
Evaluation of Wild Peanut Species and Their Allotetraploids for Resistance against Thrips and Thrips-Transmitted Tomato Spotted Wilt Orthotospovirus (TSWV)


 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Thrips Colonies
2.2. Peanut Genotypes
2.3. Thrips-Mediated TSWV Transmission
2.3.1. Incidence of TSWV Infection
2.3.2. Quantitation of Virus Loads in Infected Samples
2.3.3. Thrips Feeding Damage Index
2.3.4. Thrips Fitness
2.3.5. Correlation between Final TSWV Infection and Thrips Feeding Injury
2.4. Statistical Analyses
3. Results
3.1. TSWV Symptoms on Diploids and Allotetraploids
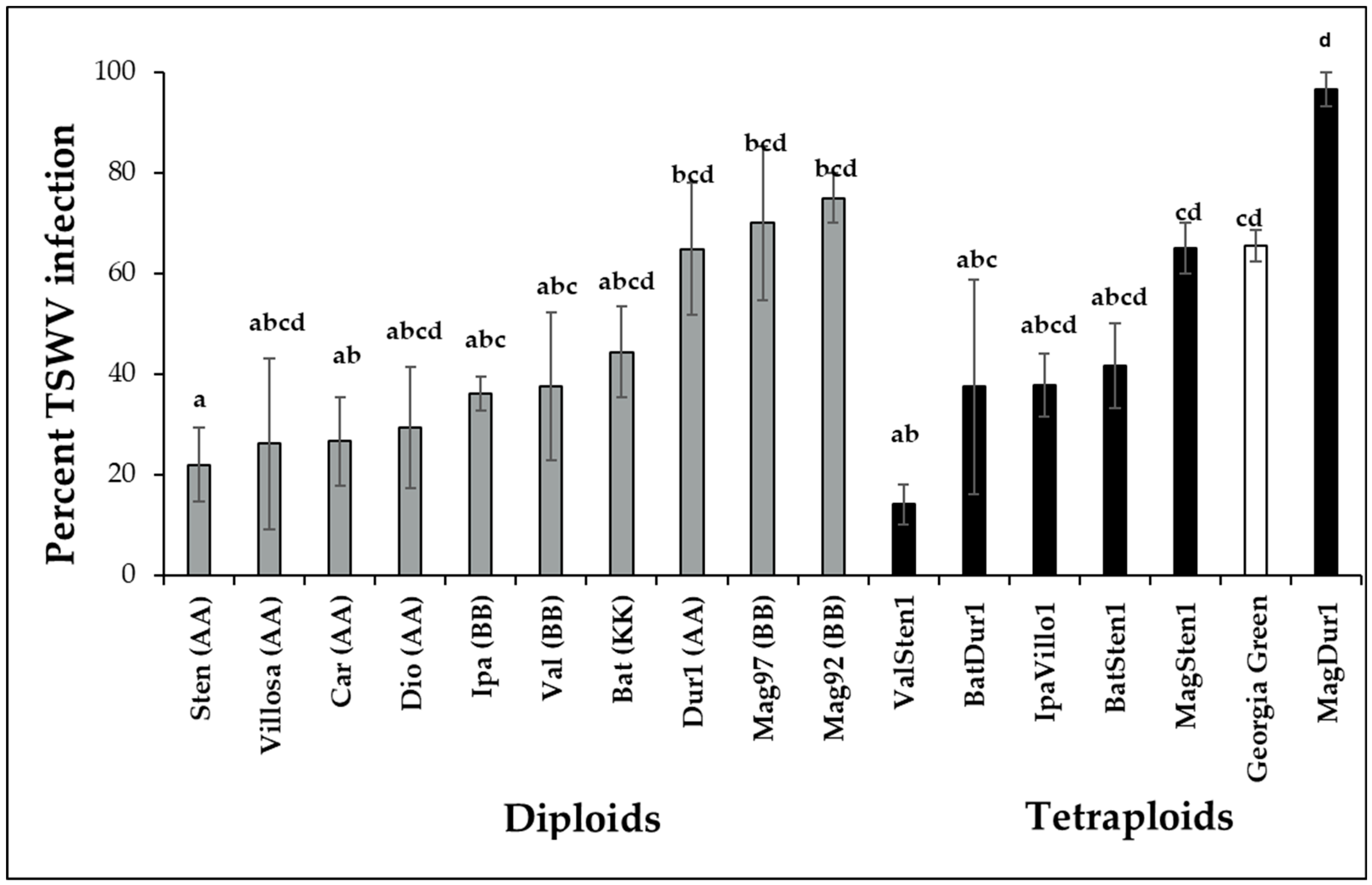
3.2. Incidence of TSWV Infection
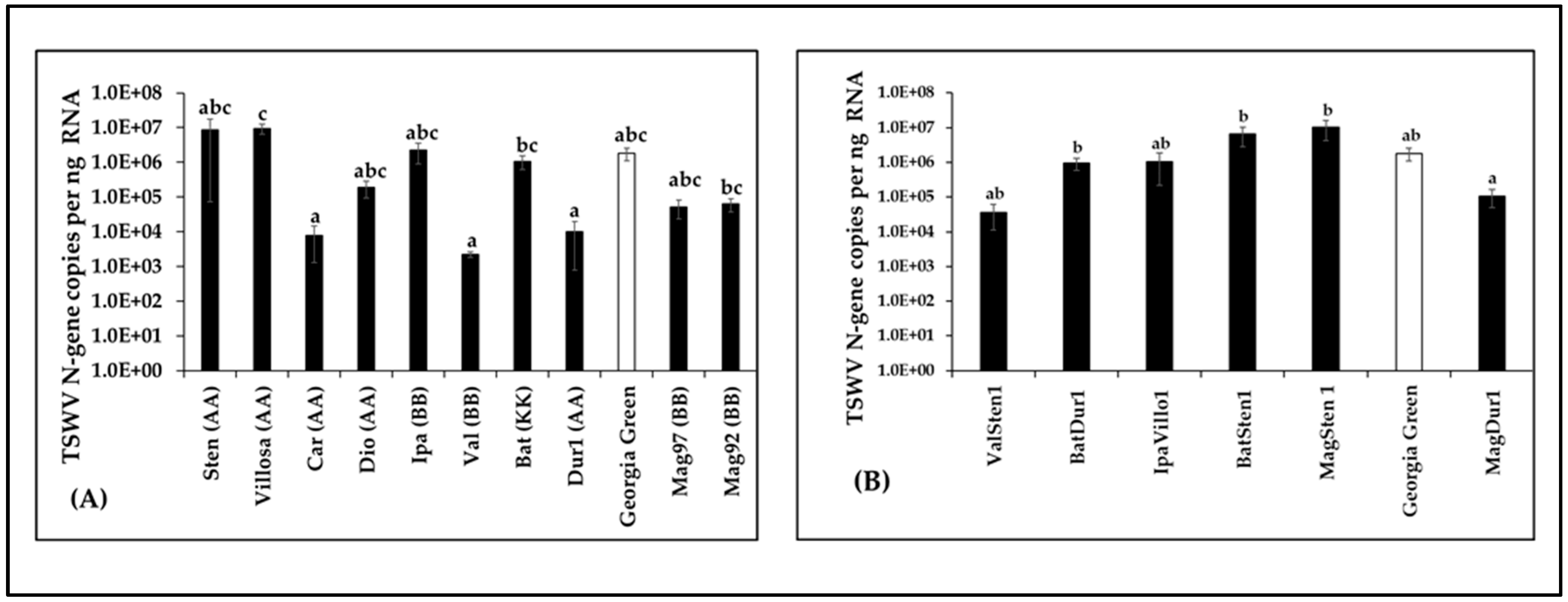
3.3. Quantitation of Virus Loads in Infected Samples
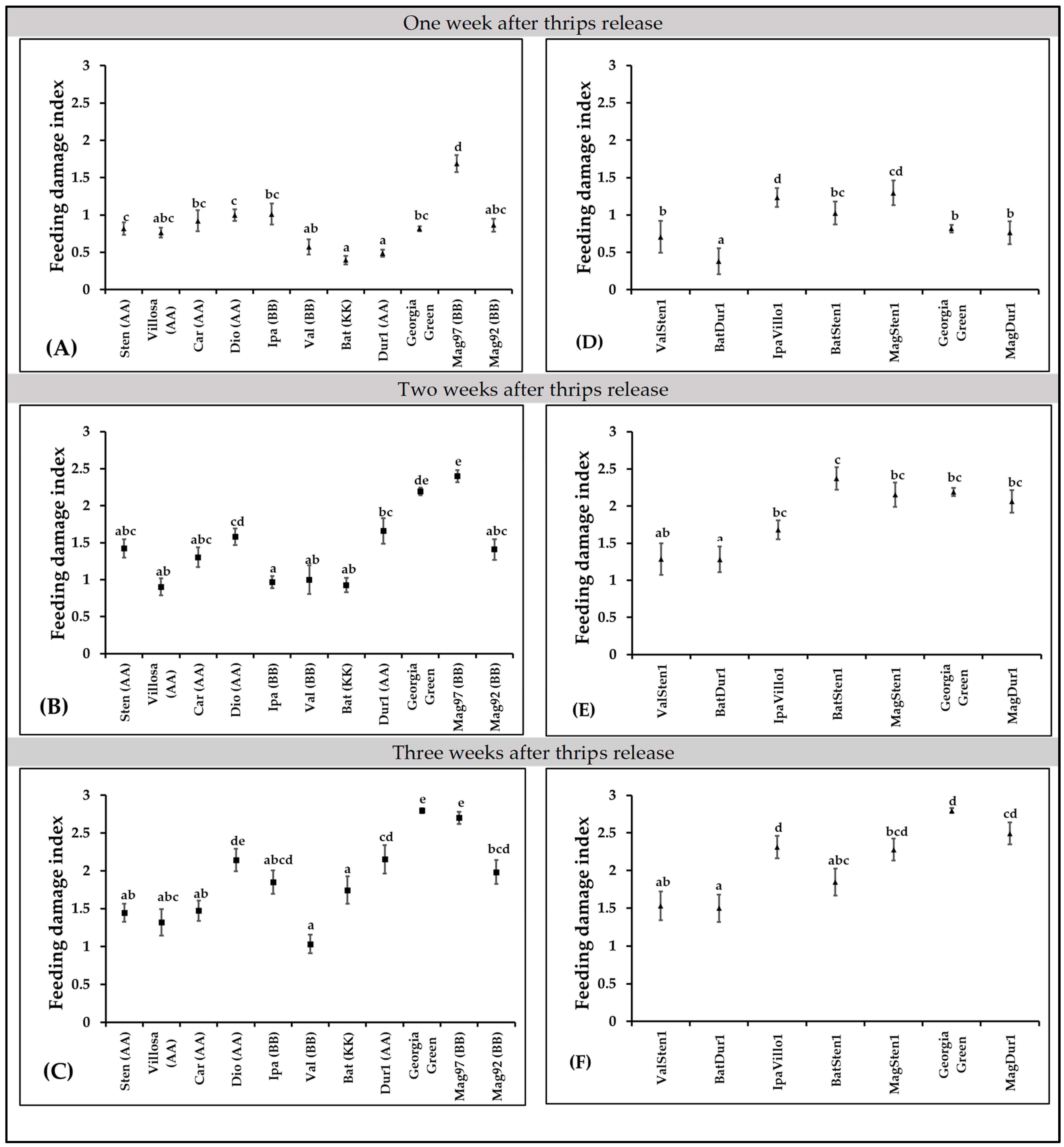
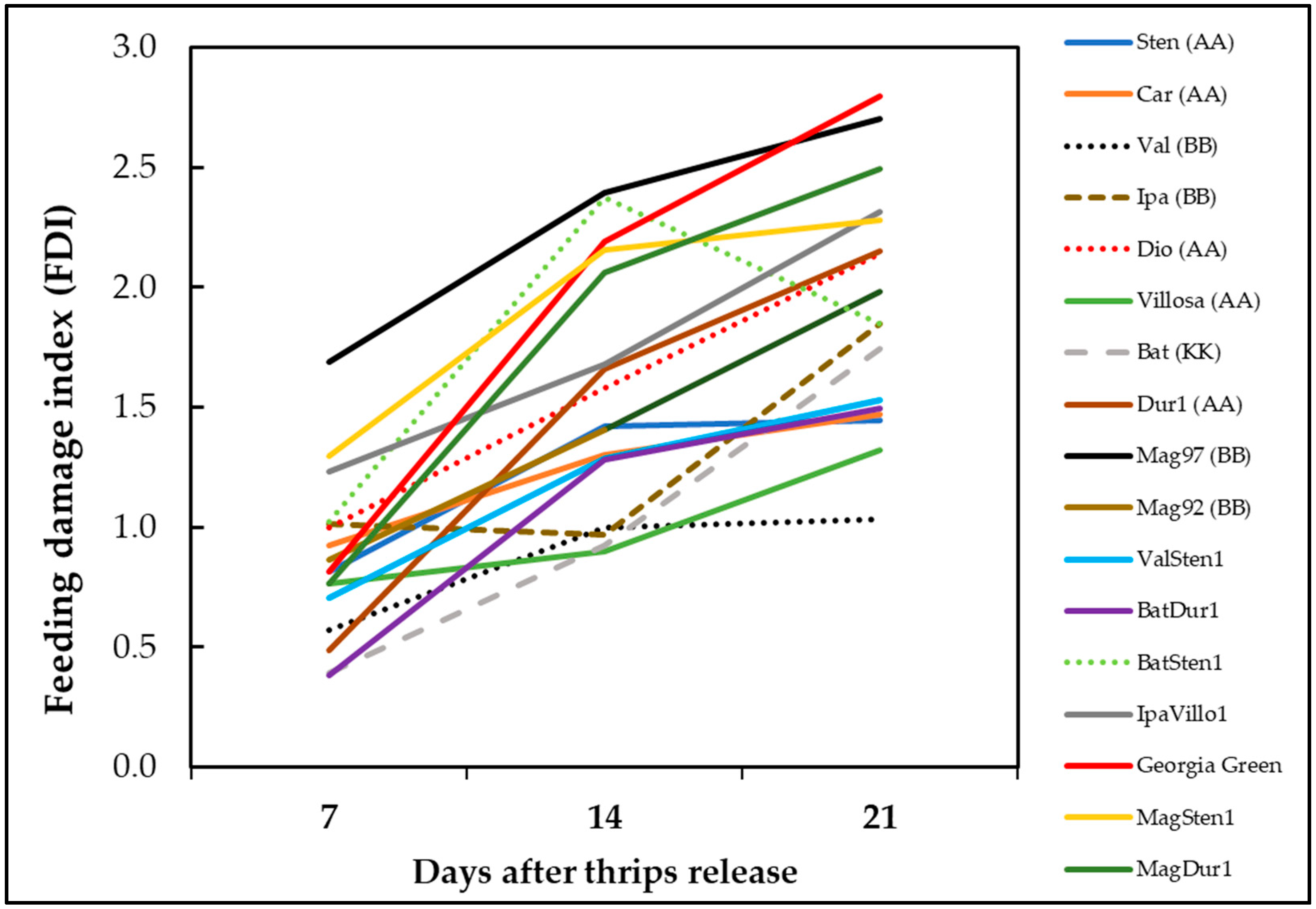
3.4. Thrips Feeding Damage Index
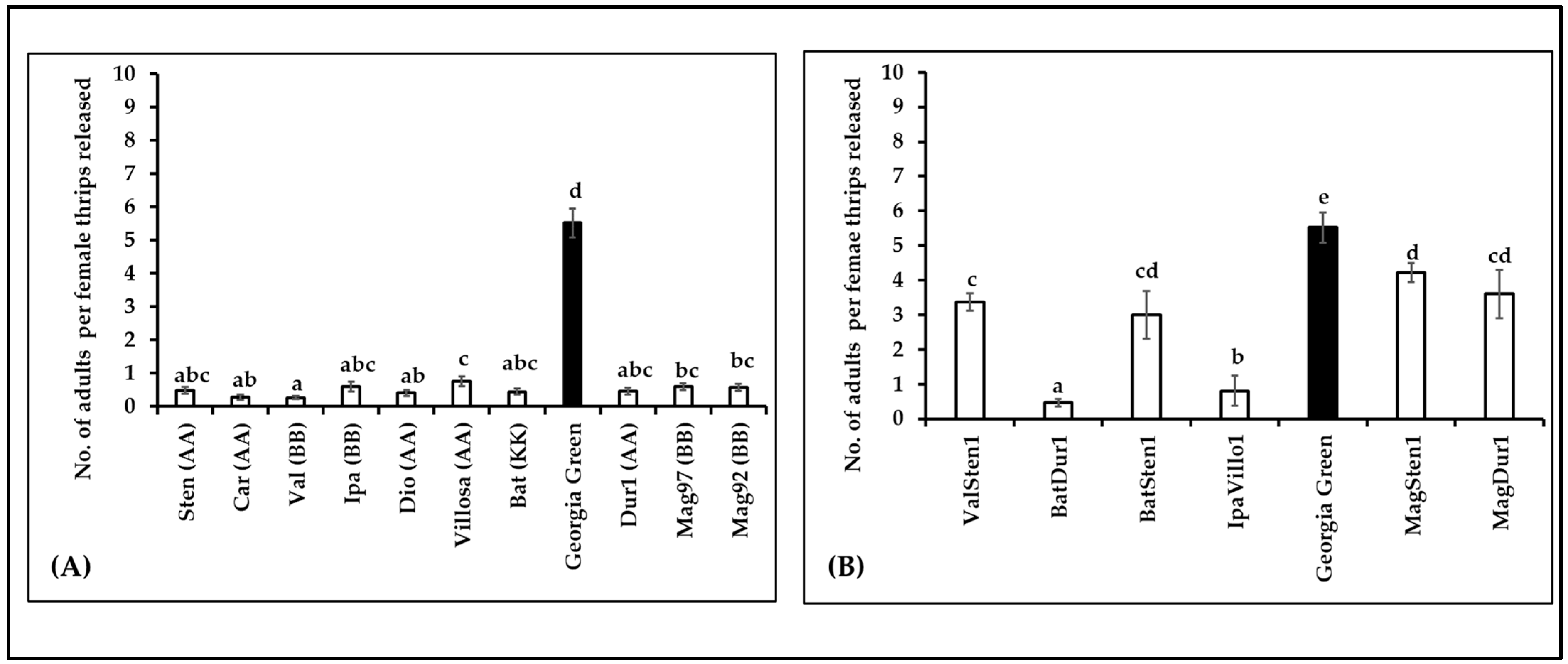
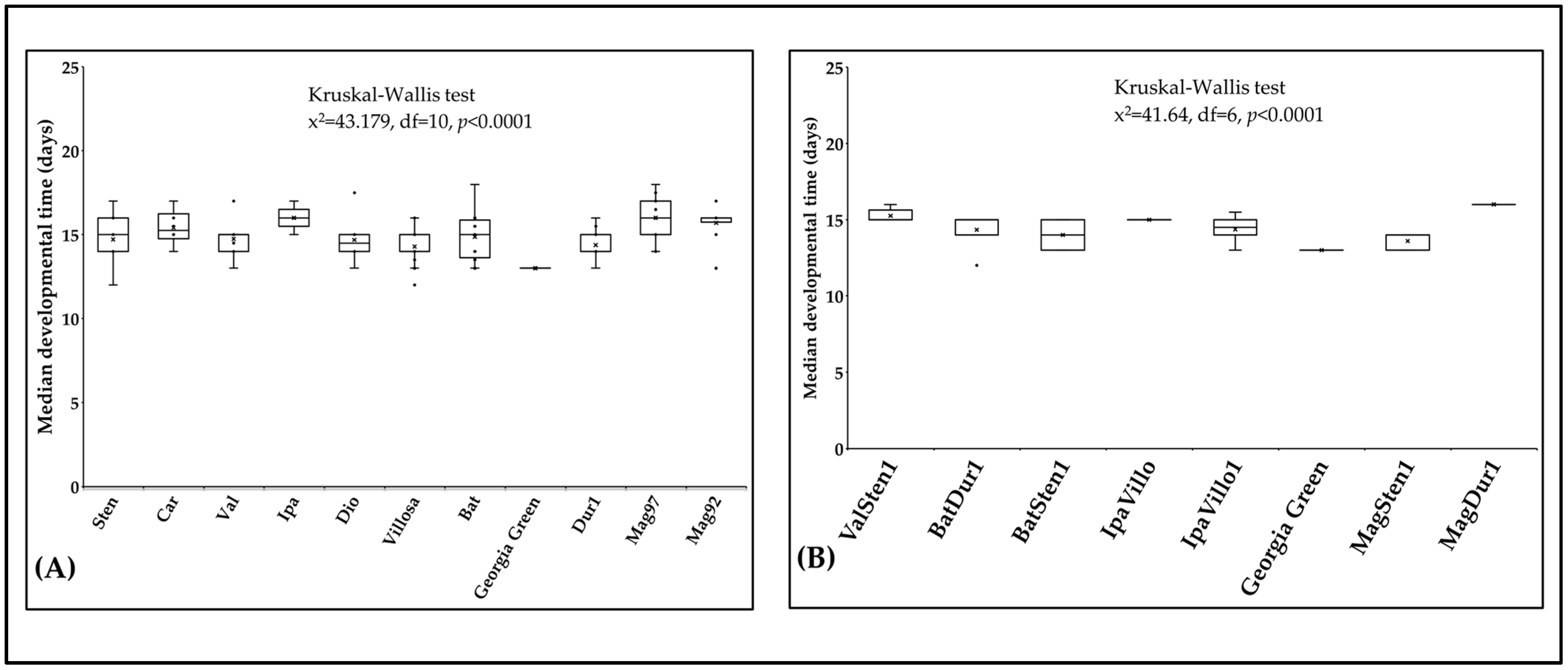
3.5. Thrips Fitness
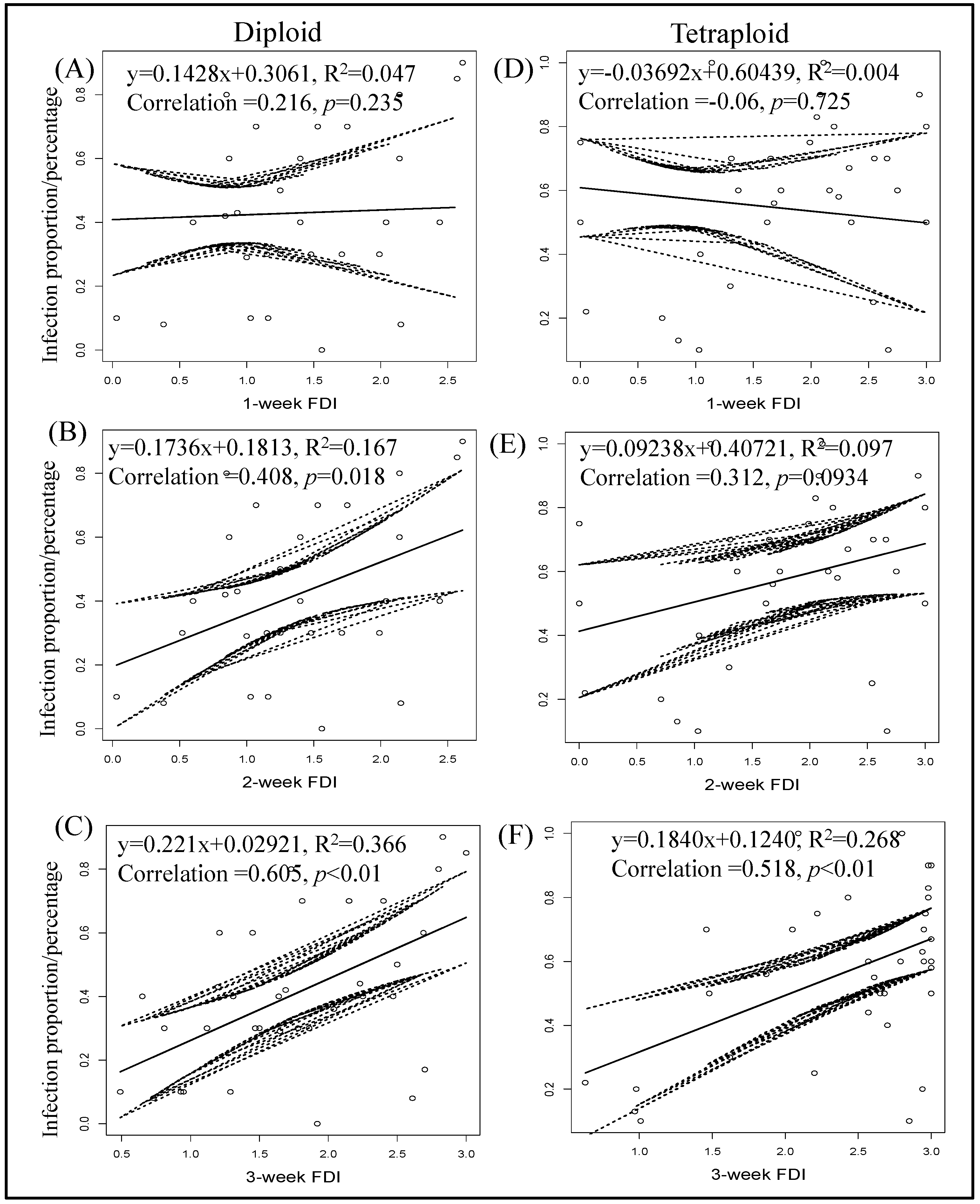
3.6. Correlation between Final Percent TSWV Infection and Thrips Feeding Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- German, T.L.; Ullman, D.E.; Moyer, J.W. Tospoviruses: Diagnosis, Molecular Biology, Phylogeny, and Vector Relationships. Annu. Rev. Phytopathol. 1992, 30, 315–348. [Google Scholar] [CrossRef] [PubMed]
- Chaisuekul, C.; Riley, D.G.; Pappu, H.R. Transmission of Tomato Spotted Wilt Virus to Tomato Plants of Different Ages1. J. Entomol. Sci. 2003, 38, 127–136. [Google Scholar] [CrossRef]
- Groves, R.L.; Walgenbach, J.F.; Moyer, J.W.; Kennedy, G.G. Overwintering of Frankliniella Fusca (Thysanoptera: Thripidae) on Winter Annual Weeds Infected with Tomato Spotted Wilt Virus and Patterns of Virus Movement between Susceptible Weed Hosts. Phytopathology 2001, 91, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R. Temporal Relationship of Thrips Populations to Tomato Spotted Wilt Incidence in Tomato in the Field. J. Entomol. Sci. 2012, 47, 65–75. [Google Scholar] [CrossRef]
- Todd, J.W.; Culbreath, A.K.; Chamberlin, J.R.; Beshear, R.J.; Mullinix, B.G. Colonization and Population Dynamics of Thrips in Peanuts in the Southern United States. In Thrips Biology and Management; Parker, B.L., Skinner, M., Lewis, T., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 453–460. [Google Scholar]
