. 2013, 43, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hope, M.; Jiang, X.; Gough, J.; Cadogan, L.; Josh, P.; Jonsson, N.; Willadsen, P. Experimental vaccination of sheep and cattle against tick infestation using recombinant 5′-nucleotidase. Paras. Immunol. 2010, 32, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; El Harrak, M. Comparative innocuity and efficacy of live and inactivated sheeppox vaccines. BMC Vet. Res. 2016, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Kocan, K.M.; Busby, A.T.; Allison, R.W.; Breshears, M.A.; Coburn, L.; Galindo, R.C.; Ayllón, N.; Blouin, E.F.; de la Fuente, J. Sheep experimentally infected with a human isolate of Anaplasma phagocytophilum serve as a host for infection of Ixodes scapularis ticks. Ticks Tick-Borne Dis. 2012, 3, 147–153. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Parameters | Fed on Control Mice | Fed on SIF Vaccinated Mice | Fed on MIP Vaccinated Mice |
|---|---|---|---|
| Engorgement (No of engorged nymphs/No of nymphs used for infestation) | 18/20 | 20/20 | 18/20 |
| 3/20 | 20/20 | 9/20 | |
| 3/20 | 15/20 | 18/20 | |
| 18/20 | 16/20 | 15/20 | |
| 17/20 | 9/20 | ||
| Total | 42/80 | 88/100 * | 69/100 |
| Group average | 53% | 88% | 69% |
| Feeding (mean weight after feeding/engorged nymph) (mg) | 4.37 (±1.35) | 4.00 (±1.33) | 3.89 (±1.13) * |
| Molting (No adults at day 109/No engorged nymphs) | 8/18 | 17/20 | 8/18 |
| 3/3 | 14/20 | 7/9 | |
| 1/3 | 15/15 | 18/18 | |
| 16/18 | 16/16 | 8/15 | |
| 15/17 | 7/9 | ||
| Total | 28/42 | 77/88 e * | 48/69 |
| Group average | 67% | 89% | 71% |
| Tick mortality (No of dead nymphs at day 109/total No engorged nymphs) | 10/18 | 3/20 | 10/18 |
| 0/3 | 6/20 | 2/9 | |
| 2/3 | 0/15 | 0/18 | |
| 2/18 | 0/16 | 7/15 | |
| 2/17 | 2/9 | ||
| Total | 14/42 | 11/88 * | 21/69 |
| Group average | 33.5% | 11% | 29.4% |
| Tick Parameters | Fed on Control Sheep | Fed on SIFa Vaccinated Sheep | Fed on MIP Vaccinated Sheep | Fed on SIFa + MIP Vaccinated Sheep |
|---|---|---|---|---|
| Larvae | ||||
| Engorgement (No of engorged larvae/No of larvae used for infestation) | 1340/2067 | 1483/1836 | 871/1747 | 1377/1836 |
| 1499/1701 | 1309/2000 | 1454/2126 | 1561/1789 | |
| 49/139 | 45/117 | 526/548 | 144/156 | |
| 156/196 | 242/308 | 56/105 | 21/38 | |
| 205/376 | 277/316 | 1027/1040 | 123/226 | |
| 207/553 | 863/900 | 187/228 | 437/576 | |
| Total | 3456/5032 | 4219/5477 ** | 4121/5794 * | 3663/4621 ** |
| Group average | 60% | 74% | 75% | 73% |
| Molting (No nymphs at Day 90/No alive engorged larvae) | 31/144 | 769/1063 | 94/212 | 87/217 |
| 54/217 | 112/339 | 108/373 | 112/616 | |
| 14/37 | 6/38 | 79/312 | 42/102 | |
| 39/128 | 32/99 | 14/42 | 6/11 | |
| 0/166 | 0/97 | 3/991 | 0/105 | |
| 0/169 | 0/722 | 0/145 | 0/424 | |
