

The Emergence of Carbapenem- and Colistin-Resistant Enterobacteria in Senegal
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Isolates
2.2. Bacteriological Analyses
2.3. Molecular Analysis
3. Results
3.1. Distribution of Samples and Isolates
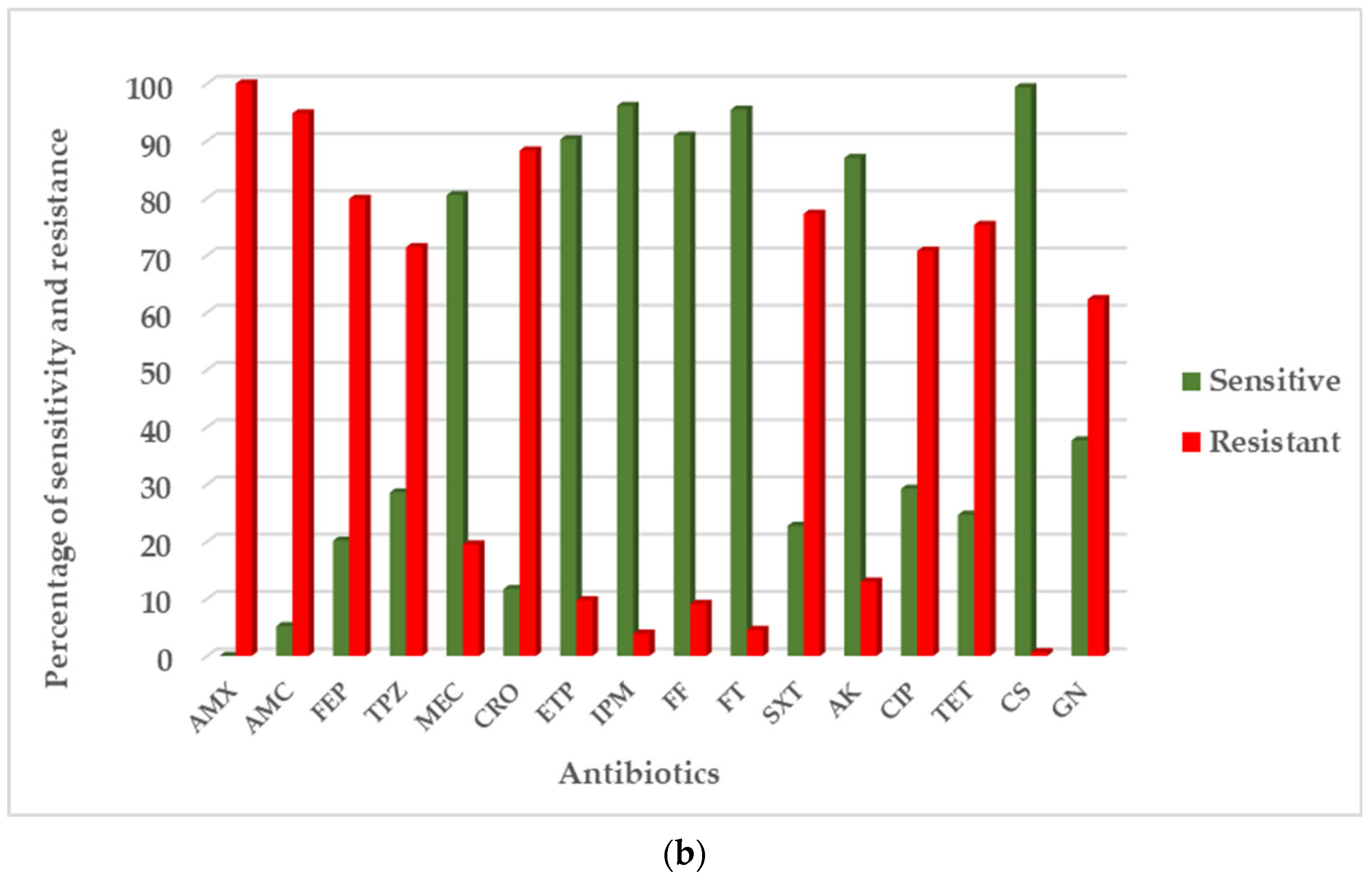
3.2. Antibiotic Resistance Phenotype
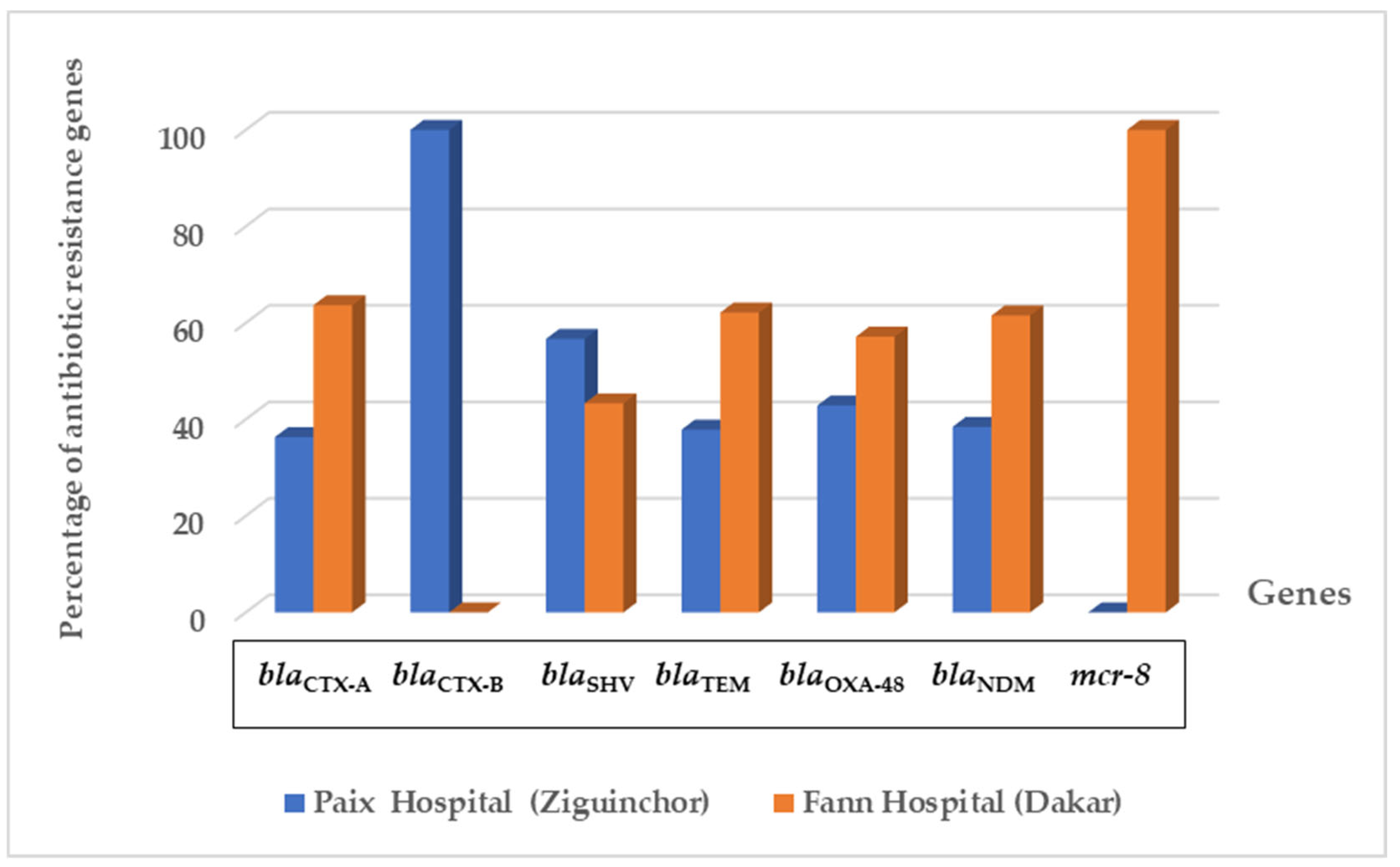
3.3. Distribution of Class A Beta-Lactamase Genes
3.4. Carbapenem Resistance Genes
3.5. Plasmid Colistin Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 521–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Cassini, A.; Plachouras, D.; Eckmanns, T.; Abu Sin, M.; Blank, H.-P.; Ducomble, T.; Haller, S.; Harder, T.; Klingeberg, A.; Sixtensson, M.; et al. Burden of six healthcare-associated infections on European population health: Estimating incidence-based disability-adjusted life years through a population prevalence-based modelling study. PLoS Med. 2016, 13, e1002150. [Google Scholar] [CrossRef] [Green Version]
- Suetens, C.; Latour, K.; Kärki, T.; Ricchizzi, E.; Kinross, P.; Moro, M.L.; Jans, B.; Hopkins, S.; Hansen, S.; Lyytikäinen, O.; et al. Prevalence of healthcare-associated infections, estimated incidence and composite antimicrobial resistance index in acute care hospitals and long-term care facilities: Results from two European point prevalence surveys, 2016 to 2017. Eurosurveillance 2018, 23, 1800516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouz, N.; Pangesti, K.N.; Yasir, M.; Al-Malki, A.L.; Azhar, E.I.; Hill-Cawthorne, G.A.; El Ghany, M.A. The contribution of wastewater to the transmission of antimicrobial resistance in the environment: Implications of mass gathering settings. Trop. Med. Infect. Dis. 2020, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Alsan, M.; Schoemaker, L.; Eggleston, K.; Kammili, N.; Kolli, P.; Bhattacharya, J. Out-of-pocket health expenditures and antimicrobial resistance in low-income and middle-income countries: An economic analysis. Lancet Infect. Dis. 2015, 15, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Okeke, I.N.; Aboderin, O.A.; Byarugaba, D.K.; Ojo, K.K.; Opintan, J.A. Growing problem of multidrug-resistant enteric pathogens in Africa. Emerg. Infect. Dis. 2007, 13, 1640–1646. [Google Scholar] [CrossRef]
- Byarugaba, D.K. Antimicrobial resistance in developing countries and responsible risk factors. Int. J. Antimicrob. Agents 2004, 24, 105–110. [Google Scholar] [CrossRef]
