
Effects of Host Plants and Their Infection Status on Acquisition and Inoculation of A Plant Virus by Its Hemipteran Vector

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Virus Maintenance
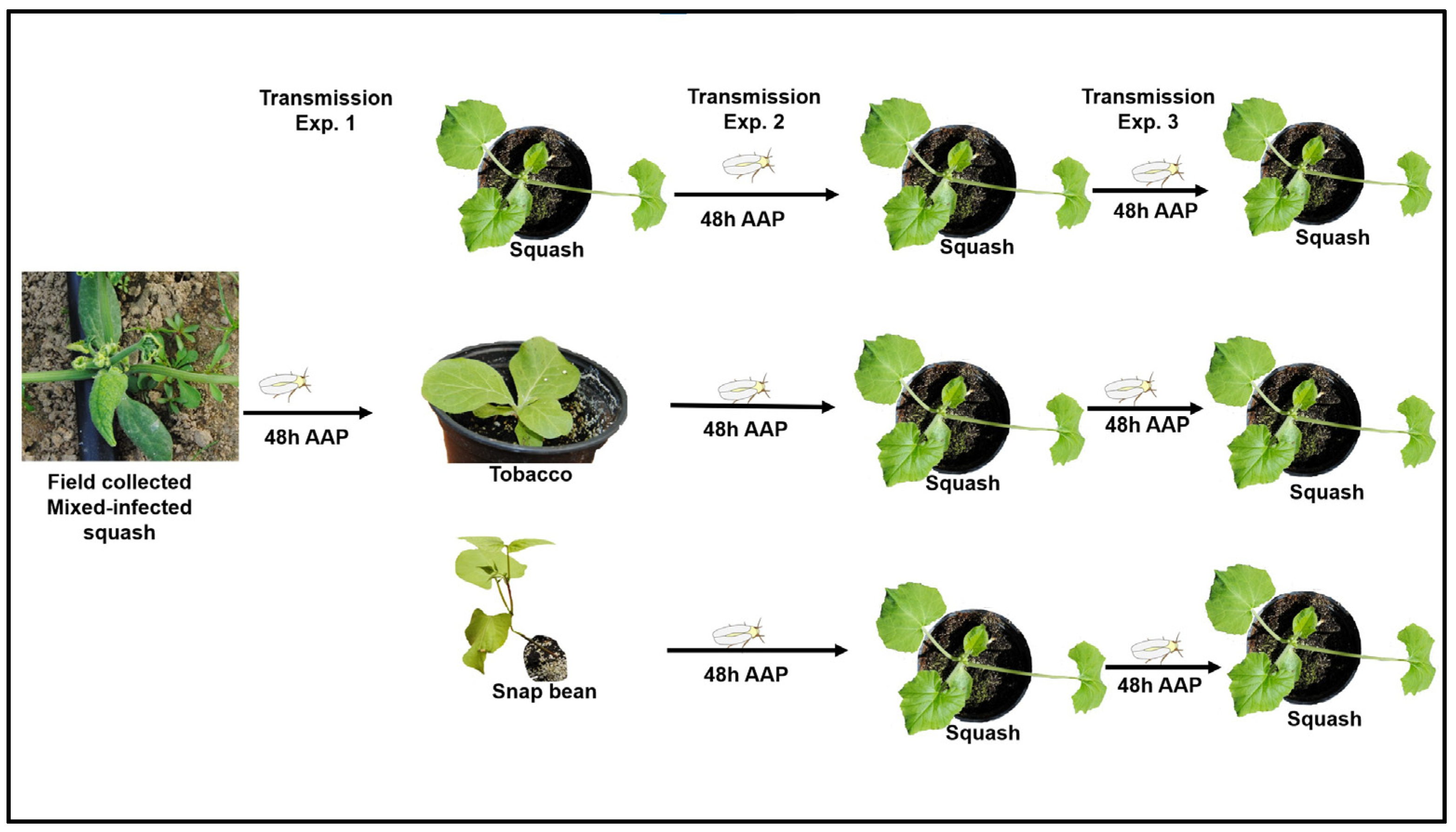
2.3. Transmission Experiment One (E1)
2.4. Transmission Experiment Two (E2)
2.5. Transmission Experiment Three (E3)
2.6. Statistical Analysis
3. Results
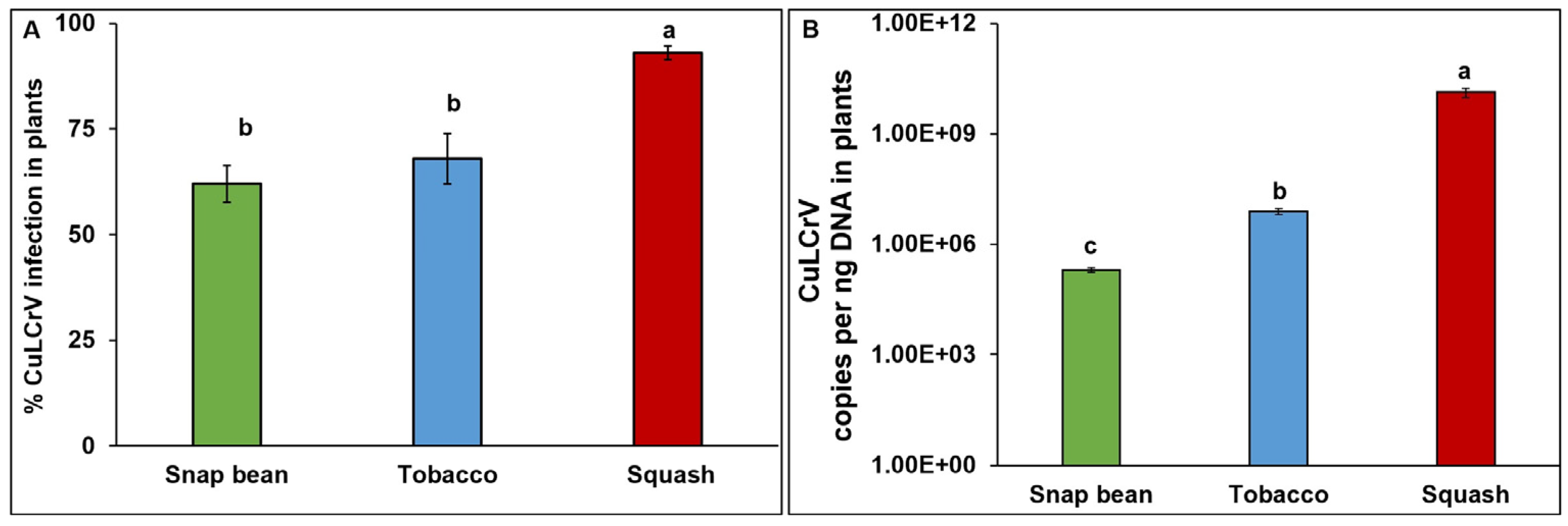
3.1. Transmission Experiment One (E1)
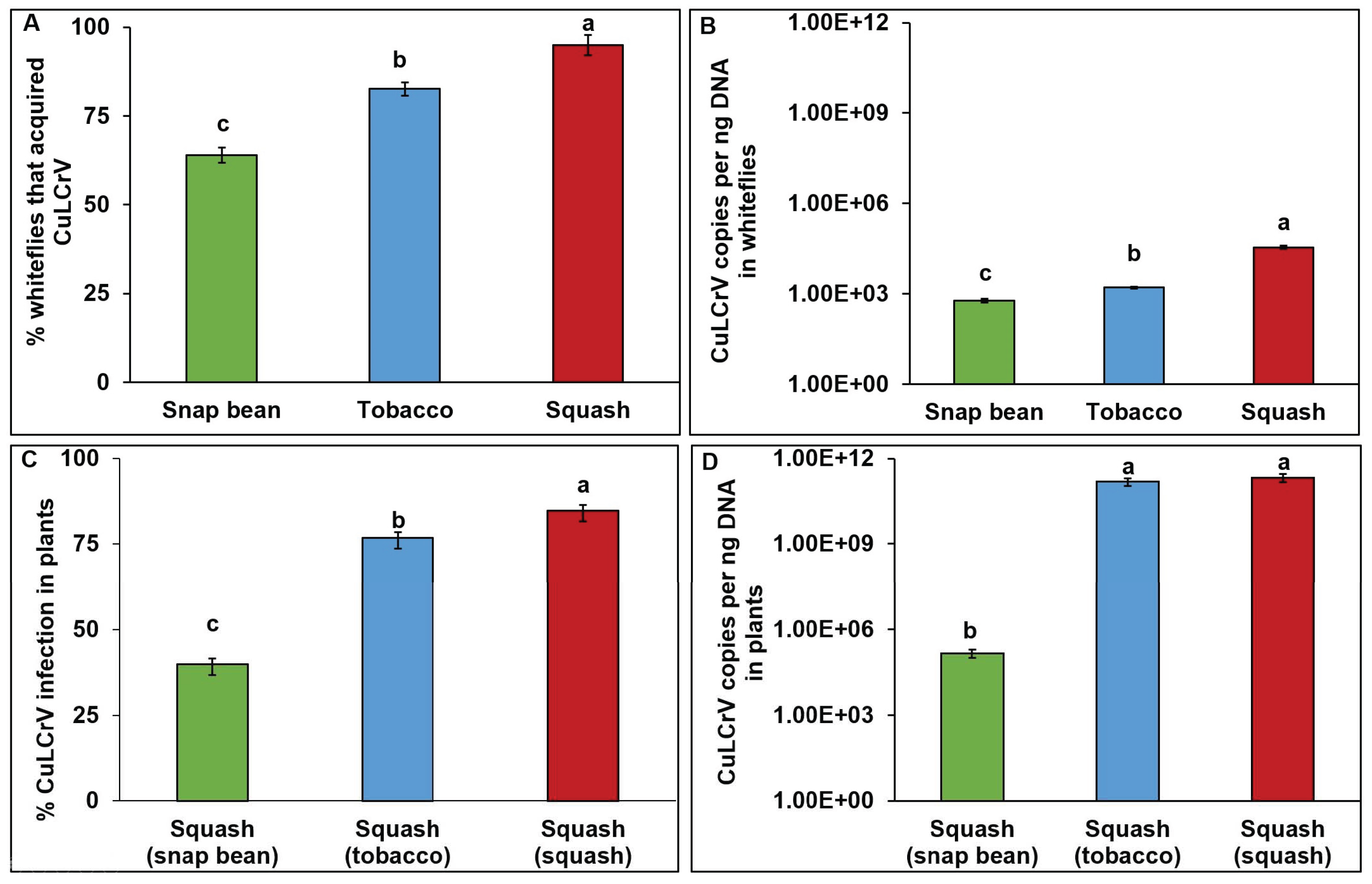
3.2. Transmission Experiment Two (E2)
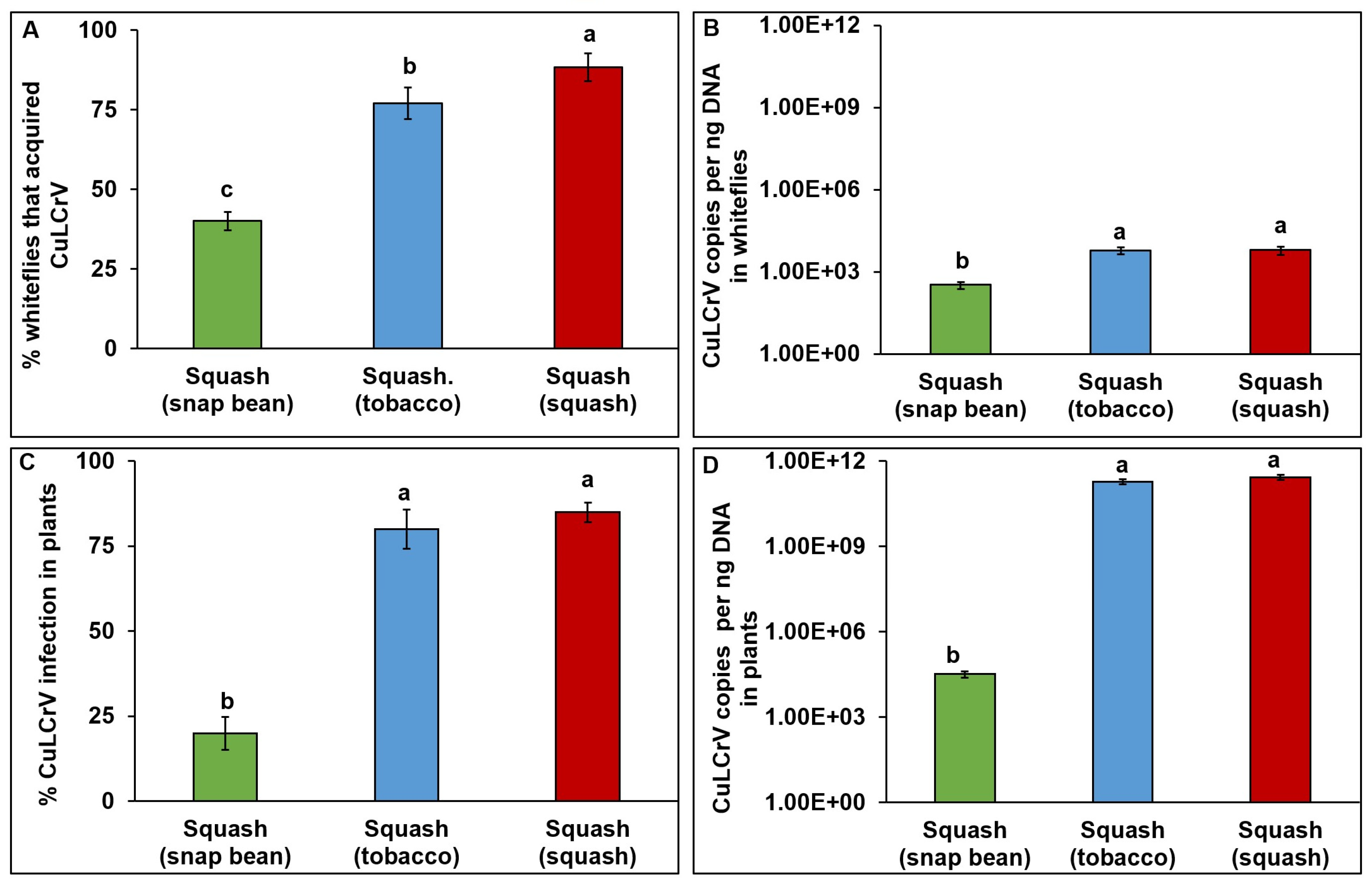
3.3. Transmission Experiment Three (E3)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunniffe, N.J.; Gilligan, C.A. Invasion, Persistence and Control in Epidemic Models for Plant Pathogens: The Effect of Host Demography. J. R. Soc. Interface 2010, 7, 439–451. [Google Scholar] [CrossRef] [PubMed]
- McLeish, M.J.; Fraile, A.; García-Arenal, F. Evolution of Plant–Virus Interactions: Host Range and Virus Emergence. Curr. Opin. Virol. 2019, 34, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect Vector-Mediated Transmission of Plant Viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef]
- Gautam, S.; Buck, J.W.; Dutta, B.; Coolong, T.; Sanchez, T.; Smith, H.A.; Adkins, S.; Srinivasan, R. Sida Golden Mosaic Virus, an Emerging Pathogen of Snap Bean (Phaseolus vulgaris L.) in the Southeastern United States. Viruses 2023, 15, 357. