
Monocytes from Uninfected Neonates Born to Trypanosoma cruzi-Infected Mothers Display Upregulated Capacity to Produce TNF-α and to Control Infection in Association with Maternally Transferred Antibodies
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Blood Obtention
2.2. Purification of Cord Blood Mononuclear Cells and Monocytes
2.3. T. cruzi Infection of Monocytes
2.4. TNF-α Production by Monocytes
2.5. Phenotypic Characterization of Monocytes by Flow Cytometry
2.6. Depletion of T. cruzi-Specific Antibodies by Affinity Chromatography
3. Results
3.1. Monocytes from M+B- Newborns Are Primed to Produce Higher TNF-α Levels
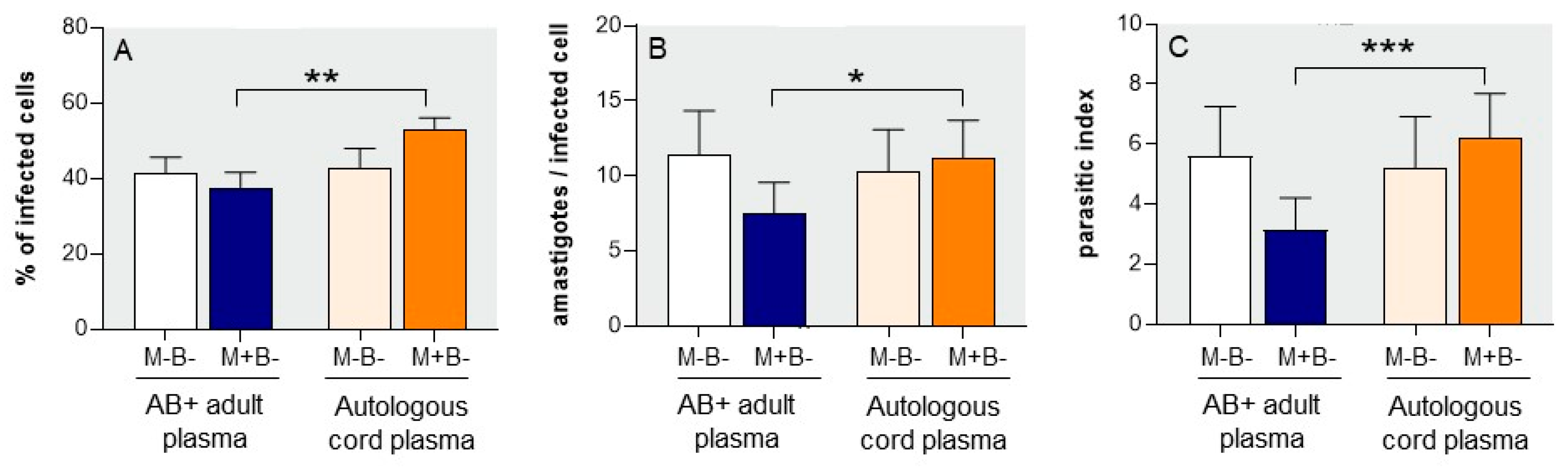
3.2. Susceptibility of M+B- Monocytes to T. cruzi Infection
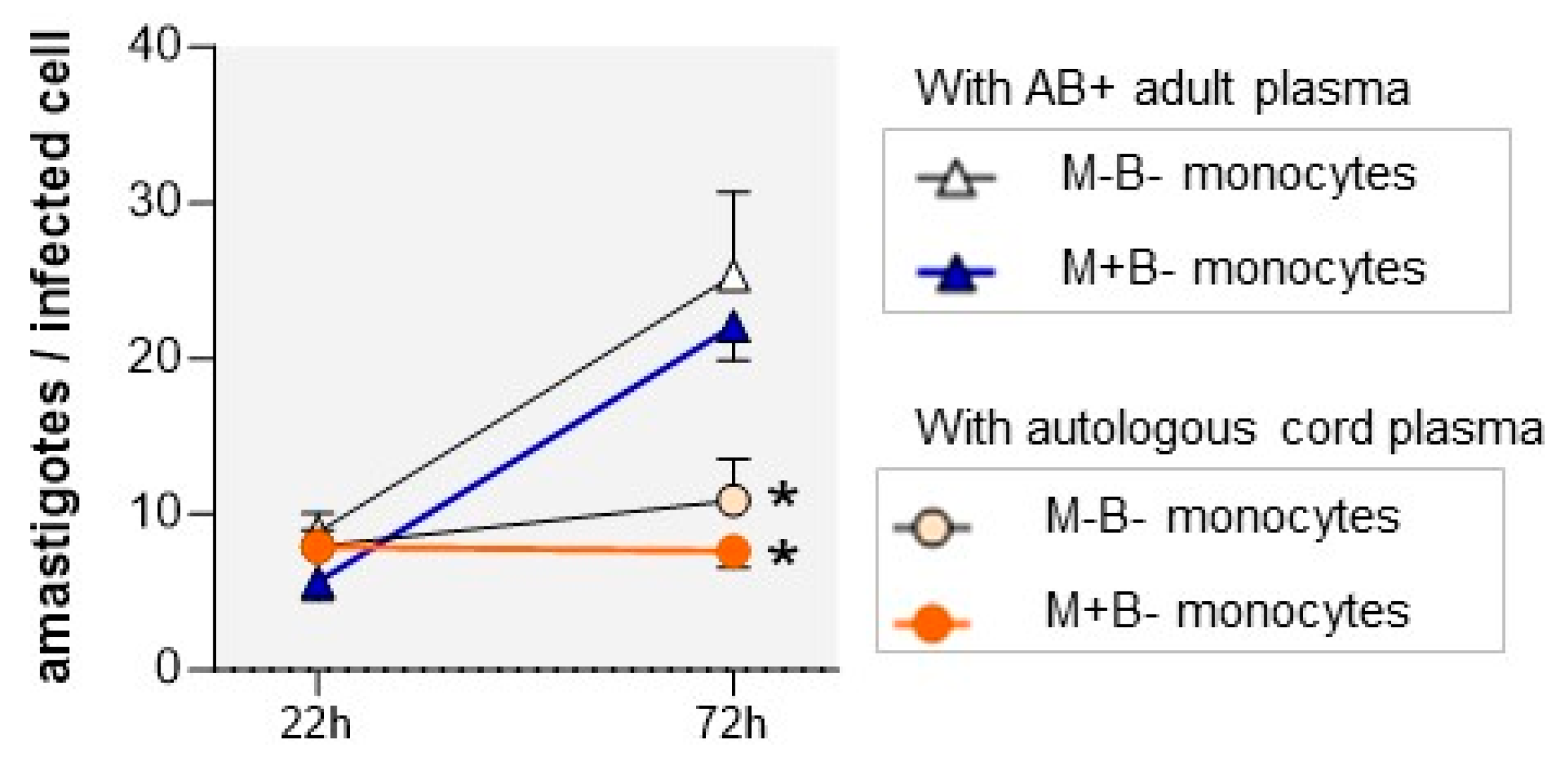
3.3. Autologous Cord Plasma Limits Intracellular T. cruzi Multiplication
3.4. M+B- Monocytes Are Occasionally Able to Completely Control Intracellular Infection
3.5. T. cruzi–Specific Antibodies Present in the Plasma of M+B- Newborns Improve the Control of the Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Echavarría, N.G.; Echeverría, L.E.; Stewart, M.; Gallego, C.; Saldarriaga, C. Chagas Disease: Chronic Chagas Cardiomyopathy. Curr. Probl. Cardiol. 2021, 46, 100507. [Google Scholar] [CrossRef]
- Gómez-Ochoa, S.A.; Rojas, L.Z.; Echeverría, L.E.; Muka, T.; Franco, O.H. Global, Regional, and National Trends of Chagas Disease from 1990 to 2019: Comprehensive Analysis of the Global Burden of Disease Study. Glob. Heart 2022, 17, 59. [Google Scholar] [CrossRef]
- Carlier, Y.; Truyens, C. Congenital Chagas Disease as an Ecological Model of Interactions between Trypanosoma cruzi Parasites, Pregnant Women, Placenta and Fetuses. Acta Trop. 2015, 151, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Kemmerling, U.; Osuna, A.; Schijman, A.; Truyens, C. Trypanosoma cruzi Infection, Maternal-Fetal Interactions and Congenital Transmission. Front. Microbiol. 2019, 10, 1854. [Google Scholar] [CrossRef]
- Carlier, Y.; Altcheh, J.; Angheben, A.; Freilij, H.; Luquetti, A.O.; Schijman, A.G.; Segovia, M.; Wagner, N.; Albajar Vinas, P. Congenital Chagas Disease: Updated Recommendations for Prevention, Diagnosis, Treatment, and Follow-up of Newborns and Siblings, Girls, Women of Childbearing Age, and Pregnant Women. PLoS Negl. Trop. Dis. 2019, 13, e0007694. [Google Scholar] [CrossRef] [PubMed]