- Fortes Déguénonvo, L.; Diallo, M.O.S.; Dia Badiane, N.M.; Lakhe, N.A.; Diop, S.A.; Manga, N.M.; Seydi, M. Evaluation de la qualité de la prescription des antibiotiques au Service d’Accueil des Urgences du CHNU de Fann à Dakar. Med. Mal. Infect. 2013, 43, 78. Available online: https://www.infectiologie.com/UserFiles/File/medias/JNI/JNI13/posters/2013-JNI-Q-03.pdf (accessed on 23 May 2023).
- Lakhe, N.A.; Sylla, K.; Mbaye, K.D.; Ndiaye, R.; Diallo, V.M.P.C.; Ka, D.; Dia, M.L.; Deguenonvo, L.F.; Ndour, C.T.; Seydi, M. Bacteremia: Profile and Antibiotic Resistance at the Infectious and Tropical Diseases Clinic in Fann Hospital, Dakar, Senegal. J. Infect. Dis. Ther. 2018, 6, 1. [Google Scholar] [CrossRef]
- Bernabé, K.J.; Langendorf, C.; Ford, N.; Ronat, J.-B.; Murphy, R.A. Antimicrobial resistance in West Africa: A systematic review and meta-analysis. Int. J. Antimicrob. Agents 2017, 50, 629–639. [Google Scholar] [CrossRef]
- Manenzhe, R.I.; Zar, H.J.; Nicol, M.P.; Kaba, M. The spread of carbapenemase-producing bacteria in Africa: A systematic review. J. Antimicrob. Chemother. 2015, 70, 23–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diene, S.M.; Fall, B.; Kempf, M.; Fenollar, F.; Sow, K.; Niang, B.; Wade, B.; Raoult, D.; Rolain, J.-M. Emergence of the OXA-23 carbapenemase-encoding gene in multidrug-resistant Acinetobacter baumannii clinical isolates from the Principal Hospital of Dakar, Senegal. Int. J. Infect. Dis. 2013, 17, e209–e210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petti, C.A.; Polage, C.R.; Quinn, T.C.; Ronald, A.R.; Sande, M.A. Laboratory Medicine in Africa: A Barrier to Effective Health Care. Clin. Infect. Dis. 2006, 42, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vounba, P.; Rhouma, M.; Arsenault, J.; Alambédji, R.B.; Fravalo, P.; Fairbrother, J.M. Prevalence of colistin resistance and mcr-1/mcr-2 genes in extended-spectrum β-lactamase/AmpC-producing Escherichia coli isolated from chickens in Canada, Senegal and Vietnam. J. Glob. Antimicrob. Resist. 2019, 19, 222–227. [Google Scholar] [CrossRef]
