Molecular Diagnosis, Prevalence and Importance of Zoonotic Vector-Borne Pathogens in Cuban Shelter Dogs—A Preliminary Study

 ,
,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
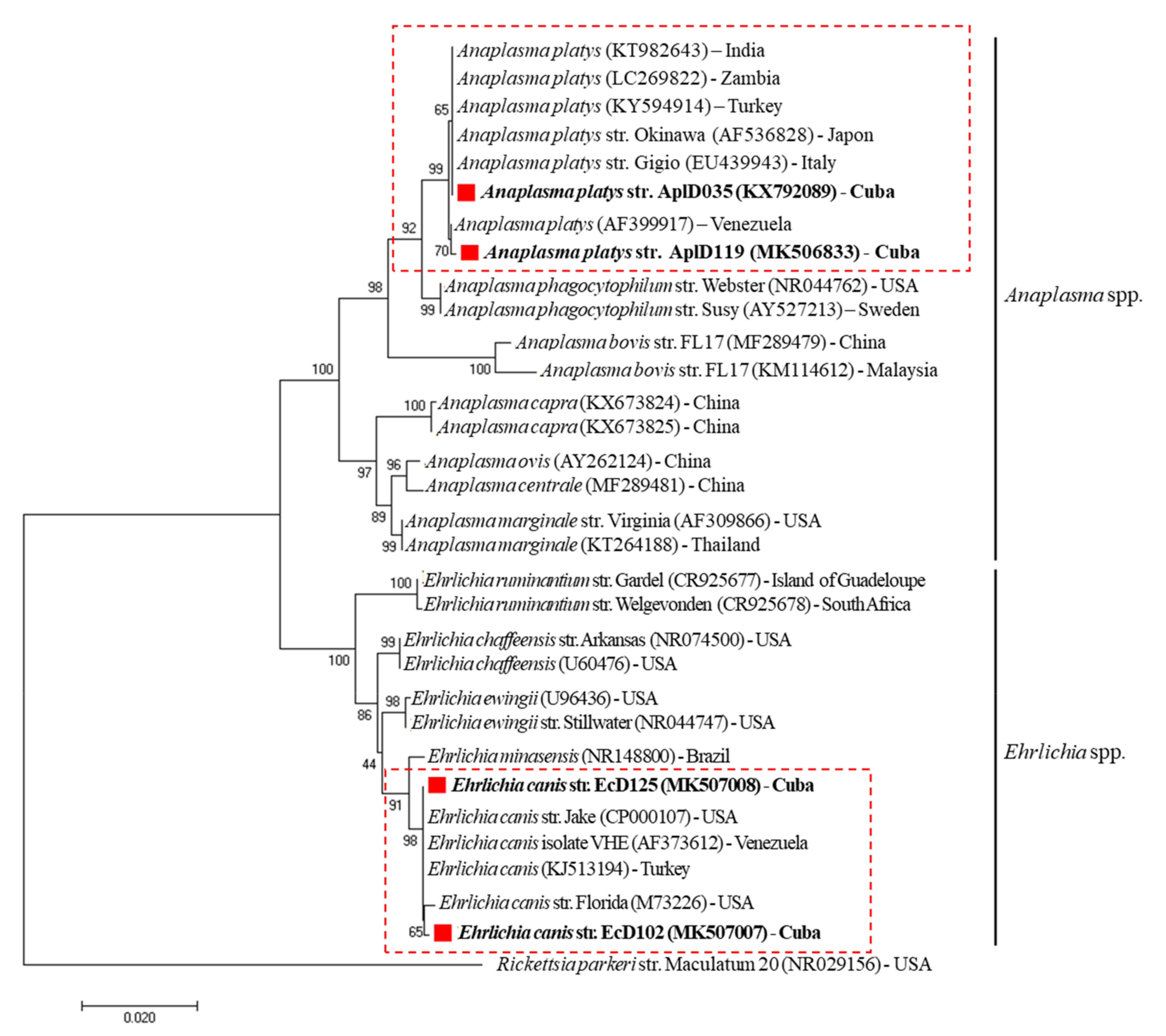
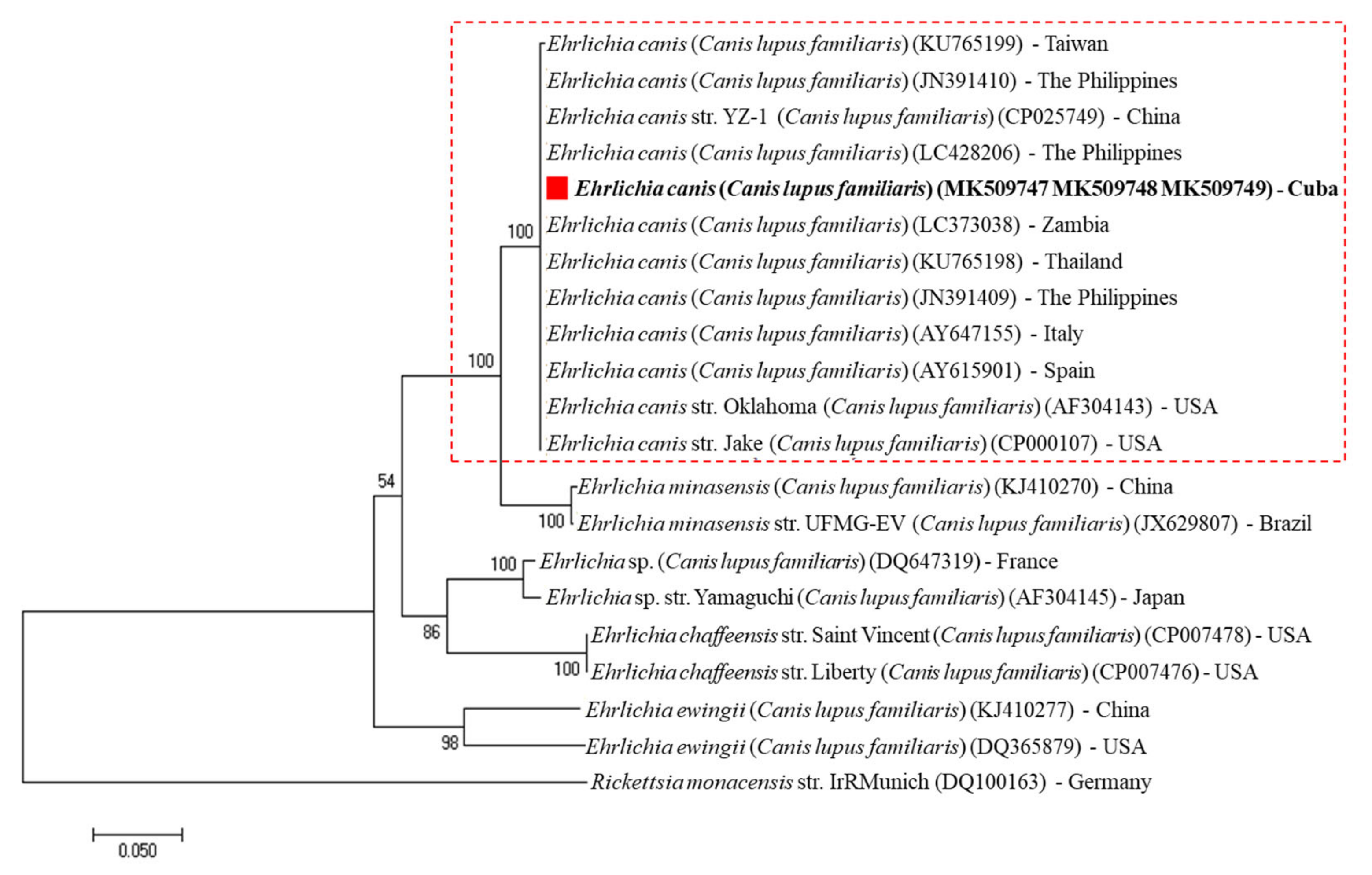
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection and DNA Extraction
4.2. Haematological Parameters
4.3. PCR Amplification and Sequencing
4.4. Sequence Analysis
4.5. Phylogenetic Analysis
4.6. Data Analysis
4.7. Ethical Approval
4.8. Nucleotide Sequence Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Otranto, D.; Dantas-Torres, F.; Mihalca, A.D.; Traub, R.J.; Lappin, M.; Baneth, G. Zoonotic Parasites of Sheltered and Stray Dogs in the Era of the Global Economic and Political Crisis. Trends Parasitol. 2017, 33, 813–825. [Google Scholar] [CrossRef]
- Chomel, B.B. Tick-borne infections in dogs—An emerging infectious threat. Veter. Parasitol. 2011, 179, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.J.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ’HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, M.; Bodor, M.; Zhang, C.; Xiong, Q.; Rikihisa, Y. Human Infection with Ehrlichia Canis Accompanied by Clinical Signs in Venezuela. Ann. N. Y. Acad. Sci. 2006, 1078, 110–117. [Google Scholar] [CrossRef]
- Bouza-Mora, L.; Dolz, G.; Solórzano-Morales, A.; Romero-Zuñiga, J.J.; Salazar-Sánchez, L.; Labruna, M.B.; Aguiar, D.M. Novel genotype of Ehrlichia canis detected in samples of human blood bank donors in Costa Rica. Ticks Tick Borne Dis. 2017, 8, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Little, S.E. Ehrlichiosis and Anaplasmosis in Dogs and Cats. Vet. Clin. Small Anim. Pract. 2010, 40, 1121–1140. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.; Soares, P.; Ramos, C.; Araújo, F.; Ramos, R.; Souza, I.; Faustino, M.; Alves, L. Molecular detection of Anaplasma platys in a naturally-infected cat in Brazil. Braz. J. Microbiol. 2010, 41, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Dahmani, M.; Davoust, B.; Benterki, M.S.; Fenollar, F.; Raoult, D.; Mediannikov, O. Development of a new PCR-based assay to detect Anaplasmataceae and the first report of Anaplasma phagocytophilum and Anaplasma platys in cattle from Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2015, 39, 39–45. [Google Scholar] [CrossRef]
- Cardoso, L.; Tuna, J.; Vieira, L.; Yisaschar-Mekuzas, Y.; Baneth, G. Molecular detection of Anaplasma platys and Ehrlichia canis in dogs from the North of Portugal. Veter. J. 2010, 183, 232–233. [Google Scholar] [CrossRef]
- Arraga-Alvarado, C.M.; Parra, O.C.; Hegarty, B.C.; Breitschwerdt, E.B.; Qurollo, B.A.; Berrueta, M.A. Molecular Evidence of Anaplasma platys Infection in Two Women from Venezuela. Am. J. Trop. Med. Hyg. 2014, 91, 1161–1165. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Veter. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef]