- Carlier, Y.; Truyens, C. 23—Maternal–Fetal Transmission of Trypanosoma cruzi. In American Trypanosomiasis Chagas Disease, 2nd ed.; Telleria, J., Tibayrenc, M., Eds.; Elsevier: London, UK, 2017; pp. 517–559. ISBN 978-0-12-801029-7. [Google Scholar]
- Torrico, F.; Alonso-Vega, C.; Suarez, E.; Rodriguez, P.; Torrico, M.-C.; Dramaix, M.; Truyens, C.; Carlier, Y. Maternal Trypanosoma cruzi Infection, Pregnancy Outcome, Morbidity, and Mortality of Congenitally Infected and Non-Infected Newborns in Bolivia. Am. J. Trop. Med. Hyg. 2004, 70, 201–209. [Google Scholar] [CrossRef]
- Carlier, Y.; Truyens, C.; Muraille, E. Is Antibody-Dependent Enhancement of Trypanosoma cruzi Infection Contributing to Congenital/Neonatal Chagas Disease? Front. Immunol. 2021, 12, 723516. [Google Scholar] [CrossRef]
- Buekens, P.; Cafferata, M.L.; Alger, J.; Althabe, F.; Belizán, J.M.; Bustamante, N.; Carlier, Y.; Ciganda, A.; Del Cid, J.H.; Dumonteil, E.; et al. Congenital Transmission of Trypanosoma cruzi in Argentina, Honduras, and Mexico: An Observational Prospective Study. Am. J. Trop. Med. Hyg. 2018, 98, 478–485. [Google Scholar] [CrossRef]
- Herrera, C.; Truyens, C.; Dumonteil, E.; Alger, J.; Sosa-Estani, S.; Cafferata, M.L.; Gibbons, L.; Ciganda, A.; Matute, M.L.; Zuniga, C.; et al. Phylogenetic Analysis of Trypanosoma cruzi from Pregnant Women and Newborns from Argentina, Honduras, and Mexico Suggests an Association of Parasite Haplotypes with Congenital Transmission of the Parasite. J. Mol. Diagn. 2019, 21, 1095–1105. [Google Scholar] [CrossRef]
- Hermann, E.; Truyens, C.; Alonso-Vega, C.; Rodriguez, P.; Berthe, A.; Torrico, F.; Carlier, Y. Congenital Transmission of Trypanosoma cruzi Is Associated with Maternal Enhanced Parasitemia and Decreased Production of Interferon- Gamma in Response to Parasite Antigens. J. Infect. Dis. 2004, 189, 1274–1281. [Google Scholar] [CrossRef]
- Sánchez Negrette, O.; Mora, M.C.; Basombrío, M.A. High Prevalence of Congenital Trypanosoma cruzi Infection and Family Clustering in Salta, Argentina. Pediatrics 2005, 115, e668–e672. [Google Scholar] [CrossRef] [PubMed]
- Lykens, J.E.; Terrell, C.E.; Zoller, E.E.; Divanovic, S.; Trompette, A.; Karp, C.L.; Aliberti, J.; Flick, M.J.; Jordan, M.B. Mice with a Selective Impairment of IFN-Gamma Signaling in Macrophage Lineage Cells Demonstrate the Critical Role of IFN-Gamma-Activated Macrophages for the Control of Protozoan Parasitic Infections In Vivo. J. Immunol. 2010, 184, 877–885. [Google Scholar] [CrossRef]