- Nation, R.L.; Li, J. Colistin in the 21st century. Curr. Opin. Infect. Dis. 2009, 22, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Moquet, O.; Bouchiat, C.; Kinana, A.; Seck, A.; Arouna, O.; Bercion, R.; Breurec, S.; Garin, B. Class D OXA-48 carbapenemase in multidrug-resistant enterobacteria, Senegal. Emerg. Infect. Dis. 2011, 17, 143. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Harbottle, H.; Thakur, S.; Zhao, S.; White, D.G. Genetics of antimicrobial resistance. Anim. Biotechnol. 2006, 17, 111–124. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Shen, Y.; Shen, J.; Wu, C. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect. Dis. 2016, 16, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPhee, J.B.; Lewenza, S.; Hancock, R.E. Cationic antimicrobial peptides activate a two-component regulatory system, PmrA-PmrB, that regulates resistance to polymyxin B and cationic antimicrobial peptides in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Panta, P.R.; Kumar, S.; Stafford, C.F.; Billiot, C.E.; Douglass, M.V.; Herrera, C.M.; Trent, M.S.; Doerrler, W.T. A DedA Family Membrane Protein Is Required for Burkholderia thailandensis Colistin Resistance. Front. Microbiol. 2019, 10, 2532. [Google Scholar] [CrossRef] [Green Version]
- Jana, B.; Cain, A.K.; Doerrler, W.T.; Boinett, C.J.; Fookes, M.C.; Parkhill, J.; Guardabassi, L. The secondary resistome of multidrug-resistant Klebsiella pneumoniae. Sci. Rep. 2017, 7, 42483. [Google Scholar] [CrossRef] [Green Version]
- Dia, M.L.; Ngom, B.; Diagne, R.; Ka, R.; Lo, S.; Cisse, M.F.; Arlet, G.; Sow, A.I. Molecular detection of CTX-M-15-type β-lactamases in Escherichia coli strains from Senegal. New Microbes New Infect. 2016, 9, 45–46. [Google Scholar] [CrossRef] [Green Version]
- Koudokpon, H.; Dougnon, V.; Hadjadj, L.; Kissira, I.; Fanou, B.; Loko, F.; Bankole, H.S.; Diene, S.; Rolain, J.-M. First sequence analysis of genes mediating extended-spectrum beta-lactamase (ESBL) bla-TEM, SHV-and CTX-M production in isolates of enterobacteriaceae in Southern Benin. Int. J. Infect. 2018, 5, e83194. [Google Scholar] [CrossRef] [Green Version]
- Dougnon, V.; Assogba, P.; Mohammed, J.; Agbankpe, J.; Deguenon, E.; Fabiyi, K.; Dougnon, J.; Baba-Moussa, L.; Bankole, H. Urinary Tract Infections in Benin: Exploring the Virulence Factors and Antibiotic Resistance and Virulence Genes among Bacterial Isolates. Int. J. Pathog. Res. 2021, 7, 28–36. [Google Scholar] [CrossRef]