- Tsiodras, S.; Spanakis, N.; Spanakos, G.; Pervanidou, D.; Georgakopoulou, T.; Campos, E.; Petra, T.; Kanellopoulos, P.; Georgiadis, G.; Antalis, E.; et al. Fatal human anaplasmosis associated with macrophage activation syndrome in Greece and the Public Health response. J. Infect. Public Health 2017, 10, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on Tick-Borne Rickettsioses around the World: A Geographic Approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, J.C.Q.; Faccini-Martínez, Á.A.; González, J.D.R.; Díaz, F.J.; García, R.R.; Ordosgoitia, P.S.; Saad, E.A.P.; Quintero, L.O.; Arbeláez, C.R. Fatal Rickettsia rickettsii infection in a child, Northwestern Colombia, 2017. Ticks Tick Borne Dis. 2019, 10, 995–996. [Google Scholar] [CrossRef]
- Harrus, S.; Perlman-Avrahami, A.; Mumcuoglu, K.Y.; Morick, D.; Eyal, O.; Baneth, G. Molecular detection of Ehrlichia canis, Anaplasma bovis, Anaplasma platys, Candidatus Midichloria mitochondrii and Babesia canis vogeli in ticks from Israel. Clin. Microbiol. Infect. 2011, 17, 459–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamel, D.; Shukullari, E.; Rapti, D.; Silaghi, C.; Pfister, K.; Rehbein, S. Parasites and vector-borne pathogens in client-owned dogs in Albania. Blood pathogens and seroprevalences of parasitic and other infectious agents. Parasitol. Res. 2015, 115, 489–499. [Google Scholar] [CrossRef]
- Harrus, S.; Waner, T. Diagnosis of canine monocytotropic ehrlichiosis (Ehrlichia canis): An overview. Vet. J. 2011, 187, 292–296. [Google Scholar] [CrossRef]
- Ben Said, M.; Belkahia, H.; El Mabrouk, N.; Saidani, M.; Alberti, A.; Zobba, R.; Cherif, A.; Mahjoub, T.; Bouattour, A.; Messadi, L. Anaplasma platys-like strains in ruminants from Tunisia. Infect. Genet. Evol. 2017, 49, 226–233. [Google Scholar] [CrossRef]
- Navarrete, M.G.; Cordeiro, M.D.; Silva, C.B.; Massard, C.L.; López, E.R.; Rodríguez, J.C.A.; Ribeiro, C.C.; Fonseca-Rodríguez, O.; Da Fonseca, A.H. Serological and molecular diagnosis of Ehrlichia canis and associated risk factors in dogs domiciled in western Cuba. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 170–175. [Google Scholar] [CrossRef]
- Da Silva, C.B.; Santos, H.A.; Navarrete, M.G.; Ribeiro, C.C.D.U.; Corona, B.; Zaldivar, M.F.; Pires, M.S.; Peckle, M.; Da Costa, R.L.; Vitari, G.L.V.; et al. Molecular detection and characterization of Anaplasma platys in dogs and ticks in Cuba. Ticks Tick Borne Dis. 2016, 7, 938–944. [Google Scholar] [CrossRef]
- Maggi, R.G.; E Mascarelli, P.; Havenga, L.N.; Naidoo, V.; Breitschwerdt, E.B. Co-infection with Anaplasma platys, Bartonella henselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasites Vectors 2013, 6, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteo, J.A.; Portillo, A.; Santibáñez, S.; Blanco, J.; Pérez-Martínez, L.; Ibarra, V. Cluster of Cases of Human Rickettsia felis Infection from Southern Europe (Spain) Diagnosed by PCR. J. Clin. Microbiol. 2006, 44, 2669–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traversa, D.; Di Cesare, A.; Simonato, G.; Cassini, R.; Merola, C.; Diakou, A.; Halos, L.; Beugnet, F.; Di Regalbono, A.F. Zoonotic intestinal parasites and vector-borne pathogens in Italian shelter and kennel dogs. Comp. Immunol. Microbiol. Infect. Dis. 2017, 51, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ruttayaporn, N.; Saechan, V.; Jirapattharasate, C.; Vudriko, P.; Moumouni, P.F.A.; Cao, S.; Inpankaew, T.; Ybañez, A.P.; Suzuki, H.; et al. Molecular survey of canine vector-borne diseases in stray dogs in Thailand. Parasitol. Int. 2016, 65, 357–361. [Google Scholar] [CrossRef]