- Culbreath, A.K.; Todd, J.W.; Brown, S.L. Epidemiology and Management of Tomato Spotted Wilt in Peanut. Annu. Rev. Phytopathol. 2003, 41, 53–75. [Google Scholar] [CrossRef]
- Latora, A.G.; Lai, P.C.; Chen, Y.J.; Gautam, S.; Abney, M.R.; Srinivasan, R. Frankliniella Fusca (Thysanoptera: Thripidae), The Vector of Tomato Spotted Wilt Orthotospovirus Infecting Peanut in the Southeastern United States. J. Integr. Pest Manag. 2022, 13, 3. [Google Scholar] [CrossRef]
- Halliwell, R.S.; Philley, G. Spotted Wilt of Peanuts in Texas. Plant Dis. Rep. 1974, 58, 23–25. [Google Scholar]
- Bertrand, P.F. 1997 Georgia Plant Disease Loss Estimates; The University of Georgias: Athens, GA, USA, 1998; Volume 81. [Google Scholar]
- Clevenger, J.; Chu, Y.; Chavarro, C.; Agarwal, G.; Bertioli, D.J.; Leal-Bertioli, S.C.M.; Pandey, M.K.; Vaughn, J.; Abernathy, B.; Barkley, N.A.; et al. Genome-Wide SNP Genotyping Resolves Signatures of Selection and Tetrasomic Recombination in Peanut. Mol. Plant 2017, 10, 309–322. [Google Scholar] [CrossRef]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R.; Diffie, S. Thrips Vectors of Tospoviruses. J. Integr. Pest Manag. 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Srinivasan, R.; Abney, M.R.; Culbreath, A.K.; Kemerait, R.C.; Tubbs, R.S.; Monfort, W.S.; Pappu, H.R. Three Decades of Managing Tomato Spotted Wilt Virus in Peanut in Southeastern United States. Virus Res. 2017, 241, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Branch, W.D. Registration of ‘Georgia Green’ Peanut. Crop Sci. 1996, 36, 806. [Google Scholar] [CrossRef]
- López, C.; Aramburu, J.; Galipienso, L.; Soler, S.; Nuez, F.; Rubio, L. Evolutionary Analysis of Tomato Sw-5 Resistance-Breaking Isolates of Tomato Spotted Wilt Virus. J. Gen. Virol. 2011, 92, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Ciuffo, M.; Pacifico, D.; Turina, M. Evidence That the Nonstructural Protein of Tomato Spotted Wilt Virus Is the Avirulence Determinant in the Interaction with Resistant Pepper Carrying the Tsw Gene. Mol. Plant-Microbe Interact. 2007, 20, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.G.; Reitz, S.R.; Perry, K.L.; Adkins, S. A Natural M RNA Reassortant Arising from Two Species of Plant- and Insect-Infecting Bunyaviruses and Comparison of Its Sequence and Biological Properties to Parental Species. Virology 2011, 413, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Sundaraj, S.; Culbreath, A.K.; Riley, D.G.; Abney, M.R.; Srinivasan, R. Effects of Thrips Density, Mode of Inoculation, and Plant Age on Tomato Spotted Wilt Virus Transmission in Peanut Plants. Environ. Entomol. 