- Carrër, A.; Poirel, L.; Eraksoy, H.; Cagatay, A.A.; Badur, S.; Nordmann, P. Spread of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in Istanbul, Turkey. Antimicrob. Agents Chemother. 2008, 52, 2950–2954. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P. Carbapenemase-producing Enterobacteriaceae: Overview of a major public health challenge. Med. Mal. Infect. 2014, 44, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, S.; Robin, F.; Beyrouthy, R.; Ba-Diallo, A.; Niang, A.A.; Diagne, R.; Diop, A.; Camara, M.; Ka, R.; Gaye-Diallo, A.; et al. OXA-48 type carbapenemase in Klebsiella pneumoniae producing extended spectrum B-lactamases (ESBL) in Senegal. Afr. J. Microbiol. Res. 2018, 12, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Camara, M.; Saad Bouh Boye, C.; Mboup, S.; Gaye-Diallo, A.; Toure-Kane, C.; Diop-Ndiaye, H.; Sow Ndoye, A.; Ngom-Cisse, S.; Diagne-Samb, H.; Karam, F. Molecular Epidemiology of Carbapenem-resistant Acinetobacter baumannii Isolates in a Senegalese University Teaching Hospital. J. Adv. Microbiol. 2022, 73–82. [Google Scholar] [CrossRef]
- Seng, P.; Drancourt, M.; Gouriet, F.; La Scola, B.; Fournier, P.-E.; Rolain, J.M.; Raoult, D. Ongoing revolution in bacteriology: Routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Infect. Dis. 2009, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 12.0. Sweden: European Committee on Antimicrobial Susceptibility Testing; [Jan; 2023]. 2022. European Committee on Antimicrobial Susceptibility Testing. EUCAST Clinical Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 12.0. Available online: https://www.eucast.org/ (accessed on 23 July 2023).
- Dashti, A.A.; Jadaon, M.M.; Abdulsamad, A.M.; Dashti, H.M. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- Roschanski, N.; Fischer, J.; Guerra, B.; Roesler, U. Development of a multiplex real-time PCR for the rapid detection of the predominant beta-lactamase genes CTX-M, SHV, TEM and CIT-type AmpCs in Enterobacteriaceae. PLoS ONE 2014, 9, e100956. [Google Scholar] [CrossRef]
- El-Sawalhi, S.; Revol, O.; Chamieh, A.; Lacoste, A.; Annessi, A.; La Scola, B.; Rolain, J.-M.; Pagnier, I. Epidemiological Description and Detection of Antimicrobial Resistance in Various Aquatic Sites in Marseille, France. Microbiol. Spectr. 2023, 11, e0142622. [Google Scholar] [CrossRef]
- Arabacı, Ç.; Dal, T.; Başyiğit, T.; Genişel, N.; Durmaz, R. Investigation of carbapenemase and mcr-1 genes in carbapenem-resistant Klebsiella pneumoniae isolates. J. Infect. Dev. Ctries. 2019, 13, 504–509. [Google Scholar] [CrossRef]
- Touati, M.; Hadjadj, L.; Berrazeg, M.; Baron, S.A.; Rolain, J.M. Emergence of Escherichia coli harbouring mcr-1 and mcr-3 genes in North West Algerian farmlands. J. Glob. Antimicrob. Resist. 2020, 21, 132–137. [Google Scholar] [CrossRef]