- Soares, R.; Ramos, C.A.; Pedroso, T.; Babo-Terra, V.; Cleveland, H.; De Araújo, F. Molecular survey of Anaplasma platys and Ehrlichia canis in dogs from Campo Grande, Mato Grosso do Sul, Brazil. Anais da Academia Brasileira de Ciências 2017, 89, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Starkey, L.A.; Newton, K.; Brunker, J.; Crowdis, K.; Edourad, E.J.P.; Meneus, P.; Little, S.E. Prevalence of vector-borne pathogens in dogs from Haiti. Vet. Parasitol. 2016, 224, 7–12. [Google Scholar] [CrossRef]
- Paulino, P.G.; Pires, M.S.; Da Silva, C.B.; Peckle, M.; Da Costa, R.L.; Vitari, G.V.; Vilela, J.A.R.; De Abreu, A.P.M.; Massard, C.L.; Santos, H.A. Epidemiology of Ehrlichia canis in healthy dogs from the Southeastern region of the state of Rio de Janeiro, Brazil. Prev. Veter. Med. 2018, 159, 135–142. [Google Scholar] [CrossRef]
- Inpankaew, T.; Hii, S.F.; Chimnoi, W.; Traub, R.J. Canine vector-borne pathogens in semi-domesticated dogs residing in northern Cambodia. Parasites Vectors 2016, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Dantas-Torres, F. The brown dog tick, Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae): From taxonomy to control. Veter. Parasitol. 2008, 152, 173–185. [Google Scholar] [CrossRef]
- Manzillo, V.F.; Cappiello, S.; Oliva, G. Tick-transmitted diseases in dogs: Clinicopathological findings. Parassitologia 2006, 48, 135–136. [Google Scholar]
- Bulla, C.; Takahira, R.K.; Trinca, L.A.; Lopes, R.S.; Wiedmeyer, C.E. The relationship between the degree of thrombocytopenia and infection with Ehrlichia canis in an endemic area. Veter. Res. 2004, 35, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.; Coppede, J.D.S.; Pereira, A.L.; Oliveira, L.P.; Roberto, P.G.; Benedetti, R.B.; Zucoloto, L.B.; Lucas, F.; Sobreira, L.; Marins, M. Molecular evaluation of the incidence of Ehrlichia canis, Anaplasma platys and Babesia spp. in dogs from Ribeirão Preto, Brazil. Veter. J. 2009, 179, 145–148. [Google Scholar] [CrossRef]
- De La Fuente, J.; Torina, A.; Naranjo, V.; Nicosia, S.; Alongi, A.; La Mantia, F.P.; Kocan, K.M. Molecular characterization of Anaplasma platys strains from dogs in Sicily, Italy. BMC Veter. Res. 2006, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Chisu, V.; Zobba, R.; Lecis, R.; Sotgiu, F.; Masala, G.; Foxi, C.; Pisu, D.; Alberti, A. GroEL typing and phylogeny of Anaplasma species in ticks from domestic and wild vertebrates. Ticks Tick Borne Dis. 2018, 9, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.; Yaaran, T.; Belshaw, A.; Curson, L.; Tisi, L.; Maurice, S.; Kiddle, G. A new TaqMan method for the reliable diagnosis of Ehrlichia spp. in canine whole blood. Parasites Vectors 2018, 11, 350. [Google Scholar] [CrossRef]