2015, 44, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Sundaraj, S.; Srinivasan, R.; Culbreath, A.K.; Riley, D.G.; Pappu, H.R. Host Plant Resistance against Tomato Spotted Wilt Virus in Peanut (Arachis hypogaea) and Its Impact on Susceptibility to the Virus, Virus Population Genetics, and Vector Feeding Behavior and Survival. Phytopathology 2014, 104, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Culbreath, A.K.; Srinivasan, R. Epidemiology of Spotted Wilt Disease of Peanut Caused by Tomato Spotted Wilt Virus in the Southeastern U.S. Virus Res. 2011, 159, 101–109. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Santos, S.P.; Dantas, K.M.; Inglis, P.W.; Nielen, S.; Araujo, A.C.G.; Silva, J.P.; Cavalcante, U.; Guimaraes, P.M.; Brasileiro, A.C.M.; et al. Arachis Batizocoi: A Study of Its Relationship to Cultivated Peanut (A. hypogaea) and Its Potential for Introgression of Wild Genes into the Peanut Crop Using Induced Allotetraploids. Ann. Bot. 2015, 115, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Cason, J.M.; Simpson, C.E.; Burow, M.D.; Tallury, S.; Pham, H.; Ravelombola, S.W. Use of Wild and Exotic Germplasm for Resistance in Peanut. J. Plant Regist. 2023, 17, 1–25. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Seijo, G.; Freitas, F.O.; Valls, J.F.M.; Leal-Bertioli, S.C.M.; Moretzsohn, M.C. An Overview of Peanut and Its Wild Relatives. Plant Genet. Resour. 2011, 9, 134–149. [Google Scholar] [CrossRef]
- Isleib, T.G.; Holbrook, C.C.; Gorbet, D.W. Use of Plant Introductions in Peanut Improvement. Peanut Sci. 2001, 28, 96–113. [Google Scholar] [CrossRef]
- Stalker, H.T. Utilizing Wild Species for Peanut Improvement. Crop Sci. 2017, 57, 1102–1120. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Cavalcante, U.; Gouvea, E.G.; Ballén-Taborda, C.; Shirasawa, K.; Guimarães, P.M.; Jackson, S.A.; Bertioli, D.J.; Moretzsohn, M.C. Identification of QTLs for Rust Resistance in the Peanut Wild Species Arachis Magna and the Development of KASP Markers for Marker-Assisted Selection. G3 Genes Genomes Genet. 2015, 5, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Stalker, H.T.; Tallury, S.P.; Ozias-Akins, P.; Bertioli, D.; Bertioli, S.C.L. The Value of Diploid Peanut Relatives for Breeding and Genomics. Peanut Sci. 2013, 40, 70–88. [Google Scholar] [CrossRef]