- Nabti, L.Z.; Sahli, F.; Ngaiganam, E.P.; Radji, N.; Mezaghcha, W.; Lupande-Mwenebitu, D.; Baron, S.A.; Rolain, J.-M.; Diene, S.M. Development of real-time PCR assay allowed describing the first clinical Klebsiella pneumoniae isolate harboring plasmid-mediated colistin resistance mcr-8 gene in Algeria. J. Glob. Antimicrob. Resist. 2020, 20, 266–271. [Google Scholar] [CrossRef]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [Green Version]
- Diakhaby, E.M.; Diop, H.; Ndiaye, A.J.S.; Macondo, E.A.; Camara, M.; Siby, T.; Boye, C.S.B.; Kane, C.T. Molecular aspect of multi-drug resistant extended spectrum betalactamases (esbl) secreting enterobacteriaceae strains isolated at both bacteriology laboratory of the teaching hospital aristide le dantec and at the private medical biology laboratory bio-24 of dakar in Senegal. Glob. J. Re-Search Anal. 2020, 9, 5. [Google Scholar] [CrossRef]
- Poirel, L.; Gniadkowski, M.; Nordmann, P. Biochemical analysis of the ceftazidime-hydrolysing extended-spectrum β-lactamase CTX-M-15 and of its structurally related β-lactamase CTX-M-3. J. Antimicrob. Chemother. 2002, 50, 1031–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, R.; Yimagou, E.K.; Hadjadj, L.; Ibrahim, A.; Davoust, B.; Barciela, A.; Hernandez-Aguilar, R.A.; Diatta, G.; Sokhna, C.; Raoult, D. Population diversity of antibiotic resistant Enterobacterales in samples from wildlife origin in Senegal: Identification of a multidrug resistance transposon carrying blaCTX-M-15 in E. coli. Front. Microbiol. 2022, 13, 838392. [Google Scholar] [CrossRef] [PubMed]
- Diene, S.M.; Fenollar, F.; Fall, B.; Sow, K.; Niang, B.; Samba Ba, P.; Wade, B.; Raoult, D.; Rolain, J.-M. CTX-M-15-producing M organella morganii from H ôpital P rincipal de Dakar, Senegal. New Microbes New Infect. 2014, 2, 46–49. [Google Scholar] [CrossRef]
- Ruppé, E. Épidémiologie des bêta-lactamases à spectre élargi: L’avènement des CTX-M. Antibiotiques 2010, 12, 3–16. [Google Scholar] [CrossRef]
- Lutgring, J.D.; Zhu, W.; de Man, T.J.; Avillan, J.J.; Anderson, K.F.; Lonsway, D.R.; Rowe, L.A.; Batra, D.; Rasheed, J.K.; Limbago, B.M. Phenotypic and genotypic characterization of Enterobacteriaceae producing oxacillinase-48–like carbapenemases, United States. Emerg. Infect. Dis. 2018, 24, 700. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Cantón, R.; Akóva, M.; Carmeli, Y.; Giske, C.G.; Glupczynski, Y.; Gniadkowski, M.; Livermore, D.M.; Miriagou, V.; Naas, T.; Rossolini, G.M.; et al. Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2012, 18, 413–431. [Google Scholar] [CrossRef] [Green Version]