- Vekemans, J.; Truyens, C.; Torrico, F.; Solano, M.; Torrico, M.C.; Rodriguez, P.; Alonso-Vega, C.; Carlier, Y. Maternal Trypanosoma cruzi Infection Upregulates Capacity of Uninfected Neonate Cells to Produce Pro- and Anti-Inflammatory Cytokines. Infect. Immun. 2000, 68, 5430–5434. [Google Scholar] [CrossRef]
- Zingales, B.; Miles, M.A.; Campbell, D.A.; Tibayrenc, M.; Macedo, A.M.; Teixeira, M.M.G.; Schijman, A.G.; Llewellyn, M.S.; Lages-Silva, E.; Machado, C.R.; et al. The Revised Trypanosoma cruzi Subspecific Nomenclature: Rationale, Epidemiological Relevance and Research Applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Hermann, E.; Truyens, C.; Alonso-Vega, C.; Even, J.; Rodriguez, P.; Berthe, A.; Gonzalez-Merino, E.; Torrico, F.; Carlier, Y. Human Fetuses Are Able to Mount an Adultlike CD8 T-Cell Response. Blood 2002, 100, 2153–2158. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bowie, A.G. The Family of Five: TIR-Domain-Containing Adaptors in Toll-like Receptor Signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Truyens, C.; Carlier, Y. 25—Protective Host Response to Trypanosoma cruzi and Its Limitations. In American Trypanosomiasis Chagas Disease, 2nd ed.; Telleria, J., Tibayrenc, M., Eds.; Elsevier: London, UK, 2017; pp. 579–604. ISBN 978-0-12-801029-7. [Google Scholar]
- de Lana, M.; de Menezes Machado, E.M. 14—Biology of Trypanosoma cruzi and Biological Diversity. In American Trypanosomiasis; Telleria, J., Tibayrenc, M., Eds.; Elsevier: London, UK, 2010; pp. 339–363. ISBN 978-0-12-384876-5. [Google Scholar]
- Caradonna, K.L.; Engel, J.C.; Jacobi, D.; Lee, C.-H.; Burleigh, B.A. Host Metabolism Regulates Intracellular Growth of Trypanosoma cruzi. Cell Host Microbe 2013, 13, 108–117. [Google Scholar] [CrossRef]
- Almeida, I.C.; Ferguson, M.A.; Schenkman, S.; Travassos, L.R. Lytic Anti-Alpha-Galactosyl Antibodies from Patients with Chronic Chagas’ Disease Recognize Novel O-Linked Oligosaccharides on Mucin-like Glycosyl-Phosphatidylinositol-Anchored Glycoproteins of Trypanosoma cruzi. Biochem. J. 1994, 304 Pt 3, 793–802. [Google Scholar] [CrossRef]
- Galili, U. Anti-Gal: An Abundant Human Natural Antibody of Multiple Pathogeneses and Clinical Benefits. Immunology 2013, 140, 1–11. [Google Scholar] [CrossRef]