- Lyerly, J.H.; Stalker, H.T.; Moyer, J.W.; Hoffman’, K.; Asst, I. Evaluation of Arachis Species for Resistance to Tomato Spotted Wilt Virus. Peanut Sci. 2002, 29, 79–84. [Google Scholar] [CrossRef]
- Ballén-Taborda, C.; Chu, Y.; Ozias-Akins, P.; Timper, P.; Holbrook, C.C.; Jackson, S.A.; Bertioli, D.J.; Leal-Bertioli, S.C.M. A New Source of Root-Knot Nematode Resistance from Arachis Stenosperma Incorporated into Allotetraploid Peanut (Arachis hypogaea). Sci. Rep. 2019, 9, 17702. [Google Scholar] [CrossRef]
- Chu, Y.; Stalker, H.T.; Marasigan, K.; Levinson, C.M.; Gao, D.; Bertioli, D.J.; Leal-Bertioli, S.C.M.; Holbrook, C.C.; Jackson, S.A.; Ozias-Akins, P. Registration of Three Peanut Allotetraploid Interspecific Hybrids Resistant to Late Leaf Spot Disease and Tomato Spotted Wilt. J. Plant Regist. 2021, 15, 562–572. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Ozias-Akins, P.; Chu, Y.; Culbreath, A.K.; Kvien, C.K.; Brenneman, T.B. Registration of ‘TifNV-High O/L’ Peanut. J. Plant Regist. 2017, 11, 228–230. [Google Scholar] [CrossRef]
- Isleib, T.G.; Rice, P.W.; Mozingo, R.W.; Copeland, S.C.; Graeber, J.B.; Shew, B.B.; Smith, D.L.; Melouk, H.A.; Stalker, H.T. Registration of N96076L Peanut Germplasm Line. Crop Sci. 2006, 46, 2329–2330. [Google Scholar] [CrossRef]
- Simpson, C.E.; Starr, J.L.; Church, G.T.; Burow, M.D.; Paterson, A.H. Registration of’NemaTAM’peanut. (Registrations of Cultivars). Crop Sci. 2003, 43, 1561–1562. [Google Scholar] [CrossRef]
- Angelella, G.M.; Riley, D.G. Effects of Pine Pollen Supplementation in an Onion Diet on Frankliniella Fusca Reproduction. Environ. Entomol. 2010, 39, 505–512. [Google Scholar] [CrossRef]
- Shrestha, A.; Srinivasan, R.; Sundaraj, S.; Culbreath, A.K.; Riley, D.G. Second Generation Peanut Genotypes Resistant to Thrips-Transmitted Tomato Spotted Wilt Virus Exhibit Tolerance Rather than True Resistance and Differentially Affect Thrips Fitness. J. Econ. Entomol. 2013, 106, 587–596. [Google Scholar] [CrossRef]
- Marasigan, K.M. Alternatives to Carbamate and Organophosphate Insecticides, Cultural Tactics, and Ecological Factors That Affect Tomato Spotted Wilt Virus Epidemics in Peanut. Master’s Thesis, University of Georgia, Athens, GA, USA, 2014. [Google Scholar]