- Cornaglia, G.; Giamarellou, H.; Rossolini, G.M. Metallo-β-lactamases: A last frontier for β-lactams? Lancet Infect. Dis. 2011, 11, 381–393. [Google Scholar] [CrossRef]
- Kieffer, N.; Poirel, L.; Nordmann, P.; Madec, J.-Y.; Haenni, M. Emergence of colistin resistance in Klebsiella pneumoniae from veterinary medicine. J. Antimicrob. Chemother. 2015, 70, 1265–1267. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Basu, S.; Rath, S.; Sahu, S.K. Colistin resistance in Gram-negative ocular infections: Prevalence, clinical outcome and antibiotic susceptibility patterns. Int. Ophthalmol. 2020, 40, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Ortwine, J.K.; Kaye, K.S.; Li, J.; Pogue, J.M. Colistin: Understanding and applying recent pharmacokinetic advances. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2015, 35, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.B.; Velkov, T.; Nation, R.L.; Forrest, A.; Tsuji, B.T.; Bergen, P.J.; Li, J. Pharmacokinetics/pharmacodynamics of colistin and polymyxin B: Are we there yet? Int. J. Antimicrob. Agents 2016, 48, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Guérin, F.; Isnard, C.; Sinel, C.; Morand, P.; Dhalluin, A.; Cattoir, V.; Giard, J.-C. Cluster-dependent colistin hetero-resistance in Enterobacter cloacae complex. J. Antimicrob. Chemother. 2016, 71, 3058–3061. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Bardet, L.; Dubourg, G.; Fichaux, M.; Rolain, J.-M. mcr-1 plasmid-mediated colistin resistance gene detection in an Enterobacter cloacae clinical isolate in France. J. Glob. Antimicrob. Resist. 2017, 10, 35–36. [Google Scholar] [CrossRef]
- Sartelli, M.; Hardcastle, T.C.; Catena, F.; Chichom-Mefire, A.; Coccolini, F.; Dhingra, S.; Haque, M.; Hodonou, A.; Iskandar, K.; Labricciosa, F.M. Antibiotic use in low and middle-income countries and the challenges of antimicrobial resistance in surgery. Antibiotics 2020, 9, 497. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Number of Isolates (%) | Total (%) | |
|---|---|---|---|
| Paix Hospital (Ziguinchor) | Fann Hospital (Dakar) | ||
| Escherichia coli | 35 (14.58%) | 111 (46.25) | 146 (60.83%) |