- Marsilio, F.; Di Martino, B.; Meridiani, I.; Bianciardi, P. Direct Identification ofEhrlichia Canisby a Novel Polymerase Chain Reaction Method and Molecular Analysis of the Citrate Synthase (gltA) Gene from Various Italian Strains. J. Veter. Diagn. Investig. 2006, 18, 215–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, A.; Zobba, R.; Chessa, B.; Addis, M.F.; Sparagano, O.A.E.; Parpaglia, M.L.P.; Cubeddu, T.; Pintori, G.; Pittau, M. Equine and Canine Anaplasma phagocytophilum Strains Isolated on the Island of Sardinia (Italy) Are Phylogenetically Related to Pathogenic Strains from the United States. Appl. Environ. Microbiol. 2005, 71, 6418–6422. [Google Scholar] [CrossRef] [Green Version]
- Barros-Battesti, D.M.; Hernandez, M.R.; Famadas, K.M.; Onofrio, V.C.; Beati, L.; Guglielmone, A.A. The ixodid ticks (Acari: Ixodidae) of Cuba. Syst. Appl. Acarol. 2009, 12, 101. [Google Scholar] [CrossRef]
- Rodríguez, I.; Fernández, C.; Sánchez, L.; Martinez, B.; Siegrist, H.H.; Lienhard, R. Serological evidences suggest Borrelia burgdorferi sensu lato infection in Cuba. Braz. J. Infect. Dis. 2012, 16, 405–406. [Google Scholar] [CrossRef] [Green Version]
- Stenos, J.; Unsworth, N.B.; Graves, S.R. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and Typhus group Rickettsiae. Am. J. Trop. Med. Hyg. 2005, 73, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Khrouf, F.; Sellami, H.; Elleuch, E.; Hattab, Z.; Ammari, L.; Khalfaoui, M.; Souissi, J.; Harrabi, H.; M’Ghirbi, Y.; Tiouiri, H.; et al. Molecular diagnosis of Rickettsia infection in patients from Tunisia. Ticks Tick Borne Dis. 2016, 7, 653–656. [Google Scholar] [CrossRef]
- Ng-Nguyen, D.; Hii, S.-F.; Hoang, M.-T.T.; Nguyen, V.-A.T.; Rees, R.; Stenos, J.; Traub, R.J. Domestic dogs are mammalian reservoirs for the emerging zoonosis flea-borne spotted fever, caused by Rickettsia felis. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Noda, A.A.; Rodriguez, I.; Miranda, J.; Mattar, S.; Cabezas-Cruz, A. First report of spotted fever group Rickettsia in Cuba. Ticks Tick Borne Dis. 2016, 7, 1057–1058. [Google Scholar] [CrossRef]
- Nicholson, W.L.; Allen, K.E.; McQuiston, J.H.; Breitschwerdt, E.B.; Little, S.E. The increasing recognition of rickettsial pathogens in dogs and people. Trends Parasitol. 2010, 26, 205–212. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.; Walker, A.R. Tick of Domestic Animals in Mediterranean Region: A Guide to Identification of Species; University of Zaragoza Press: Zaragoza, Spain, 2004; pp. 130–133. [Google Scholar]