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Montero, J.R.; Ibarra, M.A.; Arias-Zambrano, M.; Peralta, E.L.; Brown, J.K. Phylo-biogeographical Distribution of Whitefly Bemisia tabaci (Insecta: Aleyrodidae) Mitotypes in Ecuador. Ecosphere 2020, 11, e03154. [Google Scholar] [CrossRef]
- de Moya, R.S.; Brown, J.K.; Sweet, A.D.; Walden, K.K.O.; Paredes-Montero, J.R.; Waterhouse, R.M.; Johnson, K.P. Nuclear Orthologs Derived from Whole Genome Sequencing Indicate Cryptic Diversity in the Bemisia Tabaci (Insecta: Aleyrodidae) Complex of Whiteflies. Diversity 2019, 11, 151. [Google Scholar] [CrossRef]
- Adkins, S.; Webster, C.G.; Kousik, C.S.; Webb, S.E.; Roberts, P.D.; Stansly, P.A.; Turechek, W.W. Ecology and Management of Whitefly-Transmitted Viruses of Vegetable Crops in Florida. Virus Res. 2011, 159, 110–114. [Google Scholar] [CrossRef]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-Virus Interactions in a Plant Host and in a Hemipteran Vector: Implications for Vector Fitness and Virus Epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef]
- LaTora, A.G.; Codod, C.B.; Legarrea, S.; Dutta, B.; Kemerait, R.C.; Adkins, S.; Turechek, W.; Coolong, T.; da Silva, A.L.B.R.; Srinivasan, R. Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia Tabaci MEAM1, and Viruses in Yellow Squash. Horticulturae 2022, 8, 341. [Google Scholar] [CrossRef]