- Leal-Bertioli, S.C.M.; Moretzsohn, M.C.; Santos, S.P.; Brasileiro, A.C.M.; Guimarães, P.M.; Bertioli, D.J.; Araujo, A.C.G. Phenotypic Effects of Allotetraploidization of Wild Arachis and Their Implications for Peanut Domestication. Am. J. Bot. 2017, 104, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Araujo, A.C.G.; Nascimento, E.F.M.B.; Chavarro, M.C.; Xia, H.; Jackson, S.A.; Bertioli, D.J.; Leal-Bertioli, S.C.M. ValSten: A New Wild Species Derived Allotetraploid for Increasing Genetic Diversity of the Peanut Crop (Arachis hypogaea L.). Genet. Resour. Crop Evol. 2021, 68, 1471–1485. [Google Scholar] [CrossRef]
- Lai, P.C.; Abney, M.R.; Chen, Y.J.; Bag, S.; Srinivasan, R. Discrepancies in Serology-Based and Nucleic Acid-Based Detection and Quantitation of Tomato Spotted Wilt Orthotospovirus in Leaf and Root Tissues from Symptomatic and Asymptomatic Peanut Plants. Pathogens 2021, 10, 1476. [Google Scholar] [CrossRef]
- Rotenberg, D.; Kumar, N.K.K.; Ullman, D.E.; Montero-Astúa, M.; Willis, D.K.; German, T.L.; Whitfield, A.E. Variation in Tomato Spotted Wilt Virus Titer in Frankliniella Occidentalis and Its Association with Frequency of Transmission. Phytopathology 2009, 99, 404–410. [Google Scholar] [CrossRef]
- Shrestha, A.; Srinivasan, R.; Riley, D.G.; Culbreath, A.K. Direct and Indirect Effects of a Thrips-Transmitted Tospovirus on the Preference and Fitness of Its Vector, Frankliniella Fusca. Entomol. Exp. Appl. 2012, 145, 260–271. [Google Scholar] [CrossRef]
- Maris, P.C.; Joosten, N.N.; Peters, D.; Goldbach, R.W. Thrips Resistance in Pepper and Its Consequences for the Acquisition and Inoculation of Tomato Spotted Wilt Virus by the Western Flower Thrips. Phytopathology 2003, 93, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Munger, F. A Method for Rearing Citrus Thrips in the Laboratory. J. Econ. Entomol. 1942, 35, 373–375. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R--project.org (accessed on 1 May 2023).