- Sieber-Ruckstuhl, N.; Meli, M.; Boretti, F.S.; Gönczi, E.; Lutz, H.; Reusch, C. Quantitative Real-time PCR for the Measurement of 11?-HSD1 and 11?-HSD2 mRNA Levels in Tissues of Healthy Dogs. Horm. Metab. Res. 2007, 39, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Pusterla, N.; Huder, J.B.; Feige, K.; Lutz, H. Identification of a Granulocytic Ehrlichia Strain Isolated from a Horse in Switzerland and Comparison with Other Rickettsiae of the Ehrlichia phagocytophila Genogroup. J. Clin. Microbiol. 1998, 36, 2035–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann-Lehmann, R.; Wagmann, N.; Meli, M.L.; Riond, B.; Novacco, M.; Joekel, D.; Gentilini, F.; Marsilio, F.; Pennisi, M.G.; Lloret, A.; et al. Detection of ‘Candidatus Neoehrlichia mikurensis’ and other Anaplasmataceae and Rickettsiaceae in Canidae in Switzerland and Mediterranean countries. Schweizer Archiv für Tierheilkunde 2016, 158, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Leutenegger, C.M.; Pusterla, N.; Mislin, C.N.; Weber, R.; Lutz, H. Molecular Evidence of Coinfection of Ticks with Borrelia burgdorferi Sensu Lato and the Human Granulocytic Ehrlichiosis Agent in Switzerland. J. Clin. Microbiol. 1999, 37, 3390–3391. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.; Drazenovich, N.; Leutenegger, C.M.; Chomel, B.B. Association between polyarthritis and thrombocytopenia and increased prevalence of vectorborne pathogens in Californian dogs. Vet. Rec. 2007, 160, 159–162. [Google Scholar] [CrossRef] [Green Version]
- Barlough, J.E.; Madigan, J.E.; DeRock, E.; Bigornia, L. Nested polymerase chain reaction for detection of Ehrlichia equi genomic DNA in horses and ticks (Ixodes pacificus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [Green Version]
- Labruna, M.B.; McBride, J.W.; Bouyer, D.H.; Camargo, L.M.A.; Camargo, E.P.; Walker, D.H. Molecular Evidence for a Spotted Fever GroupRickettsiaSpecies in the TickAmblyomma longirostrein Brazil. J. Med. Entomol. 2004, 41, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector-Borne Pathogen(s) | Total | % | 95% IC a |
|---|---|---|---|
| Total infected dogs (≥1 pathogen) | 85 | 85.00 | 77.88–92.12 |
| Anaplasma phagocytophilum | 0 | 0.0 | |
| Anaplasma platys | 40 | 40.00 | 30.23–49.77 |
| Borrelia burgdorferi s.l. | 0 | 0.0 | |
| Ehrlichia canis | 62 | 62.00 | 52.32–71.68 |
| Rickettsia felisb | 27 | 27.00 | 18.15–35.85 |
| Single infections | 49 | 49.00 | 39.03–58.97 |
| Anaplasma phagocytophilum | 0 | 0.0 | |
| Anaplasma platys | 9 | 9.00 | 3.29–14.71 |
| Borrelia burgdorferi s.l. | 0 | 0.0 | |
| Ehrlichia canis | 29 | 29.00 | 19.95–38.05 |
| Rickettsia felisb | 11 | 11.00 | 4.76–17.24 |
| Co-infections | 36 | 36.00 | 26.43–45.57 |
| Anaplasma platys/Ehrlichia canis | 28 | 28.00 | 19.05–36.95 |
| Anaplasma platys/Rickettsia felisb | 11 | 11.00 | 4.76–17.24 |
| Ehrlichia canis/Rickettsia felisb | 13 | 13.00 | 6.29–19.71 |
| Anaplasma platys/Ehrlichia canis/Rickettsia felisb | 8 | 8.00 | 2.59–13.41 |
| Non-detected | 15 | 15.00 | 7.88–22.12 |
| Haematological Parameters | qPCR Positive | CBC Values | U Test | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| Dogs (%) | Range | Mean | Median | SD | SE | |||
| Haematocrit (L/L) | ||||||||
| Non-infected | 12 (13.33%) | 0.12–0.52 | 0.34 | 0.37 | 0.10 | 0.03 | ||
| Anaplasma platys | 8 (8.88%) | 0.15–0.47 | 0.35 | 0.39 | 0.11 | 0.04 | 38.50 | 0.485 |