- Agarwal, G.; Kavalappara, S.R.; Gautam, S.; Silva, A.D.; Simmons, A.; Srinivasan, R.; Dutta, B. Field Screen and Genotyping of Phaseolus Vulgaris against Two Begomoviruses in Georgia, USA. Insects 2021, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Akad, F.; Webb, S.; Nyoike, T.W.; Liburd, O.E.; Turechek, W.; Adkins, S.; Polston, J.E. Detection of Cucurbit Leaf Crumple Virus in Florida Cucurbits. Plant Dis. 2008, 92, 648. [Google Scholar] [CrossRef] [PubMed]
- Larsen, R.; Kmiecik, K. First Report of Cucurbit Leaf Crumple Virus in Snap Bean in Georgia. Phytopathology 2010, 100, S68. [Google Scholar]
- Keinath, A.P.; Ling, K.-S.; Adkins, S.; Hasegawa, D.K.; Simmons, A.M.; Hoak, S.; Mellinger, H.C.; Kousik, C.S. First Report of Cucurbit Leaf Crumple Virus Infecting Three Cucurbit Crops in South Carolina. Plant Health Prog. 2018, 19, 322–323. [Google Scholar] [CrossRef]
- Mascia, T.; Gallitelli, D. Synergies and Antagonisms in Virus Interactions. Plant Sci. 2016, 252, 176–192. [Google Scholar] [CrossRef]
- DaPalma, T.; Doonan, B.P.; Trager, N.M.; Kasman, L.M. A Systematic Approach to Virus–Virus Interactions. Virus Res. 2010, 149, 1–9. [Google Scholar] [CrossRef]
- Roossinck, M.J.; García-Arenal, F. Ecosystem Simplification, Biodiversity Loss and Plant Virus Emergence. Curr. Opin. Virol. 2015, 10, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Syller, J. Facilitative and Antagonistic Interactions between Plant Viruses in Mixed Infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef]