| Klebsiella pneumoniae | 29 (12.08%) | 23 (9.59%) | 52 (21.67%) |
| Enterobacter cloacae | 16 (6.65%) | 17 (7.08%) | 33 (13.75%) |
| Citrobacter freundii | 4 (1.66%) | 1 (0.42%) | 5 (2.08%) |
| Klebsiella aerogenes | 1 (0.42%) | 0 (0%) | 1 (0.42%) |
| Proteus mirabilis | 0 (0%) | 1 (0.42%) | 1 (0.42%) |
| Serratia marcesens | 1 (0.42%) | 1 (0.42%) | 2 (0.84%) |
| Total (%) | 86 (35.83%) | 154 (64.17%) | 240 (100%) |
| Species | Number (%) | Antibiotic Resistance Genes | ||||||
|---|---|---|---|---|---|---|---|---|
| blaCTX-A | blaCTX-B | blaSHV | blaTEM | blaOXA-48 | blaNDM | mcr-8 | ||
| Escherichia coli | 146 (60.83%) | 123 (84.2%) | 1 (0.6%) | 26 (17.8%) | 90 (61.6%) | 5 (3.4%) | 5 (3.4%) | 0 (0%) |
| Klebsiella pneumoniae | 52 (21.67%) | 50 (96.1%) | 0 (0%) | 52 (100%) | 46 (88.4%) | 3 (5.7%) | 4 (7.7%) | 0 (0%) |
| Enterobacter cloacae | 33 (13.75%) | 22 (66.6%) | 0 (0%) | 10 (30.3%) | 27 (81.8%) | 5 (15.1%) | 4 (12.1%) | 1 (3.0%) |
| Citrobacter freundii | 5 (2.08%) | 4 (80%) | 0 (0%) | 1 (20%) | 5 (100%) | 1 (20%) | 0 (0%) | 0 (0%) |
| Klebsiella aerogenes | 1 (0.42%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Proteus mirabilis | 1 (0.42%) | 1 (100%) | 0 (0%) | 0 (0%) | 1 (100%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Serratia marcescens | 2 (0.83%) | 1 (50%) | 0 (0%) | 1 (50%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Total (%) | 240 (100%) | 201 (83.75%) | 1 (0.42%) | 90 (37.5%) | 169 (70.41%) | 14 (5.83%) | 13 (5.41%) | 1 (0.42%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarr, H.; Niang, A.A.; Diop, A.; Mediannikov, O.; Zerrouki, H.; Diene, S.M.; Lo, S.; Dia, M.L.; Sow, A.I.; Fenollar, F.; et al. The Emergence of Carbapenem- and Colistin-Resistant Enterobacteria in Senegal. Pathogens 2023, 12, 974. https://doi.org/10.3390/pathogens12080974
Sarr H, Niang AA, Diop A, Mediannikov O, Zerrouki H, Diene SM, Lo S, Dia ML, Sow AI, Fenollar F, et al. The Emergence of Carbapenem- and Colistin-Resistant Enterobacteria in Senegal. Pathogens. 2023; 12(8):974. https://doi.org/10.3390/pathogens12080974
Chicago/Turabian StyleSarr, Habibou, Aissatou Ahmet Niang, Amadou Diop, Oleg Mediannikov, Hanane Zerrouki, Seydina M. Diene, Seynabou Lo, Mouhamadou Lamine Dia, Ahmad Iyane Sow, Florence Fenollar, and et al. 2023. "The Emergence of Carbapenem- and Colistin-Resistant Enterobacteria in Senegal" Pathogens 12, no. 8: 974. https://doi.org/10.3390/pathogens12080974