- Dauby, N.; Goetghebuer, T.; Kollmann, T.R.; Levy, J.; Marchant, A. Uninfected but Not Unaffected: Chronic Maternal Infections during Pregnancy, Fetal Immunity, and Susceptibility to Postnatal Infections. Lancet Infect. Dis. 2012, 12, 330–340. [Google Scholar] [CrossRef]
- Netea, M.G.; Quintin, J.; van der Meer, J.W.M. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Bekkering, S.; Domínguez-Andrés, J.; Joosten, L.A.B.; Riksen, N.P.; Netea, M.G. Trained Immunity: Reprogramming Innate Immunity in Health and Disease. Annu. Rev. Immunol. 2021, 39, 667–693. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, M.F.; Abu-Raya, B.; Jiang, Y.; Alter, G.; Marchant, A. Transfer of Maternal Immunity and Programming of the Newborn Immune System. Semin. Immunopathol. 2017, 39, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Dauby, N.; Alonso-Vega, C.; Suarez, E.; Flores, A.; Hermann, E.; Córdova, M.; Tellez, T.; Torrico, F.; Truyens, C.; Carlier, Y. Maternal Infection with Trypanosoma cruzi and Congenital Chagas Disease Induce a Trend to a Type 1 Polarization of Infant Immune Responses to Vaccines. PLoS Negl. Trop. Dis. 2009, 3, e571. [Google Scholar] [CrossRef] [PubMed]
- Desale, H.; Buekens, P.; Alger, J.; Cafferata, M.L.; Harville, E.W.; Herrera, C.; Truyens, C.; Dumonteil, E. Epigenetic Signature of Exposure to Maternal Trypanosoma cruzi Infection in Cord Blood Cells from Uninfected Newborns. Epigenomics 2022, 14, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Aguilar, S.; Lambot, M.-A.; Torrico, F.; Alonso-Vega, C.; Córdoba, M.; Suarez, E.; Noël, J.-C.; Carlier, Y. Placental lesions in human Trypanosoma cruzi infection. Rev. Soc. Bras. Med. Trop. 2005, 38 (Suppl. 2), 84–86. [Google Scholar] [PubMed]
- Levy, O.; Wynn, J.L. A Prime Time for Trained Immunity: Innate Immune Memory in Newborns and Infants. Neonatology 2014, 105, 136–141. [Google Scholar] [CrossRef]
- Zhao, S.; Zhong, Y.; Fu, X.; Wang, Y.; Ye, P.; Cai, J.; Liu, Y.; Sun, J.; Mei, Z.; Jiang, Y.; et al. H3K4 Methylation Regulates LPS-Induced Proinflammatory Cytokine Expression and Release in Macrophages. Shock 2019, 51, 401–406. [Google Scholar] [CrossRef]
- Rodrigues, M.M.; Oliveira, A.C.; Bellio, M. The Immune Response to Trypanosoma cruzi: Role of Toll-Like Receptors and Perspectives for Vaccine Development. J. Parasitol. Res. 2012, 2012, 507874. [Google Scholar] [CrossRef]