- Rentería-Canett, I.; Xoconostle-Cázares, B.; Ruiz-Medrano, R.; Rivera-Bustamante, R.F. Geminivirus Mixed Infection on Pepper Plants: Synergistic Interaction between PHYVV and PepGMV. Virol. J. 2011, 8, 104. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Cortez, A.A.; Anchieta, A.G.; Gulati-Sakhuja, A.; Hladky, L.L. Co-Infection by Two Criniviruses Alters Accumulation of Each Virus in a Host-Specific Manner and Influences Efficiency of Virus Transmission. Phytopathology 2008, 98, 1340–1345. [Google Scholar] [CrossRef]
- Gil-Salas, F.M.; Peters, J.; Boonham, N.; Cuadrado, I.M.; Janssen, D. Co-Infection with Cucumber Vein Yellowing Virus and Cucurbit Yellow Stunting Disorder Virus Leading to Synergism in Cucumber. Plant Pathol. 2012, 61, 468–478. [Google Scholar] [CrossRef]
- Srinivasan, R.; Alvarez, J.M. Effect of Mixed Viral Infections (Potato Virus Y-Potato Leafroll Virus) on Biology and Preference of Vectors Myzus Persicae and Macrosiphum Euphorbiae (Hemiptera: Aphididae). J. Econ. Entomol. 2007, 100, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Liao, Q.; Feng, J.; Li, D.; Chen, J. Synergy between Cucumber Mosaic Virus and Zucchini Yellow Mosaic Virus on Cucurbitaceae Hosts Tested by Real-Time Reverse Transcription-Polymerase Chain Reaction. Acta Biochim. Biophys. Sin. 2007, 39, 431–437. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, A.A.; Hanley-Bowdoin, L.; Kennedy, G.G.; Jacobson, A.L. Vector Acquisition and Co-Inoculation of Two Plant Viruses Influences Transmission, Infection, and Replication in New Hosts. Sci. Rep. 2022, 12, 20355. [Google Scholar] [CrossRef]