| Total | 138/861 | 919/2358 ** | 298/2075 | 247/1475 |
| Group average | 19% | 26% | 22% | 26% |
| Tick mortality (No of dead ticks at Day 90/total No engorged larvae) | 1196/1340 | 420/1483 | 659/871 | 1160/1377 |
| 1282/1499 | 970/1309 | 1081/1454 | 945/1561 | |
| 12/49 | 7/45 | 214/526 | 42/144 | |
| 28/156 | 143/242 | 14/56 | 10/21 | |
| 39/205 | 180/277 | 36/1027 | 18/123 | |
| 38/207 | 141/863 | 42/187 | 13/437 | |
| Total | 2595/3456 | 1738/4219 ** | 2046/4121 ** | 2188/3663 ** |
| Group average | 42% | 43% | 40% | 40% |
| Nymphs | ||||
| Engorgement (No of engorged nymphs/No of nymphs used for infestation) | 10/48 | 0/48 | 1/48 | 0/48 |
| 1/48 | 1/48 | 3/48 | 6/48 | |
| 0/48 | 0/48 | 0/48 | 1/48 | |
| 0/48 | 0/48 | 1/48 | 1/48 | |
| 0/48 | 0/48 | 0/48 | 0/48 | |
| 0/48 | 0/48 | 2/48 | 0/48 | |
| Total | 11/288 | 1/288 ** | 7/288 | 8/288 |
| Group average | 3.8% | 0.3% | 2.4% | 2.8% |
| Molting (No adults at Day 90/No engorged nymphs) | 6/10 | 0/0 | 0/1 | 0/0 |
| 1/1 | 1/1 | 3/3 | 5/6 | |
| 0/0 | 0/0 | 0/0 | 0/1 | |
| 0/0 | 0/0 | 0/1 | 0/1 | |
| 0/0 | 0/0 | 0/0 | 0/0 | |
| 0/0 | 0/0 | 1/2 | 0/0 | |
| Total | 7/11 | 1/1 | 4/7 | 5/8 |
| Group average | 80% | 100% | 37.5% | 27.8% |
| Tick mortality (No of dead ticks at Day 90/total No attached nymphs) | 35/41 | 36/36 | 27/27 | 34/34 |
| 34/35 | 28/29 | 36/39 | 31/36 | |
| 25/25 | 31/31 | 27/27 | 47/48 | |
| 30/30 | 39/39 | 29/29 | 42/43 | |
| 39/39 | 41/41 | 48/48 | 40/40 | |
| 48/48 | 45/45 | 41/42 | 46/48 | |
| Total | 211/218 | 220/221* | 218/222 | 240/249 |
| Group average | 97% | 99% | 98% | 96% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almazán, C.; Šimo, L.; Fourniol, L.; Rakotobe, S.; Borneres, J.; Cote, M.; Peltier, S.; Mayé, J.; Versillé, N.; Richardson, J.; et al. Multiple Antigenic Peptide-Based Vaccines Targeting Ixodes ricinus Neuropeptides Induce a Specific Antibody Response but Do Not Impact Tick Infestation. Pathogens 2020, 9, 900. https://doi.org/10.3390/pathogens9110900
Almazán C, Šimo L, Fourniol L, Rakotobe S, Borneres J, Cote M, Peltier S, Mayé J, Versillé N, Richardson J, et al. Multiple Antigenic Peptide-Based Vaccines Targeting Ixodes ricinus Neuropeptides Induce a Specific Antibody Response but Do Not Impact Tick Infestation. Pathogens. 2020; 9(11):900. https://doi.org/10.3390/pathogens9110900
Chicago/Turabian StyleAlmazán, Consuelo, Ladislav Šimo, Lisa Fourniol, Sabine Rakotobe, Jérémie Borneres, Martine Cote, Sandy Peltier, Jennifer Mayé, Nicolas Versillé, Jennifer Richardson, and et al. 2020. "Multiple Antigenic Peptide-Based Vaccines Targeting Ixodes ricinus Neuropeptides Induce a Specific Antibody Response but Do Not Impact Tick Infestation" Pathogens 9, no. 11: 900. https://doi.org/10.3390/pathogens9110900