- Ait Djebbara, S.; Mcheik, S.; Percier, P.; Segueni, N.; Poncelet, A.; Truyens, C. The Macrophage Infectivity Potentiator of Trypanosoma cruzi Induces Innate IFN-γ and TNF-α Production by Human Neonatal and Adult Blood Cells through TLR2/1 and TLR4. Front. Immunol. 2023, 14, 1180900. [Google Scholar] [CrossRef]
- Böhm, S.; Schwab, I.; Lux, A.; Nimmerjahn, F. The Role of Sialic Acid as a Modulator of the Anti-Inflammatory Activity of IgG. Semin. Immunopathol. 2012, 34, 443–453. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Alter, G. The Immunoregulatory Roles of Antibody Glycosylation. Trends Immunol. 2017, 38, 358–372. [Google Scholar] [CrossRef]
- Campetella, O.; Buscaglia, C.A.; Mucci, J.; Leguizamón, M.S. Parasite-Host Glycan Interactions during Trypanosoma cruzi Infection: Trans-Sialidase Rides the Show. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165692. [Google Scholar] [CrossRef]
- Freire-de-Lima, L.; Fonseca, L.M.; Oeltmann, T.; Mendonça-Previato, L.; Previato, J.O. The Trans-Sialidase, the Major Trypanosoma cruzi Virulence Factor: Three Decades of Studies. Glycobiology 2015, 25, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Irvine, E.B.; Alter, G. Understanding the Role of Antibody Glycosylation through the Lens of Severe Viral and Bacterial Diseases. Glycobiology 2020, 30, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, G.R.; Girard, M.C.; Gómez, K.A. The Unsolved Jigsaw Puzzle of the Immune Response in Chagas Disease. Front. Immunol. 2018, 9, 1929. [Google Scholar] [CrossRef]
- Ehrhart, J.; Sanberg, P.R.; Garbuzova-Davis, S. Plasma Derived from Human Umbilical Cord Blood: Potential Cell-Additive or Cell-Substitute Therapeutic for Neurodegenerative Diseases. J. Cell Mol. Med. 2018, 22, 6157–6166. [Google Scholar] [CrossRef] [PubMed]
- de Lana, M.; de Menezes Machado, E.M. 16—Biology of Trypanosoma cruzi and Biological Diversity. In American Trypanosomiasis Chagas Disease, 2nd ed.; Telleria, J., Tibayrenc, M., Eds.; Elsevier: London, UK, 2017; pp. 345–369. ISBN 978-0-12-801029-7. [Google Scholar]
- Ferri, G.; Edreira, M.M. All Roads Lead to Cytosol: Trypanosoma cruzi Multi-Strategic Approach to Invasion. Front. Cell Infect. Microbiol. 2021, 11, 634793. [Google Scholar] [CrossRef]
- Nájera, C.A.; Batista, M.F.; Meneghelli, I.; Bahia, D. Mixed Signals—How Trypanosoma cruzi Exploits Host-Cell Communication and Signaling to Establish Infection. J. Cell Sci. 2021, 134, jcs255687. [Google Scholar] [CrossRef]