- Leal-Bertioli, S.C.; Farias, M.P.; Silva, P.T.; Guimarães, P.M.; Brasileiro, A.C.M.; Bertioli, D.J.; Araujo, A.C.G. Ultrastructure of the Initial Interaction of Puccinia Arachidis and Cercosporidium Personatum with Leaves of Arachis Hypogaea and Arachis Stenosperma. J. Phytopathol. 2010, 158, 792–796. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Moretzsohn, M.C.; Roberts, P.A.; Ballén-Taborda, C.; Borba, T.C.O.; Valdisser, P.A.; Vianello, R.P.; Araújo, A.C.G.; Guimarães, P.M.; Bertioli, D.J. Genetic Mapping of Resistance to Meloidogyne Arenaria in Arachis Stenosperma: A New Source of Nematode Resistance for Peanut. G3 Genes Genomes Genet. 2016, 6, 377–390. [Google Scholar] [CrossRef]
- Proite, K.; Carneiro, R.; Falcão, R.; Gomes, A.; Leal-Bertioli, S.; Guimarães, P.; Bertioli, D. Post-Infection Development and Histopathology of Meloidogyne Arenaria Race 1 on Arachis spp. Plant Pathol. 2008, 57, 974–980. [Google Scholar] [CrossRef]
- Lai, P.-C. Evaluation of Cultural Tactics, Insecticides, and Peanut Genotypes for Thrips and Spotted Wilt Disease Management in Peanut. Master’s Thesis, University of Georgia, Athens, GA, USA, 2015; pp. 1–190. [Google Scholar]
- Lai, P.C.; Abney, M.R.; Bag, S.; Culbreath, A.K.; Srinivasan, R. Impact of Host Resistance to Tomato Spotted Wilt Orthotospovirus in Peanut Cultivars on Virus Population Genetics and Thrips Fitness. Pathogens 2021, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Clevenger, J.; Kale, S.M.; Wang, H.; Pandey, M.K.; Choudhary, D.; Yuan, M.; Wang, X.; Culbreath, A.K.; Holbrook, C.C.; et al. A Recombination Bin-Map Identified a Major QTL for Resistance to Tomato Spotted Wilt Virus in Peanut (Arachis hypogaea). Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Khera, P.; Pandey, M.K.; Wang, H.; Feng, S.; Qiao, L.; Culbreath, A.K.; Kale, S.; Wang, J.; Holbrook, C.C.; Zhuang, W.; et al. Mapping Quantitative Trait Loci of Resistance to Tomato Spotted Wilt Virus and Leaf Spots in a Recombinant Inbred Line Population of Peanut (Arachis hypogaea L.) from SunOleic 97R and NC94022. PLoS ONE 2016, 11, e0158452. [Google Scholar] [CrossRef]
- Pandey, M.K.; Wang, H.; Khera, P.; Vishwakarma, M.K.; Kale, S.M.; Culbreath, A.K.; Holbrook, C.C.; Wang, X.; Varshney, R.K.; Guo, B. Genetic Dissection of Novel QTLs for Resistance to Leaf Spots and Tomato Spotted Wilt Virus in Peanut (Arachis hypogaea L.). Front. Plant Sci. 2017, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Moury, B.; Palloix, A.; Gebre Selassie, K.; Marchoux, G. Hypersensitive Resistance to Tomato Spotted Wilt Virus in Three Capsicum Chinense Accessions Is Controlled by a Single Gene and Is Overcome by Virulent Strains. Euphytica 1997, 94, 45–52. [Google Scholar] [CrossRef]