| Ehrlichia canis | 24 (26.67%) | 0.13–0.53 | 0.34 | 0.36 | 0.11 | 0.02 | 140.5 | 0.920 |
| Rickettsia spp. | 11 (12.22%) | 0.30–0.57 | 0.41 | 0.42 | 0.09 | 0.03 | 42 | 0.146 |
| Co-infected | 35 (38.89%) | 0.16–0.60 | 0.37 | 0.36 | 0.12 | 0.02 | 191 | 0.651 |
| Haemoglobin (g/L) | ||||||||
| Non-infected | 12 (13.33%) | 50–189 | 124.08 | 131.00 | 33.90 | 9.77 | ||
| Anaplasma platys | 8 (8.88%) | 59–172 | 131.30 | 137.50 | 37.70 | 13.79 | 40 | 0.563 |
| Ehrlichia canis | 24 (26.67%) | 48–192 | 124.00 | 126.00 | 40.20 | 8.02 | 135 | 0.775 |
| Rickettsia spp. | 11 (12.22%) | 111–202 | 148.50 | 150.00 | 28.60 | 8.63 | 39 | 0.103 |
| Co-infected | 35 (38.89%) | 63–211 | 130.30 | 132.00 | 39.50 | 6.68 | 191 | 0.643 |
| RBC Count (×1012/L) | ||||||||
| Non-infected | 12 (13.33%) | 1.80–7.47 | 5.30 | 5.81 | 1.50 | 0.42 | ||
| Anaplasma platys | 8 (8.88%) | 2.28–7.46 | 5.60 | 5.97 | 1.80 | 0.68 | 42 | 0.678 |
| Ehrlichia canis | 24 (26.67%) | 1.97–7.80 | 5.40 | 5.39 | 1.80 | 0.36 | 140 | 0.906 |
| Rickettsia spp. | 11 (12.22%) | 4.25–8.49 | 6.20 | 6.32 | 1.20 | 0.36 | 43 | 0.169 |
| Co-infected | 35 (38.89%) | 2.53–9.90 | 5.80 | 5.70 | 1.70 | 0.28 | 190 | 0.493 |
| MCV (fL) | ||||||||
| Non-infected | 12 (13.33%) | 51–71 | 65.30 | 65.00 | 5.50 | 1.58 | ||
| Anaplasma platys | 8 (8.88%) | 63–69 | 66.00 | 66.50 | 2.40 | 0.80 | 42.5 | 0.698 |
| Ehrlichia canis | 24 (26.67%) | 53–89 | 64.80 | 63.50 | 6.90 | 1.45 | 113 | 0.304 |
| Rickettsia spp. | 11 (12.22%) | 61–72 | 66.50 | 67.00 | 3.60 | 1.07 | 62 | 0.828 |
| Co-infected | 35 (38.89%) | 55–74 | 64.40 | 65.00 | 4.80 | 0.81 | 182 | 0.493 |
| MCH (pg) | ||||||||
| Non-infected | 12 (13.33%) | 17.1–27.6 | 23.80 | 23.50 | 2.60 | 0.76 | ||
| Anaplasma platys | 8 (8.88%) | 21.7–26.2 | 23.90 | 23.60 | 1.70 | 0.62 | 41 | 0.616 |
| Ehrlichia canis | 24 (26.67%) | 17.3–31.7 | 23.40 | 23.10 | 3.20 | 0.67 | 123 | 0.491 |
| Rickettsia spp. | 11 (12.22%) | 21.9–26.8 | 24.00 | 23.80 | 1.60 | 0.48 | 64.5 | 0.951 |
| Co-infected | 35 (38.89%) | 17.8–27.4 | 22.60 | 22.40 | 2.20 | 0.37 | 126 | 0.042 * |
| MCHC (g/L) | ||||||||
| Non-infected | 12 (13.33%) | 332–400 | 363.30 | 361.00 | 15.90 | 4.60 | ||
| Anaplasma platys | 8 (8.88%) | 328–387 | 361.90 | 359.00 | 17.70 | 6.61 | 42 | 0.671 |
| Ehrlichia canis | 24 (26.67%) | 320–380 | 354.70 | 357.50 | 15.20 | 3.23 | 109 | 0.247 |
| Rickettsia spp. | 11 (12.22%) | 346–373 | 361.20 | 361.00 | 9.70 | 2.91 | 64 | 0.926 |
| Co-infected | 35 (38.89%) | 324–382 | 350.10 | 347.00 | 16.30 | 2.76 | 119 | 0.027 * |
| Platelets (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 60–610 | 307.90 | 303.50 | 205.30 | 59.26 | ||
| Anaplasma platys | 8 (8.88%) | 43–354 | 185.10 | 153.00 | 103.30 | 37.42 | 37 | 0.418 |
| Ehrlichia canis | 24 (26.67%) | 35–461 | 138.50 | 102.50 | 104.00 | 20.95 | 73 | 0.018 * |
| Rickettsia spp. | 11 (12.22%) | 132–698 | 300.00 | 264.00 | 167.20 | 50.42 | 64 | 0.926 |
| Co-infected | 35 (38.89%) | 44–480 | 134.40 | 104.00 | 89.90 | 15.20 | 111 | 0.016 * |