- Truyens, C.; Rivera, M.T.; Ouaissi, A.; Carlier, Y. High Circulating Levels of Fibronectin and Antibodies against Its RGD Adhesion Site during Mouse Trypanosoma cruzi Infection: Relation to Survival. Exp. Parasitol. 1995, 80, 499–506. [Google Scholar] [CrossRef]
- Zhong, C.; Qu, X.; Tan, M.; Meng, Y.G.; Ferrara, N. Characterization and Regulation of Bv8 in Human Blood Cells. Clin. Cancer Res. 2009, 15, 2675–2684. [Google Scholar] [CrossRef] [PubMed]
- Khusal, K.G.; Tonelli, R.R.; Mattos, E.C.; Soares, C.O.; Di Genova, B.M.; Juliano, M.A.; Urias, U.; Colli, W.; Alves, M.J.M. Prokineticin Receptor Identified by Phage Display Is an Entry Receptor for Trypanosoma cruzi into Mammalian Cells. Parasitol. Res. 2015, 114, 155–165. [Google Scholar] [CrossRef]
- Lattanzi, R.; Miele, R. Prokineticin-Receptor Network: Mechanisms of Regulation. Life 2022, 12, 172. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.S.; Cueto, J.A.; Casassa, A.F.; Vanrell, M.C.; Gottlieb, R.A.; Colombo, M.I. Molecular and Cellular Mechanisms Involved in the Trypanosoma cruzi/Host Cell Interplay. IUBMB Life 2012, 64, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Levy, O. Innate Immunity of the Newborn: Basic Mechanisms and Clinical Correlates. Nat. Rev. Immunol. 2007, 7, 379–390. [Google Scholar] [CrossRef]
- Espinoza, J.; Espinoza, A.F.; Power, G.G. High Fetal Plasma Adenosine Concentration: A Role for the Fetus in Preeclampsia? Am. J. Obstet. Gynecol. 2011, 205, 485.e24–485.e27. [Google Scholar] [CrossRef]
- Haskó, G.; Cronstein, B. Regulation of Inflammation by Adenosine. Front. Immunol. 2013, 4, 85. [Google Scholar] [CrossRef]
- Antonioli, L.; Csóka, B.; Fornai, M.; Colucci, R.; Kókai, E.; Blandizzi, C.; Haskó, G. Adenosine and Inflammation: What’s New on the Horizon? Drug Discov. Today 2014, 19, 1051–1068. [Google Scholar] [CrossRef]
- Zhong, L.; Peng, Q.; Zeng, X. The Role of Adenosine A1 Receptor on Immune Cells. Inflamm. Res. 2022, 71, 1203–1212. [Google Scholar] [CrossRef]
- Elahi, S.; Ertelt, J.M.; Kinder, J.M.; Jiang, T.T.; Zhang, X.; Xin, L.; Chaturvedi, V.; Strong, B.S.; Qualls, J.E.; Steinbrecher, K.A.; et al. Immunosuppressive CD71+ Erythroid Cells Compromise Neonatal Host Defence against Infection. Nature 2013, 504, 158–162. [Google Scholar] [CrossRef]