- Dietzgen, R.; Mann, K.; Johnson, K. Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef]
- Martinière, A.; Bak, A.; Macia, J.-L.; Lautredou, N.; Gargani, D.; Doumayrou, J.; Garzo, E.; Moreno, A.; Fereres, A.; Blanc, S.; et al. A Virus Responds Instantly to the Presence of the Vector on the Host and Forms Transmission Morphs. eLife 2013, 2, e00183. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Michalakis, Y.; Munster, M.; Blanc, S. Plant Feeding by Insect Vectors Can Affect Life Cycle, Population Genetics and Evolution of Plant Viruses. Funct. Ecol. 2013, 27, 610–622. [Google Scholar] [CrossRef]
- Moreno, A.B.; López-Moya, J.J. When Viruses Play Team Sports: Mixed Infections in Plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Milner, H.; Sparks, A.; McGregor, C.; Wintermantel, W.M.; Bag, S. First Report of Cucurbit Chlorotic Yellows Virus in Association with Other Whitefly-Transmitted Viruses in Yellow Squash (Cucurbita pepo) in Georgia, U.S.A. Plant Dis. 2021, 105, 1862. [Google Scholar] [CrossRef]
- Czosnek, H.; Ghanim, M.; Ghanim, M. The Circulative Pathway of Begomoviruses in the Whitefly Vector Bemisia Tabaci—Insights from Studies with Tomato Yellow Leaf Curl Virus. Ann. Appl. Biol. 2002, 140, 215–231. [Google Scholar] [CrossRef]