| WBC Count (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 4.60–13.70 | 10.30 | 10.60 | 2.60 | 0.75 | ||
| Anaplasma platys | 8 (8.88%) | 4.56–18 | 11.60 | 12.50 | 4.40 | 1.35 | 32 | 0.232 |
| Ehrlichia canis | 24 (26.67%) | 3.60–21.10 | 10.20 | 9.40 | 5.10 | 1.06 | 130.5 | 0.663 |
| Rickettsia spp. | 11 (12.22%) | 6–19.50 | 11.20 | 11.00 | 4.30 | 1.30 | 63.5 | 0.902 |
| Co-infected | 35 (38.89%) | 5.20–23.90 | 12.00 | 11.60 | 5.10 | 0.87 | 175 | 0.393 |
| Total Neutrophils (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 3.60–11.20 | 6.88 | 6.80 | 2.13 | 0.62 | ||
| Anaplasma platys | 8 (8.88%) | 4.30–13.50 | 7.58 | 6.35 | 3.70 | 1.33 | 45 | 0.847 |
| Ehrlichia canis | 24 (26.67%) | 1.32–14.60 | 6.64 | 5.85 | 3.63 | 0.76 | 129 | 0.626 |
| Rickettsia spp. | 11 (12.22%) | 3.90–14.50 | 7.65 | 6.10 | 3.72 | 1.12 | 64.5 | 0.951 |
| Co-infected | 35 (38.89%) | 2.20–70 | 11.73 | 7.40 | 14.26 | 0.73 | 181 | 0.487 |
| Lymphocytes (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 0.70–4.20 | 2.74 | 2.74 | 1.06 | 0.31 | ||
| Anaplasma platys | 8 (8.88%) | 0.80–8.90 | 3.39 | 3.15 | 2.30 | 0.79 | 35 | 0.334 |
| Ehrlichia canis | 24 (26.67%) | 0.50–10.80 | 2.42 | 2.25 | 2.05 | 0.43 | 102 | 0.163 |
| Rickettsia spp. | 11 (12.22%) | 1.40–6 | 2.75 | 2.40 | 1.29 | 0.39 | 61.5 | 0.805 |
| Co-infected | 35 (38.89%) | 0.80–31 | 4.11 | 2.50 | 5.91 | 0.27 | 186 | 0.566 |
| Monocytes (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 0.30–1.10 | 0.68 | 0.70 | 0.22 | 0.06 | ||
| Anaplasma platys | 8 (8.88%) | 0.30–1.10 | 0.73 | 0.80 | 0.24 | 0.07 | 33.5 | 0.272 |
| Ehrlichia canis | 24 (26.67%) | 0.10–1.10 | 0.61 | 0.60 | 0.27 | 0.06 | 130 | 0.648 |
| Rickettsia spp. | 11 (12.22%) | 0.20–1.50 | 0.76 | 0.70 | 0.43 | 0.13 | 65 | 0.975 |
| Co-infected | 35 (38.89%) | 0.20–6 | 1.12 | 0.70 | 1.22 | 0.21 | 175 | 0.396 |
| Eosinophils (×109/L) | ||||||||
| Non-infected | 12 (13.33%) | 0.09–1.19 | 0.39 | 0.24 | 0.38 | 0.11 | ||
| Anaplasma platys | 8 (8.88%) | 0.06–1.01 | 0.47 | 0.49 | 0.37 | 0.13 | 41 | 0.616 |
| Ehrlichia canis | 24 (26.67%) | 0.07–1.05 | 0.43 | 0.35 | 0.32 | 0.07 | 129 | 0.626 |
| Rickettsia spp. | 11 (12.22%) | 0.09–1.57 | 0.53 | 0.32 | 0.51 | 0.15 | 55 | 0.518 |
| Co-infected | 35 (38.89%) | 0.08–4 | 0.64 | 0.52 | 0.71 | 0.12 | 149 | 0.140 |
| Pathogens | Primers/Probes Sequences [5′—3′] | Target Gene | Amplicon Size | Reference |
|---|---|---|---|---|
| Internal control PCR | ||||
| cGAPDH.427p | 6-FAM—CCCTCAAGATTGTCAGCAATGCCTCCT—TAMRA | cGADPH | 131 bp | Sieber-Ruckstuhl et al. [48] |
| cGAPDH.395f | GATGGGCGTGAACCATGAG | |||
| cGAPDH.525r | TCATGAGGCCCTCCACGAT | |||
| Anaplasma phagocytophilum | ||||
| Ep.80p | 6-FAM—CCTATGCATTACTCACCCGTCTGCCACT—TAMRA | 16S rRNA | 106 bp | Pusterla et al. [49] |
| Ep.145f | CCATTTCTAGTGGCTATCCCATACTAC | |||
| Ep.50r | TCGAACGGATTATTCTTTATAGCTTG | |||
| Anaplasma platys | ||||
| Aplat_34p | 6-FAM—AGCTACGACAAAAATCCGTTCGACTTGCA—TAMRA | 16S rRNA | 75 bp | Hofmann-Lehmann et al. [50] |
| Aplat.14f | CTGGCGGCAAGCTTAACAC | |||
| Aplat.89r | CGTCTGCCACTATTTATCATAGC | |||