- Brook, B.; Harbeson, D.; Ben-Othman, R.; Viemann, D.; Kollmann, T.R. Newborn Susceptibility to Infection vs. Disease Depends on Complex in Vivo Interactions of Host and Pathogen. Semin. Immunopathol. 2017, 39, 615–625. [Google Scholar] [CrossRef]
- Sánchez-Valdéz, F.J.; Padilla, A.; Wang, W.; Orr, D.; Tarleton, R.L. Spontaneous Dormancy Protects Trypanosoma cruzi during Extended Drug Exposure. Elife 2018, 7, e34039. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lu, X.; Huang, J.; Jordan, P.; Ma, S.; Xu, L.; Hu, F.; Gui, H.; Zhao, H.; Bai, Z.; et al. Induction of Trained Immunity Protects Neonatal Mice Against Microbial Sepsis by Boosting Both the Inflammatory Response and Antimicrobial Activity. J. Inflamm. Res. 2022, 15, 3829–3845. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Hoffmann, F.W.; Fay, J.D.; Hashimoto, A.C.; Chapagain, M.L.; Kaufusi, P.H.; Hoffmann, P.R. Stimulation of Unprimed Macrophages with Immune Complexes Triggers a Low Output of Nitric Oxide by Calcium-Dependent Neuronal Nitric-Oxide Synthase. J. Biol. Chem. 2012, 287, 4492–4502. [Google Scholar] [CrossRef]
- de Taeye, S.W.; Rispens, T.; Vidarsson, G. The Ligands for Human IgG and Their Effector Functions. Antibodies 2019, 8, 30. [Google Scholar] [CrossRef]
- Vanderven, H.A.; Kent, S.J. The Protective Potential of Fc-Mediated Antibody Functions against Influenza Virus and Other Viral Pathogens. Immunol. Cell Biol. 2020, 98, 253–263. [Google Scholar] [CrossRef]
- Semmes, E.C.; Miller, I.G.; Wimberly, C.E.; Phan, C.T.; Jenks, J.A.; Harnois, M.J.; Berendam, S.J.; Webster, H.; Hurst, J.H.; Kurtzberg, J.; et al. Maternal Fc-Mediated Non-Neutralizing Antibody Responses Correlate with Protection against Congenital Human Cytomegalovirus Infection. J. Clin. Investig. 2022, 132, e156827. [Google Scholar] [CrossRef]
- Reyes, L.; Golos, T.G. Hofbauer Cells: Their Role in Healthy and Complicated Pregnancy. Front. Immunol. 2018, 9, 2628. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Chelariu-Raicu, A.; Makri, D.; Ori, C.; Ribeiro, P.C.P.; Kolben, T.; Kuhn, C.; Kessler, M.; Schulz, C.; Mahner, S.; et al. Updates of Placental Macrophages: Origins, Molecular Markers, Functions, and Related Diseases. J. Reprod. Immunol. 2023, 157, 103942. [Google Scholar] [CrossRef]
- Thomas, J.R.; Naidu, P.; Appios, A.; McGovern, N. The Ontogeny and Function of Placental Macrophages. Front. Immunol. 2021, 12, 771054. [Google Scholar] [CrossRef]