- Branch, W.D.; Culbreath, A.K. Stability of TSWV General Field Resistance in the “Georgia Green” Peanut Cultivar. Plant Health Prog. 2015, 16, 95–99. [Google Scholar] [CrossRef]
- Branch, W.D.; Culbreath, A.K. Transgressive Segregation and Long-Term Consistency for High TSWV Field Resistance in the “Georgia-06G” Peanut Cultivar. Plant Health Prog. 2018, 19, 201–206. [Google Scholar] [CrossRef]
- Catto, M.A.; Shrestha, A.; Abney, M.R.; Champagne, D.E.; Culbreath, A.K.; Leal-Bertioli, S.C.M.; Hunt, B.G.; Srinivasan, R. Defense-Related Gene Expression Following an Orthotospovirus Infection Is Influenced by Host Resistance in Arachis Hypogaea. Viruses 2021, 13, 1303. [Google Scholar] [CrossRef]
- Stalker, H.T.; Campbell, W.V. Resistance of Wild Species of Peanut to an Insect Complex1. Peanut Sci. 1983, 10, 30–33. [Google Scholar] [CrossRef]
- Yang, G.; Espelie, K.E.; Todd, J.W.; Culbreath, A.K.; Pittman, R.N.; Demski, J.W. Cuticular Lipids from Wild and Cultivated Peanuts and the Relative Resistance of These Peanut Species to Fall Armyworm and Thrips. J. Agric. Food Chem. 1993, 41, 814–818. [Google Scholar] [CrossRef]
- Michelotto, M.D.; De Godoy, I.J.; Pirotta, M.Z.; Dos Santos, J.F.; Finoto, E.L.; Favero, A.P. Resistance to Thrips (Enneothrips flavens) in Wild and Amphidiploid Arachis Species. PLoS ONE 2017, 12, e0176811. [Google Scholar] [CrossRef] [PubMed]
- Mouden, S.; Leiss, K.A. Host Plant Resistance to Thrips (Thysanoptera: Thripidae)—Current State of Art and Future Research Avenues. Curr. Opin. Insect Sci. 2021, 45, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kortbeek, R.W.J.; Galland, M.D.; Muras, A.; van der Kloet, F.M.; André, B.; Heilijgers, M.; van Hijum, S.A.F.T.; Haring, M.A.; Schuurink, R.C.; Bleeker, P.M. Natural Variation in Wild Tomato Trichomes; Selecting Metabolites That Contribute to Insect Resistance Using a Random Forest Approach. BMC Plant Biol. 2021, 21, 315. [Google Scholar] [CrossRef]
- Margaria, P.; Ciuffo, M.; Rosa, C.; Turina, M. Evidence of a Tomato Spotted Wilt Virus Resistance-Breaking Strain Originated through Natural Reassortment between Two Evolutionary-Distinct Isolates. Virus Res. 2015, 196, 157–161. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild Diploid | Plant ID | Accession No. (USDA No.) | Genome Type | Collection Site |