- Wisler, G.C.; Duffus, J.E.; Liu, H.-Y.; Li, R.H. Ecology and Epidemiology of Whitefly-Transmitted Closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Celix, A.; Lopez-Sese, A.; Almarza, N.; Gomez-Guillamon, M.L.; Rodriguez-Cerezo, E. Characterization of Cucurbit Yellow Stunting Disorder Virus, a Bemisia Tabaci-Transmitted Closterovirus. Phytopathology 1996, 8, 1370–1376. [Google Scholar]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Sparks, A.N.; Adkins, S.; Srinivasan, R. First Report of Cucurbit Yellow Stunting Disorder Virus in Cucurbits in Georgia, United States. Plant Health Prog. 2018, 19, 9–10. [Google Scholar] [CrossRef]
- Hagen, C.; Rojas, M.R.; Sudarshana, M.R.; Xoconostle-Cazares, B.; Natwick, E.T.; Turini, T.A.; Gilbertson, R.L. Biology and Molecular Characterization of Cucurbit Leaf Crumple Virus, an Emergent Cucurbit-Infecting Begomovirus in the Imperial Valley of California. Plant Dis. 2008, 92, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Wintermantel, W.M.; Hladky, L.L.; Cortez, A.A.; Natwick, E.T. A New Expanded Host Range of Cucurbit Yellow Stunting Disorder Virus Includes Three Agricultural Crops. Plant Dis. 2009, 93, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, N.A.; Sudarshana, M.R.; Guzman, P.; Gilbertson, R.L. Generation and Characterization of Infectious Clones of Cucurbit Leaf Crumple Virus, a New Bipartite Geminivirus from the Imperial Valley of California. Phytopathology 2001, 91. Available online: https://apsjournals.apsnet.org/doi/pdf/10.1094/PHYTO.2000.90.6.S1 (accessed on 20 July 2023).