- Arruda, L.V.; Salomão, N.G.; Alves, F.d.A.V.; Rabelo, K. The Innate Defense in the Zika-Infected Placenta. Pathogens 2022, 11, 1410. [Google Scholar] [CrossRef] [PubMed]
- Doster, R.S.; Sutton, J.A.; Rogers, L.M.; Aronoff, D.M.; Gaddy, J.A. Streptococcus Agalactiae Induces Placental Macrophages To Release Extracellular Traps Loaded with Tissue Remodeling Enzymes via an Oxidative Burst-Dependent Mechanism. mBio 2018, 9, e02084-18. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, P.; Tang, Z.; Silasi, M.; Racicot, K.E.; Mor, G.; Abrahams, V.M.; Guller, S. Herpesvirus-Infected Hofbauer Cells Activate Endothelial Cells through an IL-1β-Dependent Mechanism. Placenta 2020, 91, 59–65. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M-B- | M+B- | ||
|---|---|---|---|
| CD54 | % (a) | 67.1 ± 6.2 (10) | 56.1 ± 8.0 (16) |
| MFI (b) | 14.1 ± 1.1 | 12.2 ± 2.2 | |
| HLA-DR | % (a) | 98.0 ± 0.3 (10) | 90.6 ± 4.9 (16) |
| MFI (b) | 95.9 ± 6.8 | 86.2 ± 12.4 | |
| TLR2 | % (a) | 98.1 ± 0.5 (9) | 96.2 ± 1.5 (11) |
| MFI (b) | 58.8 ± 0.5 | 52.3 ± 4.4 | |
| TLR4 | % (a) | 76.0 ± 4.8 (9) | 75.6 ± 5.1 (11) |
| MFI (b) | 30.8 ± 1.5 | 28.6 ± 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores, A.; Alonso-Vega, C.; Hermann, E.; Torrico, M.-C.; Montaño Villarroel, N.A.; Torrico, F.; Carlier, Y.; Truyens, C. Monocytes from Uninfected Neonates Born to Trypanosoma cruzi-Infected Mothers Display Upregulated Capacity to Produce TNF-α and to Control Infection in Association with Maternally Transferred Antibodies. Pathogens 2023, 12, 1103. https://doi.org/10.3390/pathogens12091103
Flores A, Alonso-Vega C, Hermann E, Torrico M-C, Montaño Villarroel NA, Torrico F, Carlier Y, Truyens C. Monocytes from Uninfected Neonates Born to Trypanosoma cruzi-Infected Mothers Display Upregulated Capacity to Produce TNF-α and to Control Infection in Association with Maternally Transferred Antibodies. Pathogens. 2023; 12(9):1103. https://doi.org/10.3390/pathogens12091103
Chicago/Turabian StyleFlores, Amilcar, Cristina Alonso-Vega, Emmanuel Hermann, Mary-Cruz Torrico, Nair Alaide Montaño Villarroel, Faustino Torrico, Yves Carlier, and Carine Truyens. 2023. "Monocytes from Uninfected Neonates Born to Trypanosoma cruzi-Infected Mothers Display Upregulated Capacity to Produce TNF-α and to Control Infection in Association with Maternally Transferred Antibodies" Pathogens 12, no. 9: 1103. https://doi.org/10.3390/pathogens12091103