|---|---|---|---|---|
| A. batizocoi | Bat | K9484 (PI 298639) | KK | Parape, Bolivia |
| A. cardenasii | Car | GKP10017 (PI262141) | AA | Robore, Bolivia |
| A. diogoi | Dio | V10602 (PI 276235) | AA | Paraguay |
| A. duranensis | Dur1 | V14167 | AA | Salta, Argentina |
| A. ipaënsis | Ipa | K30076 (PI 468322) | BB | Gran Chao, Bolivia |
| A. magna | Mag92 | K30092 (PI 468337) | BB | Bolivia |
| A. magna | Mag97 | K30097 (PI468340) | BB | Santa Cruz, Bolivia |
| A. stenosperma | Sten | V10309 (PI 666100) | AA | Mato Grosso, Brazil |
| A. valida | Val | GK30011 (PI 468154) | BB | Mato Grosso, Brazil |
| A. villosa | Villo | V12812 | AA | Bella Union, Uruguay |
| Induced Allotetraploids | Plant ID | Genome Type | Collection/Registration |
|---|---|---|---|
| A. batizocoi K9484 × A. duranensis V14167 | BatDur1 | KKAA | [20] |
| A. batizocoi K9484 × A. stenosperma V10309 | BatSten1 (PI 695418) | KKAA | [20] |
| A. ipaënsis K30076 × A. villosa V12812 | IpaVillo1 | BBAA | [36] |
| A. magna K30097 × A. duranensis V14167 | MagDur1 | BBAA | Unpublished |
| A. magna K30097 × A. stenosperma V10309 | MagSten1 (PI 695417) | BBAA | [36] |
| A. valida GK30011 × A. stenosperma V10309 | ValSten1 (PI 695393) | BBAA | [29,37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-J.; Pandey, S.; Catto, M.; Leal-Bertioli, S.; Abney, M.R.; Bag, S.; Hopkins, M.; Culbreath, A.; Srinivasan, R. Evaluation of Wild Peanut Species and Their Allotetraploids for Resistance against Thrips and Thrips-Transmitted Tomato Spotted Wilt Orthotospovirus (TSWV). Pathogens 2023, 12, 1102. https://doi.org/10.3390/pathogens12091102
Chen Y-J, Pandey S, Catto M, Leal-Bertioli S, Abney MR, Bag S, Hopkins M, Culbreath A, Srinivasan R. Evaluation of Wild Peanut Species and Their Allotetraploids for Resistance against Thrips and Thrips-Transmitted Tomato Spotted Wilt Orthotospovirus (TSWV). Pathogens. 2023; 12(9):1102. https://doi.org/10.3390/pathogens12091102
Chicago/Turabian StyleChen, Yi-Ju, Sudeep Pandey, Michael Catto, Soraya Leal-Bertioli, Mark R. Abney, Sudeep Bag, Mark Hopkins, Albert Culbreath, and Rajagopalbabu Srinivasan. 2023. "Evaluation of Wild Peanut Species and Their Allotetraploids for Resistance against Thrips and Thrips-Transmitted Tomato Spotted Wilt Orthotospovirus (TSWV)" Pathogens 12, no. 9: 1102. https://doi.org/10.3390/pathogens12091102