| Borrelia burgdorferi s.l. | ||||
| B.421p | 6-FAM—ATGTGCATTTGGTTATATTGAGCTTGATCAGCAA—TAMRA | flaB | 88 pb | Leutenegger et al. [51] |
| B.398f | GGGAAGCAGATTTGTTTGACA | |||
| B.484r | ATAGAGCAACTTACAGACGAAATTAATAGA | |||
| Ehrlichia canis | ||||
| Ec.61p | 6-FAM—TCTGCCACTAACAATTTCCTATAGCCAGAGGC—TAMRA | 16S rRNA | 108 pb | Foley et al. [52] |
| Ec.139f | ATGGCTATTCCGTACTACTAGGTAGATTC | |||
| Ec.32r | CATGCAAGTCGAACGGACAAT | |||
| Rickettsia spp. | ||||
| CS-P | 6-FAM—TGCAATAGCAAGAACCGTAGGCTGGATG—BHQ-1 | gltA | 74 pb | Stenos et al. [42] |
| CS-F | TCGCAAATGTTCACGGTACTTT | |||
| CS-R | TCGTGCATTTCTTTCCATTGTG | |||
| Pathogens | Primers Sequences (5′—3′) | Target Gene | Amplicon Size | Cycling Conditions * | References |
|---|---|---|---|---|---|
| Anaplasma spp./Ehrlichia spp. | 40 cycles: 10 s 98 °C; 1.5 min 72 °C | ||||
| EE1 | TCCTGGCTCAGAACGAACGCTGGCGGC | 16SrRNA | 1400 pb | Barlough et al. [53] | |
| EE2 | AGTCACTGACCCAACCTTAAATGGCTG | ||||
| Anaplasma platys | 35 cycles: 10 s 98 °C; 30 s 58 °C; 1 min 72 °C | ||||
| EphplgroEL.F | ATGGTATGCAGTTTGATCGC | groEL | 625 bp | Alberti et al. [39] | |
| EphplgroEL.R | TCTACTCTGTCTTTGCGTTC | ||||
| Ehrlichia canis | 35 cycles: 10 s 98 °C; 30 s 54 °C; 1 min 72 °C | ||||
| Ec.gltA.522f | CAGGAGTATATGCCTCCTGA | gltA | 507 pb | Marsilio et al. [38] | |
| Ec.gltA.1031r | GTTACTTTTTTCAATTGCC | ||||
| Rickettsia spp. | 40 cycles: 10 s 98 °C; 30 s 55 °C; 1 min 72 °C | ||||
| Rr190.70p | ATGGCGAATATTTCTCCAAAA | ompA | 532 pb | Regnery et al. [55] | |
| Rr190.620n | AGTGCAGCATTCGCTCCCCCT | ||||
| 120-M59 | CCGCAGGGTTGGTAACTGC | ompB | 862 bp | 10 s 98 °C; 30 s 55 °C; 1 min 72 °C | Roux and Raoult [54] |
| 120-807 | CCTTTTAGATTACCGCCTAA | ||||
| 17kD1 | GCTCTTGCAACTTCTATGTT | htrA | 434 bp | 10 s 98 °C; 30 s 55 °C; 1 min 72 °C | Labruna et al. [56] |
| 17kD2 | CATTGTTCGTCAGGTTGGCG | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Sánchez, A.A.; Corona-González, B.; Meli, M.L.; Roblejo-Arias, L.; Fonseca-Rodríguez, O.; Pérez Castillo, A.; Vega Cañizares, E.; Lobo Rivero, E.; Hofmann-Lehmann, R. Molecular Diagnosis, Prevalence and Importance of Zoonotic Vector-Borne Pathogens in Cuban Shelter Dogs—A Preliminary Study. Pathogens 2020, 9, 901. https://doi.org/10.3390/pathogens9110901
Díaz-Sánchez AA, Corona-González B, Meli ML, Roblejo-Arias L, Fonseca-Rodríguez O, Pérez Castillo A, Vega Cañizares E, Lobo Rivero E, Hofmann-Lehmann R. Molecular Diagnosis, Prevalence and Importance of Zoonotic Vector-Borne Pathogens in Cuban Shelter Dogs—A Preliminary Study. Pathogens. 2020; 9(11):901. https://doi.org/10.3390/pathogens9110901
Chicago/Turabian StyleDíaz-Sánchez, Adrian Alberto, Belkis Corona-González, Marina L. Meli, Lisset Roblejo-Arias, Osvaldo Fonseca-Rodríguez, Anisleidy Pérez Castillo, Ernesto Vega Cañizares, Evelyn Lobo Rivero, and Regina Hofmann-Lehmann. 2020. "Molecular Diagnosis, Prevalence and Importance of Zoonotic Vector-Borne Pathogens in Cuban Shelter Dogs—A Preliminary Study" Pathogens 9, no. 11: 901. https://doi.org/10.3390/pathogens9110901