- Brown, J.K.; Idris, A.M.; Alteri, C.; Stenger, D.C. Emergence of a New Cucurbit-Infecting Begomovirus Species Capable of Forming Viable Reassortants with Related Viruses in the Squash Leaf Curl Virus Cluster. Phytopathology 2002, 92, 734–742. [Google Scholar] [CrossRef]
- The University of Georgia. Georgia Farm Gate Value Report 2021; University of Georgia College of Agricultural and Environmental Sciences Center for Agribusiness and Economic Development: Athens, GA, USA, 2022; p. 176, AR-22-01; Available online: https://caed.uga.edu/content/dam/caes-subsite/caed/publications/annual-reports-farm-gate-value-reports/2021_GeorgiaFGVReportDec2022%20(1).pdf (accessed on 20 July 2023).
- Kuo, Y.-W.; Rojas, M.R.; Gilbertson, R.L.; Wintermantel, W.M. First First Report of Cucurbit Yellow Stunting Disorder Virus in California and Arizona, in Association with Cucurbit Leaf Crumple Virus and Squash Leaf Curl Virus. Plant Dis. 2007, 91, 330. [Google Scholar] [CrossRef]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal Effects of a Begomovirus Infection and Host Plant Resistance on the Preference and Development of an Insect Vector, Bemisia Tabaci, and Implications for Epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar] [CrossRef]
- Frohlich, D.R.; Torres-Jerez, I.; Bedford, I.D.; Markham, P.G.; Brown, J.K. A Phylogeographical Analysis of the Bemisia Tabaci Species Complex Based on Mitochondrial DNA Markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Cooper, I.; Jones, R.A.C. Wild Plants and Viruses: Under-Investigated Ecosystems. Adv. Virus Res. 2006, 67, 1–47. [Google Scholar] [PubMed]
- Liu, B.; Yan, F.; Chu, D.; Pan, H.; Jiao, X.; Xie, W.; Wu, Q.; Wang, S.; Xu, B.; Zhou, X.; et al. Difference in Feeding Behaviors of Two Invasive Whiteflies on Host Plants with Different Suitability: Implication for Competitive Displacement. Int. J. Biol. Sci. 2012, 8, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Shi, X.; Liu, B.; Pan, H.; Wei, W.; Zeng, Y.; Sun, X.; Xie, W.; Wang, S.; Wu, Q.; et al. Transmission of Tomato Yellow Leaf Curl Virus by Bemisia Tabaci as Affected by Whitefly Sex and Biotype. Sci. Rep. 2015, 5, 10744. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M.; Pilowsky, M.; Ben-Joseph, R.; Cohen, S. Effect of Host Plant Resistance to Tomato Yellow Leaf Curl Virus (TYLCV) on Virus Acquisition and Transmission by Its Whitefly Vector. Phytopathology 2001, 91, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Kollenberg, M.; Winter, S.; Götz, M. Quantification and Localization of Watermelon Chlorotic Stunt Virus and Tomato Yellow Leaf Curl Virus (Geminiviridae) in Populations of Bemisia Tabaci (Hemiptera, Aleyrodidae) with Differential Virus Transmission Characteristics. PLoS ONE 2014, 9, e111968. [Google Scholar] [CrossRef]
- Hagen, C.; Rojas, M.R.; Kon, T.; Gilbertson, R.L. Recovery from Cucurbit Leaf Crumple Virus (Family Geminiviridae, Genus Begomovirus) Infection Is an Adaptive Antiviral Response Associated with Changes in Viral Small RNAs. Phytopathology 2008, 98, 1029–1037. [Google Scholar] [CrossRef]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA Silencing against Geminivirus: Complementary Action of Posttranscriptional Gene Silencing and Transcriptional Gene Silencing in Host Recovery. J. Virol. 2009, 83, 1332–1340. [Google Scholar] [CrossRef]
- Raja, P.; Wolf, J.N.; Bisaro, D.M. RNA Silencing Directed against Geminiviruses: Post-Transcriptional and Epigenetic Components. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2010, 1799, 337–351. [Google Scholar] [CrossRef]
- Pooggin, M.; Shivaprasad, P.V.; Veluthambi, K.; Hohn, T. RNAi Targeting of DNA Virus in Plants. Nat. Biotechnol. 2003, 21, 131–132. [Google Scholar] [CrossRef]
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, C.M. Effect of Temperature on Geminivirus-Induced RNA Silencing in Plants. Plant Physiol. 2005, 138, 1828–1841. [Google Scholar] [CrossRef]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. Short Interfering RNA Accumulation Correlates with Host Recovery in DNA Virus-Infected Hosts, and Gene Silencing Targets Specific Viral Sequences. J. Virol. 2004, 78, 7465–7477. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, J.; Lozoya-Gloria, E.; Rivera-Bustamante, R.F. Symptom Remission and Specific Resistance of Pepper Plants after Infection by Pepper Golden Mosaic Virus. Phytopathology 2007, 97, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Moissiard, G.; Voinnet, O. Viral Suppression of RNA Silencing in Plants. Mol. Plant Pathol. 2004, 5, 71–82. [Google Scholar] [CrossRef]
- Bisaro, D.M. Silencing Suppression by Geminivirus Proteins. Virology 2006, 344, 158–168. [Google Scholar] [CrossRef]
- Moury, B.; Fabre, F.; Senoussi, R. Estimation of the Number of Virus Particles Transmitted by an Insect Vector. Proc. Natl. Acad. Sci. USA 2007, 104, 17891–17896. [Google Scholar] [CrossRef] [PubMed]
- Pirone, T.P. Quantity of Virus Required for Aphid Transmission of a Potyvirus. Phytopathology 1988, 78, 104. [Google Scholar] [CrossRef]
- Murphy, J.F.; Bowen, K.L. Synergistic Disease in Pepper Caused by the Mixed Infection of Cucumber Mosaic Virus and Pepper Mottle Virus. Phytopathology 2006, 96, 240–247. [Google Scholar] [CrossRef] [PubMed]
- bance, V.B. Replication of Potato Virus X RNA Is Altered in Coinfections with Potato Virus Y. Virology 1991, 182, 486–494. [Google Scholar] [CrossRef]
- Mauck, K.; Bosque-Pérez, N.A.; Eigenbrode, S.D.; De Moraes, C.M.; Mescher, M.C. Transmission Mechanisms Shape Pathogen Effects on Host-Vector Interactions: Evidence from Plant Viruses. Funct. Ecol. 2012, 26, 1162–1175. [Google Scholar] [CrossRef]
- Lefèvre, T.; Thomas, F. Behind the Scene, Something Else Is Pulling the Strings: Emphasizing Parasitic Manipulation in Vector-Borne Diseases. Infect. Genet. Evol. 2008, 8, 504–519. [Google Scholar] [CrossRef]
- Guzman, P.; Sudarshana, M.R.; Seo, Y.-S.; Rojas, M.R.; Natwick, E.; Turini, T.; Mayberry, K.; Gilbertson, R.L. A New Bipartite Geminivirus (Begomovirus) Causing Leaf Curl and Crumpling in Cucurbits in the Imperial Valley of California. Plant Dis. 2000, 84, 488. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J. Symbiosis versus Competition in Plant Virus Evolution. Nat. Rev. Microbiol. 2005, 3, 917–924. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Simmons, A.M.; Srinivasan, R. Effects of Host Plants and Their Infection Status on Acquisition and Inoculation of A Plant Virus by Its Hemipteran Vector. Pathogens 2023, 12, 1119. https://doi.org/10.3390/pathogens12091119
Gautam S, Gadhave KR, Buck JW, Dutta B, Coolong T, Adkins S, Simmons AM, Srinivasan R. Effects of Host Plants and Their Infection Status on Acquisition and Inoculation of A Plant Virus by Its Hemipteran Vector. Pathogens. 2023; 12(9):1119. https://doi.org/10.3390/pathogens12091119
Chicago/Turabian StyleGautam, Saurabh, Kiran R. Gadhave, James W. Buck, Bhabesh Dutta, Timothy Coolong, Scott Adkins, Alvin M. Simmons, and Rajagopalbabu Srinivasan. 2023. "Effects of Host Plants and Their Infection Status on Acquisition and Inoculation of A Plant Virus by Its Hemipteran Vector" Pathogens 12, no. 9: 1119. https://doi.org/10.3390/pathogens12091119